










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.26 no.2 Chapingo may./ago. 2020 Epub 23-Abr-2021
https://doi.org/10.5154/r.rchscfa.2019.04.036
Artículo científico
Crecimiento radial en Pinus patula Schltdl. & Cham. y su relación con el espacio de crecimiento y los factores climáticos
1Biodiversidad Oaxaqueña, S. C. Niño Perdido 306, Santa María Ixcotel. C. P. 71229. Santa Lucía del Camino, Oaxaca, México.
2Colegio de Postgraduados, Postgrado en Ciencias Forestales. km 36.5 carretera México-Texcoco. C. P. 56230. Montecillo, Texcoco, Estado de México, México.
3Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Campo experimental San Martinito. km 56.5 carretera México-Puebla. C. P. 74100. Tlahuapan, Puebla, México.
Introducción:
La variación anual de las características de los anillos de crecimiento tiene repercusiones en la productividad y calidad de la madera.
Objetivo:
Evaluar el crecimiento radial en dos plantaciones de Pinus patula Schltdl. & Cham. de diferente edad e intensidad de aclareo, y su relación con la variación interanual de temperatura y precipitación.
Materiales y métodos:
Las características de los anillos de crecimiento se analizaron en muestras de madera de plantaciones de 13 y 23 años, sometidas a tres intensidades de aclareo (nulo, ligero y moderado). Las características se relacionaron con los datos climáticos de los años de formación de los anillos mediante análisis de correlación de Pearson.
Resultados y discusión:
En la plantación 1995, las características de los anillos no mostraron diferencias significativas (P > 0.10) entre las parcelas con y sin aclareo, debido al exceso de competencia y recesión de la copa. La anchura de anillo, de madera temprana y tardía se asociaron significativamente (P < 0.10; r ≥ 0.40) con la temperatura mínima promedio. En la plantación 2005, los árboles de las parcelas con aclareo mostraron anchura de anillos, anchura de madera temprana e incremento en área basal mayores que los de la parcela sin aclareo. En esta plantación solo la anchura de madera tardía se asoció positivamente con la temperatura (P < 0.10; r = 0.69) y negativamente con la precipitación anual (P < 0.10; r = -0.61).
Conclusión:
El espacio de crecimiento y la intensidad de competencia afectaron principalmente la anchura de anillo y de madera temprana. Los factores ambientales influyen en las características de los anillos.
Palabras clave: Aclareo; anillos de crecimiento; competencia; madera temprana; madera tardía
Introduction:
Annual variation in growth ring characteristics has an impact on productivity and wood quality.
Objective:
To evaluate radial growth in two Pinus patula Schltdl. & Cham. plantations of different ages and thinning intensity, as well as its relationship to interannual variation in temperature and rainfall.
Materials and methods:
Growth ring characteristics were analyzed in wood samples from 13- and 23-year-old plantations, subjected to three thinning intensities (null, light and moderate). The characteristics were related to climate data during the years of ring formation using Pearson's correlation analysis.
Results and discussion:
In the 1995 plantation, ring characteristics did not show significant differences (P > 0.10) between plots with and without thinning, due to excessive competition and crown recession. Ring width, and early and latewood width, was significantly associated (P < 0.10; r ≥ 0.40) with average minimum temperature. In the 2005 plantation, trees from the thinned plots showed greater ring width, earlywood width and basal area increment than those from the unthinned plot. In this plantation only latewood width was positively associated (P < 0.10; r = 0.69) with temperature and negatively (P < 0.10; r = -0.61) with annual rainfall.
Conclusion:
Growing space and competition intensity mainly affected ring and earlywood width. Environmental factors influence ring characteristics.
Keywords: Thinning; growth rings; competition; earlywood; latewood
Introducción
Las plantaciones forestales desarrollan un papel importante para satisfacer las necesidades mundiales de fibra y madera (Schimleck, Antony, Dahlen, & Moore, 2018). Por ello, los programas de mejoramiento genético se centran en especies prioritarias para aumentar la productividad (McKeand, Mullin, Byram, & White, 2003), utilizando silvicultura intensiva. La productividad resulta del balance entre el potencial de crecimiento fisiológico (actividad fotosintética y capacidad de absorción de agua y nutrientes) y las restricciones de crecimiento debido al espacio (competencia con otros individuos y disponibilidad limitada de recursos) (Álvarez-Taboada, Barrio-Anta, Gorgoso-Varela, & Álvarez-González, 2003).
El espaciamiento inicial de los árboles influye en la programación de los tratamientos silvícolas (Hébert et al., 2016). La combinación ideal del espacio de crecimiento con la edad de rotación, calidad del sitio y tratamientos silvícolas, en particular podas y aclareos, favorece las propiedades de la madera (Guerra-Bugueño, Célis-Mosqueira, & Moreno-García, 2014; Hébert et al., 2016). El espaciamiento influye en el volumen total, forma del fuste, anillos de crecimiento, madera juvenil, y longitud y diámetro de ramas (Macdonald & Hubert, 2002).
El clima es un factor determinante en las características de los anillos de crecimiento (Haghshenas et al., 2016). Varios estudios relacionan la tasa de crecimiento radial y las características de los anillos con variables ambientales como la temperatura, precipitación e intensidad luminosa (García-Suárez, Butler, & Baillie, 2009), así como los cambios de las características de los anillos con los gradientes altitudinales (Hernández & Castellanos, 2006). Por ejemplo, la precipitación durante el invierno y principios de la primavera influyen en gran medida en el crecimiento radial de Pinus halepensis Mill., especialmente en la formación de madera temprana (Olivar, Bogino, Spiecker, & Bravo, 2012; Olivar, Rathgeber, Ordóñez & Bravo, 2013). En Pinus patula Schltdl. & Cham. se ha demostrado que las poblaciones ubicadas a mayor elevación son menos productivas que las de menor elevación; sin embargo, también puede ocurrir lo contrario si las poblaciones de menor elevación se encuentran en el extremo inferior del límite de distribución natural de la especie (Salazar et al., 1999). Debido a lo anterior, la anchura de los anillos de crecimiento y de sus componentes (madera temprana y tardía) son variables importantes y confiables para evaluar el efecto del ambiente y del espacio de crecimiento sobre la productividad y la calidad de la madera (Zhu, Scott, Scallon, & Myers, 2007). El conocimiento de esta relación facilita el estudio de las propiedades de la madera y permite separar el efecto de la edad asociado a los procesos fisiológicos del árbol, con respecto al efecto de los factores ambientales del sitio.
El objetivo general del estudio fue evaluar el crecimiento radial de P. patula en dos plantaciones con edad e intensidad de aclareo diferentes, así como su relación con la variación interanual de temperatura y precipitación. Los objetivos específicos fueron a) evaluar el efecto del espacio de crecimiento en el crecimiento radial del árbol; b) evaluar el efecto de la edad de la plantación en la respuesta del arbolado al espacio de crecimiento; y c) estimar la correlación entre el crecimiento radial de los árboles y las condiciones de temperatura y precipitación durante el año de formación del anillo.
Materiales y métodos
Área de estudio
El estudio se hizo en dos plantaciones de P. patula en la comunidad de Ixtlán de Juárez, Oaxaca. La primera se estableció en 1995, a 2 538 m de altitud (17° 22´ 38.3" N y 96° 28´ 43.2” O) y con espaciamiento inicial de 1.5 x 1.5 m (4 444 árboles·ha-1). La segunda se estableció en 2005, a 2 877 m (17° 40´ 17.6" N y 96° 46´ 05.3” O) y con espaciamiento de 2.5 x 2.5 m (1 600 árboles·ha-1).
En cada plantación se evaluaron 10 árboles de cada una de tres parcelas de 400 m2 (20 x 20 m), que incluyeron dos parcelas aclaradas y una sin aclareo. El aclareo de la plantación 1995 se hizo en 2004, 2008 y 2017, de acuerdo con el plan de manejo establecido por el prestador de servicios técnicos forestales en la comunidad. En el tercer aclareo (2017) se removió 20 % (parcela 1) y 28 % (parcela 2) de los árboles por hectárea. El aclareo de la plantación 2005 se hizo en 2016, con la remoción de 16 % (parcela 1) y 26 % (parcela 2) de los árboles, de acuerdo con el plan de manejo establecido. La densidad estimada de estas parcelas, antes del aclareo, fue 1 400 árboles·ha-1 (parcela 1) y 950 árboles·ha-1 (parcela 2). En 2018, se hicieron mediciones de diámetro normal y altura total, y se estimó el área basal y volumen total de las parcelas en cada plantación (Cuadro 1).
Cuadro 1 Características dasométricas en árboles de Pinus patula en las plantaciones de 13 y 23 años.
| Plantación | Característica dasométrica | Parcela 1 (aclareo ligero) | Parcela 2 (aclareo moderado) | Parcela 3 (sin aclareo) |
|---|---|---|---|---|
| 1995 (23 años) | NA (árboles·ha-1) | 950 | 825 | 2 475 |
| DNP (cm) | 24.20 | 27.20 | 19.73 | |
| AB (m2·ha-1) | 46.11 | 58.51 | 30.98 | |
| VOL (m3·ha-1) | 107.81 | 131.88 | 68.30 | |
| 2005 (13 años) | NA (árboles·ha-1) | 1 175 | 700 | 1 550 |
| DNP (cm) | 14.95 | 16.85 | 13.65 | |
| AB (m2·ha-1) | 18.41 | 23.44 | 15.34 | |
| VOL (m3·ha-1) | 42.35 | 44.50 | 39.65 |
NA = número de árboles, DNP = diámetro normal promedio, AB = área basal; VOL = volumen total.
Distribución y muestreo de árboles
Se elaboró un plano de distribución de los árboles en las parcelas. Cada parcela se dividió en cuadrantes de 10 x 10 m; posteriormente, se registraron ángulos y distancias de cada individuo, tomando de referencia un árbol dominante por cuadrante. Los datos se registraron en una hoja Excel versión 2013 y fueron importados por el software Quantum GIS versión 2.18 (QGIS Development Team, 2002) para crear el plano de distribución.
Con base en la distribución espacial de los árboles por parcela, se seleccionaron 10 que fueran representativos del nivel de competencia promedio en ellas. De cada árbol se extrajo la muestra de madera (viruta) a 80 cm del suelo, con el propósito de obtener un mayor número de anillos de crecimiento. La muestra se tomó perpendicular a la pendiente, con un taladro de Pressler® de 5 mm de diámetro, en la primavera de 2018 (el último anillo de la muestra se formó en 2017). Las muestras se transportaron en popotes de plástico como medio de protección, se almacenaron en un ambiente fresco, se montaron en bases de madera y se pulieron con lijas de madera (grano 60 y 100) y lijas de agua (1 200), para ver los anillos de crecimiento con detalle.
Medición de árboles vecinos
En la misma fecha del muestreo en campo se midieron los cuatro árboles vecinos más cercanos a cada árbol seleccionado (10 árboles por parcela), para estimar el índice de competencia individual y por parcela. En cada árbol seleccionado se establecieron cuadrantes siguiendo la orientación cardinal (N, E, S y O). En cada cuadrante se identificó el árbol más cercano, se midió el diámetro normal, la altura total y la distancia al árbol seleccionado. Con estos datos se calculó el índice de competencia (ICi) para cada árbol muestreado con la fórmula siguiente (Daniels, Burkhart, & Clason, 1986):
donde,
DNj |
diámetro normal del j-ésimo árbol vecino |
dij |
distancia del j-ésimo árbol vecino al i-ésimo árbol muestra. |
Con los valores de ICi se calculó el índice de competencia promedio por parcela para los 10 árboles muestreados en cada una de ellas.
Procesamiento de muestras
Las muestras de madera (viruta) de cada árbol se escanearon y las imágenes se procesaron con el software winDENDROTM (Regent Instruments Inc., 2008). En cada imagen identificada se contaron los anillos de crecimiento, tomando como referencia el anillo formado en 2017. Una vez identificado el año de formación de cada anillo, se midió la anchura total del anillo (AA, mm), así como la anchura de madera temprana (ATE) y de madera tardía (ATA). La proporción de madera tardía (PT) se estimó con la ecuación:
Con la AA se determinó el incremento anual en área basal (IAB). Para ello, se generó el crecimiento acumulado en diámetro y área basal, y por diferencia con el valor del año previo se obtuvo el IAB.
Análisis estadístico
Se hizo un análisis de varianza con el procedimiento GLM de SAS, versión 9.3 (SAS Institute, 2004), utilizando valores relativos de las características de los anillos. Los valores relativos se obtuvieron mediante el ajuste de los valores originales, con base en el valor promedio de la característica en cada árbol y parcela, para reducir el efecto de la velocidad de crecimiento o tamaño entre árboles dentro de cada parcela. La ecuación utilizada para obtener los valores relativos fue:
donde,
Xijr |
valor relativo en el i-ésimo anillo del j-ésimo árbol |
Xij |
valor absoluto en el i-ésimo anillo del j-ésimo árbol |
|
valor promedio del j-ésimo árbol |
|
valor promedio de la parcela. |
El crecimiento radial de las plantaciones y su relación con las condiciones climáticas (temperatura y precipitación) en los años de formación del anillo se comparó mediante el siguiente procedimiento: primero se eliminó el efecto de la edad cambial sobre AA y ATE con un modelo de regresión exponencial negativo; luego, a los residuales de este modelo se sumó el valor medio de la variable en la plantación, para regresar los valores a la escala original; y después con los valores ajustados de AA y ATE se estimaron, por diferencia, ATA y PT en cada anillo. Finalmente, con los valores ajustados se hizo un análisis de varianza para evaluar las diferencias de las características de los anillos formados en los años 2008 a 2017 entre plantaciones, usando el procedimiento MIXED de SAS (SAS Institute, 2004).
La relación de las características de los anillos de crecimiento con la temperatura y precipitación, durante los años de formación de la madera, se estimó con un análisis de correlación entre las variables AA, ATE y ATA con las variables de temperatura media anual, máxima promedio, mínima promedio y precipitación anual estimadas para cada uno de los sitios de plantación. Los datos climáticos se estimaron con el modelo del software “Climate NA v5.00” (Wang, Hamman, Spittlehouse, & Carroll, 2016). En el análisis se utilizó la serie de tiempo 1999-2016 para la plantación 1995 y la serie 2009-2016 para la plantación 2005. Estas series incluyen todos los anillos de crecimiento disponibles en las muestras de las dos plantaciones.
Resultados y discusión
Efecto del espacio de crecimiento en las características de los anillos
El análisis de varianza con valores relativos permitió detectar mayor número de variables con diferencias significativas (P ≤ 0.10) entre las parcelas de P. patula. En la plantación 1995 se encontraron diferencias en IAB en 2011, en ATA en 2006 y 2008, y en PT en 2008 y 2017. En cambio, la plantación 2005 sí tuvo diferencias en las cinco características evaluadas y en un mayor número de años, tal como se muestra en el Cuadro 2.
Cuadro 2 Análisis de varianza (valores de P) de las características de los anillos de crecimiento en árboles de Pinus patula en tres parcelas (con aclareo ligero, moderado y sin aclareo) de plantaciones de 13 y 23 años.
| Año de formación | Plantación 1995 (23 años) | Plantación 2005 (13 años) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| AA | IAB | ATE | ATA | PT | AA | IAB | ATE | ATA | PT | ||
| 1999 | 0.710 | 0.704 | 0.555 | 0.887 | 0.871 | ||||||
| 2000 | 0.894 | 0.819 | 0.986 | 0.702 | 0.746 | ||||||
| 2001 | 0.916 | 0.711 | 0.498 | 0.289 | 0.235 | ||||||
| 2002 | 0.222 | 0.952 | 0.436 | 0.411 | 0.883 | ||||||
| 2003 | 0.395 | 0.996 | 0.336 | 0.498 | 0.284 | ||||||
| 2004 | 0.677 | 0.885 | 0.637 | 0.848 | 0.881 | ||||||
| 2005 | 0.596 | 0.938 | 0.737 | 0.445 | 0.713 | ||||||
| 2006 | 0.900 | 0.920 | 0.629 | 0.096* | 0.303 | ||||||
| 2007 | 0.813 | 0.855 | 0.460 | 0.656 | 0.623 | ||||||
| 2008 | 0.856 | 0.748 | 0.355 | 0.031* | 0.041* | ||||||
| 2009 | 0.897 | 0.731 | 0.971 | 0.975 | 0.871 | 0.252 | 0.283 | 0.306 | 0.285 | 0.004* | |
| 2010 | 0.606 | 0.528 | 0.492 | 0.178 | 0.200 | 0.060* | 0.157 | 0.082* | 0.545 | 0.955 | |
| 2011 | 0.208 | 0.091* | 0.200 | 0.910 | 0.592 | 0.014* | 0.030* | 0.015* | 0.141 | 0.194 | |
| 2012 | 0.670 | 0.681 | 0.750 | 0.413 | 0.141 | 0.009* | 0.092* | 0.383 | 0.009* | 0.126 | |
| 2013 | 0.710 | 0.503 | 0.696 | 0.131 | 0.194 | 0.001* | 0.001* | 0.003* | 0.715 | 0.751 | |
| 2014 | 0.759 | 0.814 | 0.735 | 0.514 | 0.351 | 0.149 | 0.018* | 0.342 | 0.069* | 0.074* | |
| 2015 | 0.864 | 0.766 | 0.907 | 0.267 | 0.549 | 0.005* | 0.001* | 0.008* | 0.290 | 0.750 | |
| 2016 | 0.383 | 0.316 | 0.380 | 0.404 | 0.733 | 0.479 | 0.010* | 0.665 | 0.002* | 0.003* | |
| 2017 | 0.197 | 0.192 | 0.185 | 0.198 | 0.046* | 0.109 | 0.001* | 0.353 | 0.029* | 0.118 | |
AA = anchura de anillo, IAB = incremento en área basal, ATE = anchura de madera temprana, ATA = anchura de madera tardía, PT = proporción de madera tardía. *Los valores P ≤ 0.10 indican diferencias significativas entre parcelas.
La Figura 1 muestra que las características de los anillos de los árboles tuvieron un patrón similar entre las parcelas. En ambas plantaciones se observó un efecto juvenil con un incremento ligero de anchura de anillo (1a y 1f) en los primeros años, especialmente en la plantación joven. En la plantación 1995, las tres parcelas siguieron el mismo comportamiento. El IAB aumentó rápidamente en los primeros años, alcanzó su punto máximo a los ocho y, posteriormente, declinó de manera gradual (Figura 1b). Las características AA y ATE presentaron valores elevados, pero después de tres o cuatro años disminuyeron y en los últimos años se estabilizaron con valores muy bajos (Figuras 1a y 1c). Por otra parte, la ATA mostró fluctuaciones amplias en los primeros 10 años y después una reducción gradual en la formación de madera tardía (Figura 1d). Finalmente, la PT aumentó con la edad de los árboles (Figura 1e) y fue la única con respuesta significativa al aclareo de 2017 (Cuadro 2); el valor más alto se obtuvo en la parcela con mayor espacio de crecimiento (28 %).
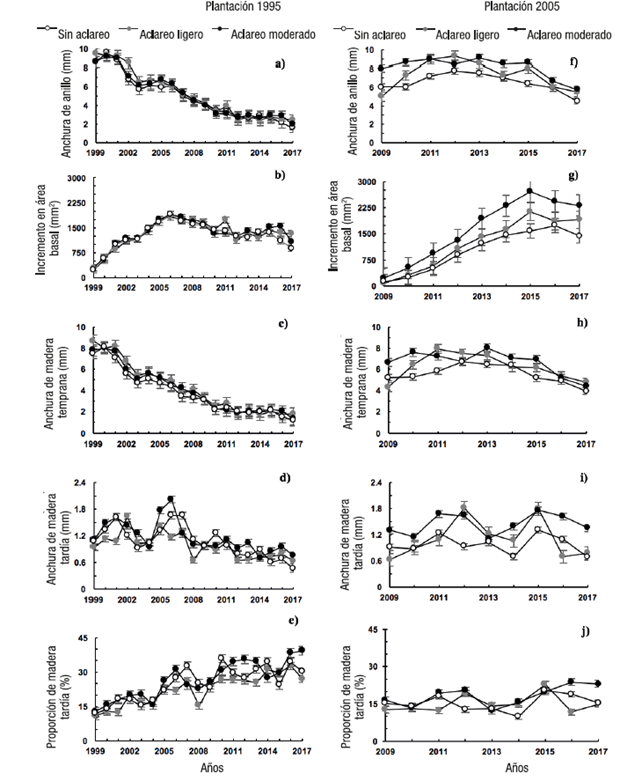
Figura 1 Comportamiento de los valores promedio (± error estándar) de las características de los anillos de crecimiento en los árboles de las parcelas con y sin aclareo en dos plantaciones de Pinus patula.
En la plantación 2005, la tendencia de AA y ATE fue similar. Dichas características aumentaron en los primeros cuatro o cinco años y luego disminuyeron gradualmente (Figura 1f y 1h). El IAB aumentó rápidamente en los primeros siete años y luego se redujo ligeramente (Figura 1g). La ATA no mostró una tendencia definida (Figura 1i), mientras que la PT presentó incremento gradual con la edad (Figura 1j). En comparación con la plantación 1995, las diferencias entre las parcelas de la plantación 2005 fueron mayores, a favor de las parcelas aclaradas, especialmente en la parcela 2 con mayor espacio de crecimiento (26 %).
De acuerdo con la literatura, el espaciamiento inicial y el preaclareo son las herramientas más efectivas, para controlar el crecimiento del árbol y la calidad de la madera en los primeros años de la plantación, ya que afectan la competencia entre árboles y el cierre del dosel (Guerra-Bugueño et al., 2014; Tong & Zhang, 2005). La plantación de 1995 se estableció con un espaciamiento reducido (1.5 x 1.5 m), lo que generó fuerte competencia desde los primeros años y recesión rápida de la copa, situación que anuló el efecto esperado de los aclareos. Además, a pesar de que en los aclareos se eliminó un número distinto de árboles, el IC promedio fue similar entre las parcelas aclaradas, debido a diferencias en las dimensiones de los árboles al momento del aclareo (0.30 ≤ IC1995 ≤ 0.39 y 0.20 ≤ IC2005 ≤ 0.28). El efecto combinado del espaciamiento inicial reducido con la recesión de la copa y la poca variación en el espacio de crecimiento entre las parcelas explican en gran medida la baja respuesta del crecimiento radial de los árboles a los aclareos realizados con fines de manejo.
Latham y Tappeiner (2002) y Auty et al. (2018) indican que la reducción de la competencia por espacio induce aumento en el crecimiento radial del árbol. A su vez, el crecimiento acelerado ocasiona mayor número de células de madera temprana y menor formación de madera tardía (Koga, Zhang, & Bégin, 2002). Por su parte, Hébert et al. (2016) mencionan que las especies de rápido crecimiento suelen producir mayor porcentaje de madera temprana. Por ello se esperaba encontrar un aumento importante en el crecimiento en diámetro de los árboles en las parcelas aclaradas; sin embargo, en la plantación 1995, dichas parcelas mostraron anillos de crecimiento muy estrechos en los últimos años, con mayor proporción de madera tardía y sin diferencias significativas (P > 0.10) con los árboles de la parcela sin aclareo, excepto en el último año. Al parecer, aunque los árboles en las dos parcelas aclaradas tenían más espaciamiento promedio, también tenían mayor diámetro y área basal por hectárea, por lo que en realidad estuvieron sometidos a una competencia similar (Arenas-Castro, Fernández-Haeger, & Jordano-Barbudo, 2015).
En la plantación 2005, los árboles en las parcelas aclaradas mostraron mayor crecimiento que en la parcela testigo desde los primeros anillos. En particular, los árboles con más espacio mostraron mayor crecimiento. De acuerdo con los datos dasométricos al momento del aclareo de 2016, la parcela 2 quedó con una densidad de 700 árboles·ha-1 con diámetro normal promedio (DNP) de 16.85 cm (Cuadro 1), mientras que las parcelas 1 y 3 presentaron mayor densidad y menor DNP. Debido al mayor DNP, el área basal de la parcela 2 era mayor en 5.03 y 8.1 m2·ha-1 con respecto a las parcelas 1 y 3, respectivamente (Cuadro 1).
Los árboles adquieren los recursos necesarios para generar fotosintatos y usarlos en diversos tejidos y funciones (Binkley, 2004). Akers, Kane, Zhao, Teskey, y Daniels (2013) señalan que la limitación de la luz influye más en el crecimiento de los árboles que la limitación de recursos del suelo. En este sentido, aunque las tres parcelas se establecieron con la misma densidad de plantación (1 600 árboles·ha-1), la parcela 2 tuvo más mortalidad inicial. Esto ocasionó mayor espaciamiento entre los árboles que sobrevivieron en esa parcela, favoreciendo el crecimiento radial.
Diferencias en las características de los anillos de los árboles entre plantaciones
El Cuadro 3 muestra que las características de los anillos de crecimiento de P. patula presentaron diferencias (P ≤ 0.10) entre las dos plantaciones durante el periodo evaluado en común (2009-2017). Los efectos fueron más notorios en AA y ATE durante los años 2009 a 2015, excluyendo solo el 2011. En el caso de la ATA hubo diferencias en 2010 y 2015, mientras que la PT fue diferente en varios años, que incluyeron el inicio y el final de la serie de tiempo en común.
Cuadro 3 Análisis de varianza (*valores significativos P ≤ 0.10) de las características de los anillos de crecimiento ajustados por edad cambial entre las dos plantaciones de Pinus patula.
| Año de formación | Anchura de anillo | Anchura de madera temprana | Anchura de madera tardía | Proporción de madera tardía |
|---|---|---|---|---|
| 2009 | <0.001* | <0.001* | 0.590 | <0.001* |
| 2010 | 0.013* | 0.034* | 0.018* | 0.107 |
| 2011 | 0.608 | 0.638 | 0.911 | 0.239 |
| 2012 | 0.008* | 0.062* | 0.107 | 0.279 |
| 2013 | 0.001* | <0.001* | 0.500 | 0.009* |
| 2014 | 0.088* | 0.012* | 0.228 | 0.002* |
| 2015 | 0.011* | 0.038* | 0.034* | 0.183 |
| 2016 | 0.501 | 0.191 | 0.348 | 0.091* |
| 2017 | 0.703 | 0.364 | 0.264 | 0.073* |
El comportamiento de las características de los anillos en las dos plantaciones fue diferente durante el periodo en común (2009-2017). De acuerdo con la Figura 2, en los primeros años (2009 y 2010), los árboles de la plantación 1995 desarrollaron anillos de crecimiento más anchos y con mayor cantidad de madera temprana que los de la plantación 2005, mientras que en el periodo 2012-2015 sucedió lo opuesto. Tal comportamiento se relaciona con el aumento gradual en la anchura de los anillos en la plantación joven, mientras que en la otra plantación, la anchura fue relativamente estable durante 2009-2017. Al parecer, la recesión de la copa en la plantación de mayor edad, debido a la competencia excesiva entre árboles, redujo la capacidad de respuesta de los anillos. Aunque se seleccionaron los árboles con competencia promedio, el nivel de competencia fue similar entre las parcelas de una misma plantación, pero no entre las dos plantaciones. La competencia fue más intensa en la plantación 1995, debido a las diferencias en el espaciamiento inicial del arbolado. Al inicio del periodo de comparación (2009), la plantación 1995 tenía una densidad promedio de 2 358 árboles·ha-1 con área basal promedio de 47.09 m2·ha-1, mientras que la plantación 2005 tenía 1 300 árboles·ha-1 con área basal de 19.06 m2·ha-1.

Figura 2 Características promedio de los anillos de crecimiento en plantaciones de Pinus patula de diferente edad.
La plantación joven presentó mayor crecimiento a partir del 2012, lo cual se atribuye al espaciamiento entre los árboles y al tamaño y posición de la copa viva (Domínguez-Domínguez, Bravo, & del Río, 2006). La longitud de la copa está determinada en gran medida por el espacio entre los árboles (Castelán-Lorenzo & Arteaga-Martínez, 2009; Gartner, North, Johnson, & Singleton, 2002). Por otro lado, al parecer, la variación en el crecimiento radial no se debe a las diferencias en la productividad del sitio, ya que las dos plantaciones tienen índice parecido (IS1995 = 33 m e IS2005 = 29 m), estimado a partir de la ecuación ajustada por Santiago-García et al. (2017) en rodales de P. patula para la misma zona de estudio. Aunque la anchura de anillo varía con las condiciones del sitio, las especies de pino mantienen una proporción estable de 30 % de madera tardía en climas fríos (Düthorn, Schneider, Günther, Gläser, & Esper, 2016); por tanto, el valor encontrado para ambas plantaciones (aproximadamente 20 %) indica un efecto combinado del potencial productivo de la especie y de las condiciones ambientales favorables. Pompa-García y Camarero-Martínez (2015) reportaron una proporción similar de madera tardía (18 %) en Pinus cooperi Blanco en sitios productivos.
La variación observada entre las características de los anillos de crecimiento en ambas plantaciones se atribuye a las condiciones ambientales en las que crecen los árboles (Schimleck et al., 2018). De acuerdo con Baldwin et al. (2000), a mayor espaciamiento inicial, los árboles desarrollan mayor diámetro y longitud de copa. Esto favorece la formación de anillos más amplios y más madera temprana. Rodríguez-Ortíz (2010) recomienda un espaciamiento inicial de 2.40 a 2.75 m para P. patula, debido a que es una especie sensible a la competencia en sus primeros años de vida.
Relación de las características de los anillos de crecimiento con las condiciones climáticas
De acuerdo con el Cuadro 4, las características de los anillos (AA, ATE y ATA) en la plantación 1995 mostraron correlación significativa (P < 0.10) y positiva únicamente con la temperatura mínima promedio (TMIP). En cambio, en la plantación 2005 solo la ATA mostró correlación significativa, la cual fue positiva con la temperatura media anual y negativa con la precipitación anual.
Cuadro 4 Coeficientes de correlación de Pearson entre las características de los anillos de crecimiento y las condiciones climáticas en dos plantaciones de Pinus patula.
| Variables | Plantación 1995 | Plantación 2005 | ||||||
|---|---|---|---|---|---|---|---|---|
| TMA | TMAP | TMIP | PA | TMA | TMAP | TMIP | PA | |
| AA | 0.054 | 0.239 | 0.506* | -0.295 | 0.156 | 0.266 | 0.120 | -0.084 |
| ATE | 0.037 | 0.343 | 0.446* | -0.248 | 0.054 | 0.190 | 0.042 | 0.009 |
| ATA | 0.063 | -0.094 | 0.398* | -0.257 | 0.698* | 0.581 | 0.534 | -0.619* |
*Coeficientes de correlación significativos P ≤ 0.10 (n1995 = 18; n2005 = 8). TMA = temperatura media anual; TMAP = temperatura máxima promedio; TMIP = temperatura mínima promedio; PA = precipitación anual; AA = anchura de anillo; ATE = anchura de madera temprana; ATA = anchura de madera tardía.
Las correlaciones positivas de AA, ATE y ATA con TMIP en la plantación 1995 (Figuras 3a, 3b y 3c) indican que el aumento en la temperatura mínima induce un aumento en el crecimiento radial. En la plantación 2005, esta relación positiva entre temperatura y crecimiento radial solo se observó para ATA (Figura 3d); en cambio, la correlación entre ATA y la precipitación anual fue negativa, la cual indica que en los años más húmedos hubo menor formación de madera tardía (Figura 3e).

Figura 3 Correlación de Pearson de las características de los anillos de crecimiento de Pinus patula con las condiciones ambientales. a, b y c: plantación 1995 (n = 18); d y e: plantación 2005 (n = 8).
La temperatura y precipitación son elementos importantes en la dinámica del crecimiento radial (Chacón-de la Cruz & Pompa-García, 2015). Por ello, se esperaba una correlación positiva entre las condiciones ambientales y el crecimiento radial para ambas plantaciones. Las características de los anillos de crecimiento en la plantación 1995 se asociaron de manera positiva con la temperatura mínima promedio. Estos resultados coinciden con el estudio de formación de madera realizado en Juniperus przewalskii Kom. (Zhang et al., 2018), así como en la anchura de madera temprana y tardía en P. cooperi (Pompa-García & Camarero-Martínez, 2015). Por otra parte, la relación negativa entre la anchura de madera tardía y la precipitación anual en la plantación 2005 fue inesperada, ya que los estudios realizados en otras especies, generalmente, han indicado una relación positiva. Por ejemplo, Olivar et al. (2013) señalan una relación positiva entre el crecimiento radial en P. halepensis con la precipitación, y atribuyen el desarrollo de la madera tardía al efecto de la lluvia invernal. Lebourgeois (2000) también encontró una relación positiva (r = 0.46) entre la anchura de madera tardía y la precipitación media mensual en Pinus nigra Arn. No obstante, debido a la dinámica de la actividad cambial durante la formación de la madera, la relación del crecimiento con las condiciones climáticas es muy compleja y variada (Żywiec et al., 2017).
La temperatura y precipitación afectan la velocidad y duración de la actividad cambial, así como la transición de madera temprana a madera tardía en las especies leñosas (Rossi, Morin, & Deslauriers, 2012). Por ejemplo, es posible que una mayor precipitación a mediados del periodo de crecimiento promueva mayor actividad del follaje y del cámbium, lo que prolonga la formación de madera temprana y retrasa la formación de madera tardía. Se ha demostrado que la transición de madera temprana a tardía está asociada con el cese del crecimiento apical (Larson, 1969) y que la formación de madera temprana favorece una alta conductividad hidráulica (Cruickshank & Filipescu, 2017). En cambio, una menor precipitación a mediados del periodo de crecimiento puede ocasionar una transición más rápida a la formación de madera tardía, como parte de la respuesta al estrés hídrico y para reducir la vulnerabilidad hidráulica (Abe, Nakai, Utsumi, & Kagawa, 2003; Domec & Gartner, 2002; Wilkinson, Ogée, Domec, Rayment, & Wingate, 2015).
Conclusiones
La respuesta de Pinus patula al espacio de crecimiento fue diferente en las plantaciones. En la plantación de mayor edad (23 años), los aclareos tuvieron un impacto reducido sobre el crecimiento radial del fuste y las características de los anillos de crecimiento, debido a que los árboles, por su mayor tamaño en las parcelas aclaradas, tuvieron niveles similares de competencia que en la parcela testigo. En la plantación de menor edad (13 años), el mayor espacio de crecimiento en las parcelas aclaradas estimuló el crecimiento radial, con mayor amplitud de anillos y madera temprana. Por lo anterior, es importante tomar en cuenta la edad, el tamaño de los árboles y el nivel de competencia entre ellos para definir la intensidad del aclareo. Por otro lado, la temperatura y la precipitación influyeron en las características de los anillos durante el periodo de formación de la madera. La temperatura tuvo efecto positivo en la anchura del anillo y de sus componentes (madera temprana y tardía), un aspecto importante en el contexto del cambio climático.
References
Abe, H., Nakai, T., Utsumi, Y., & Kagawa, A. (2003). Temporal water deficit and wood formation in Cryptomeria japonica. Tree Physiology, 23(12), 859-863. doi: 10.1093/treephys/23.12.859 [ Links ]
Akers, M. K., Kane, M., Zhao, D., Teskey, R. O., & Daniels, R. F. (2013). Effects of planting density and cultural intensity on stand and crown attributes of mid-rotation loblolly pine plantations. Forest Ecology and Management, 310, 468-475. doi: 10.1016/j.foreco.2013.07.062 [ Links ]
Álvarez-Taboada, M. F., Barrio-Anta, M., Gorgoso-Varela, J., & Álvarez-González, J. G. (2003). Influencia de la competencia en el crecimiento en sección en Pinus radiata D. Don. Investigación Agraria: Sistema Recursos Forestales, 12(2), 25-35. Retrieved from https://recyt.fecyt.es/index.php/IA/article/view/2493/1874 [ Links ]
Arenas-Castro, S., Fernández-Haeger, J., & Jordano-Barbudo, D. (2015). Estructura de edades, tamaños y crecimiento en una población local de piruétano (Córdoba, España). Ecosistemas, 24(2), 7-14. Retrieved from https://revistaecosistemas.net/index.php/ecosistemas/article/view/1053 [ Links ]
Auty, D., Moore, J., Achim, A., Lyon, A., Mochan, S., & Gardiner, B. (2018). Effects of early respacing on the density and microfibril angle of sitka spruce wood. Forestry: An International Journal of Forest Research, 91(3), 307-319. doi:10.1093/forestry/cpx004 [ Links ]
Baldwin, V. C., Peterson, K. D., Clark III, A., Ferguson, R. B., Strub, M. R., & Bower, D. R. (2000). The effects of spacing and thinning on stand and tree characteristics of 38-year-old loblolly pine. Forest Ecology and Management, 137(1-3), 91-102. doi: 10.1016/S0378-1127(99)00340-0 [ Links ]
Binkley, D. (2004). A hypothesis about the interaction of tree dominance and stand production through stand development. Forest Ecology and Management, 190(2-3), 265-271. doi: 10.1016/j.foreco.2003.10.018 [ Links ]
Castelán-Lorenzo, M., & Arteaga-Martínez, B. (2009). Establecimiento de regeneración de Pinus patula Schl. et Cham., en cortas bajo el método de árboles padres. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 15(1), 49-57. Retrieved from https://www.chapingo.mx/revistas/forestales/contenido.php?seccion=numero&id_revista_numero=39 [ Links ]
Chacón-de la Cruz, J. E., & Pompa-García, M. (2015). Response of tree radial growth to evaporation, as indicated by earlywood and latewood. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 21(1), 57-65. doi: 10.5154/r.rchscfa.2014.10.050 [ Links ]
Cruickshank, M. G., & Filipescu, C. N. (2017). The interactive effect of root disease and climate on wood properties in halfsibling Douglas-fir families. Forest Ecology and Management, 392, 58-67. doi: 10.1016/j.foreco.2017.03.002 [ Links ]
Daniels, R. F., Burkhart, H. E., & Clason, T. R. (1986). A comparison of competition measures for predicting growth of Ioblolly pine trees. Canadian Journal of Forest Research, 16(6), 1230-1237. doi: 10.1139/x86-218 [ Links ]
Domec, J. C., & Gartner, B. L. (2002). How do water transport and water storage differ in coniferous earlywood and latewood? Journal of Experimental Botany, 53(379), 2369-2379. doi: 10.1093/jxb/erf100 [ Links ]
Domínguez-Domínguez, M., Bravo, F., & del Río, M. (2006). Modelo del tamaño de copa de Pinus sylvestris L. en bosque del centro de España. Interciencia, 31(3), 168-175. Retrieved from http://www.redalyc.org/pdf/339/33911403.pdf [ Links ]
Düthorn, E., Schneider, L., Günther, B., Gläser, S., & Esper, J. (2016). Ecological and climatological signals in tree-ring width and density chronologies along a latitudinal boreal transect. Scandinavian Journal of Forest Research, 31(8), 750-757. Retrieved from https://www.tandfonline.com/doi/abs/10.1080/02827581.2016.1181201 [ Links ]
García-Suárez, A. M., Butler, C. J., & Baillie, M. G. L. (2009). Climate signal in tree-ring chronologies in a temperate climate: A multi-species approach. Dendrochronologia, 27(3), 183-198. doi: 10.1016/j.dendro.2009.05.003 [ Links ]
Gartner, B. L., North, E. M., Johnson, G. R., & Singleton, R. (2002). Effects of live crown on vertical patterns of wood density and growth in Douglas-fir. Canadian Journal of Forest Research, 32(3), 439-447. doi: 10.1139/x01-218 [ Links ]
Guerra-Bugueño, E., Célis-Mosqueira, F., & Moreno-García, N. (2014). Efecto de la densidad de plantación en la rentabilidad de plantaciones de Eucalyptus globulus. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 20(1), 21-31. doi: 10.5154/r.rchscfa.2012.08.051 [ Links ]
Haghshenas, M., Mohadjer, M. R. M., Attarod, P., Pourtahmasi, K., Feldhaus, J., & Sadeghi, S. M. M. (2016). Climate effect on tree-ring widths of Fagus orientalis in the Caspian forests, northern Iran. Forest Science and Technology, 12(4), 176-182. doi: 10.1080/21580103.2016.1144542 [ Links ]
Hébert, F., Krause, C., Pierre-Yves, P., Achim, A., Prégent, G., & Ménétrier, J. (2016). Effect of tree spacing on tree level volume growth, morphology, and wood properties in a 25-year-old Pinus banksiana plantation in the boreal forest of Quebec. Forests, 7(11), 1-16. doi: 10.3390/f7110276 [ Links ]
Hernández, L., & Castellanos, H. (2006). Crecimiento diamétrico arbóreo en bosques de Sierra de Lema, Guayana venezolana: Primeras evaluaciones. Interciencia, 31(11), 779-786. Retrieved from http://www.redalyc.org/articulo.oa?id=33912205 [ Links ]
Koga, S., Zhang, S. Y., & Bégin, J. (2002). Effects of precommercial thinning on annual radial growth and wood density in balsam fir (Abies balsamea). Wood and Fiber Science, 34(4), 625-642. Retrieved from https://wfs.swst.org/index.php/wfs/article/viewFile/329/329 [ Links ]
Larson, P. R. (1969). Wood formation and the concept of wood quality. Retrieved from https://www.fs.fed.us/nrs/pubs/other/1969/nc_1969_larson_001.pdf [ Links ]
Latham, P., & Tappeiner, J. (2002). Response of old-growth conifers to reduction in stand density in western Oregon forests. Tree Physiology, 22(2-3), 137-146. doi: 10.1093/treephys/22.2-3.137 [ Links ]
Lebourgeois, F. (2000). Climatic signals in earlywood, latewood and total ring width of Corsican pine from western France. Annals of Forest Science, 57(2), 155-164. doi: 10.1051/forest:2000166 [ Links ]
Macdonald, E., & Hubert, J. (2002). A review of the effects of silviculture on wood quality. Forestry, 75(2), 107-138. Retrieved from https://open.library.ubc.ca/cIRcle/collections/ undergraduateresearch/52966/items/1.0103111 [ Links ]
McKeand, S., Mullin, T., Byram, T., & White, T. (2003). Deployment of genetically improved loblolly and slash pines in the south. Journal of Forestry, 101(3), 32-37. Retrieved from https://academic.oup.com/jof/article/101/3/32/4608659 [ Links ]
Olivar, J., Bogino, S., Spiecker, H., & Bravo, F. (2012). Climate impact on growth dynamic and intra-annual density fluctuations in Aleppo pine (Pinus halepensis) trees of different crown classes. Dendrochronologia, 30(1), 35-47. doi: 10.1016/j.dendro.2011.06.001 [ Links ]
Olivar, J., Rathgeber, C. B. K., Ordoñez, C., & Bravo, F. (2013). Influencia del clima en la densidad de la madera de pinos mediterráneos (Pinus halepensis y Pinus pinaster). In Sociedad Española de Ciencias Forestales (Ed.), 6° Congreso Forestal Español (pp. 1-13). España: Author. Retrieved from https://www.researchgate.net/publication/262270215_Influencia_del_clima_en_la_densidad_de_la_madera_de_pinos_mediterraneos_Pinus_halepensis_y_Pinus_pinaster [ Links ]
Pompa-García, M., & Camarero-Martínez, J. J. (2015). Potencial dendroclimático de la madera temprana y tardía de Pinus cooperi Blanco. Agrociencia, 49(2), 177-187. Retrieved from http://www.scielo.org.mx/pdf/agro/v49n2/v49n2a6.pdf [ Links ]
QGIS Development Team. (2002). QGIS Geographic Information System. Open source geospatial foundation project. Retrieved from https://docs.qgis.org/3.4/es/docs/user_manual/ [ Links ]
Regent Instruments Inc. (2008). WinDENDRO™ An image analysis system for tree ring analysis. Quebec, Canadá: Author. [ Links ]
Rodríguez-Ortíz, G. (2010). Efecto de aclareos en el crecimiento de una plantación de Pinus patula Schl. et Cham. en Ixtlán, Oaxaca. Tesis doctoral, Colegio de Postgraduados, Montecillo. Texcoco, Estado de México, México. Retrieved from https://nanopdf.com/download/efecto-de-aclareos-en-el-crecimiento-de_pdf [ Links ]
Rossi, S., Morin, H., & Deslauriers, A. (2012). Causes and correlations in cambium phenology: towards an integrated framework of xylogenesis. Journal of Experimental Botany, 63(5), 2117-2126. doi: 10.1093/jxb/err423 [ Links ]
Salazar, G. J. G., Vargas, H. J. J., Jasso, M. J., Molina, G. J. D., Ramírez, H. C., & López, U. J. (1999). Variación en el patrón de crecimiento en altura de cuatro especies de Pinus en edades tempranas. Madera y Bosques, 5(2), 19-34. doi: 10.21829/myb.1999.521345 [ Links ]
Santiago-García, W., Pérez-López, E., Quiñonez-Barraza, G., Rodríguez-Ortiz, G., Santiago-García, E., Ruiz-Aquino, F., & Tamarit-Urias, J. C. (2017). A dynamic system of growth and yield equations for Pinus patula. Forests, 8(12), 1-13. doi: 10.3390/f8120465 [ Links ]
SAS Institute Inc. (2004). The SAS system for windows. Versión 9.4. Cary, NC, USA: Author. [ Links ]
Schimleck, L., Antony, F., Dahlen, J., & Moore, J. (2018). Wood and fiber quality of plantation-grown conifers: A summary of research with an emphasis on loblolly and radiata pine. Forests, 9(6), 1-16. doi: 10.3390/f9060298 [ Links ]
Tong, Q. J., & Zhang, S. Y. (2005). Impact of initial spacing and precommercial thinning on jack pine tree growth and stem quality. Forestry Chronicle, 81(3), 418-428. doi: 10.5558/tfc81418-3 [ Links ]
Wang, T., Hamann, A., Spittlehouse, D., & Carroll, C. (2016). Locally downscaled and spatially customizable climate data for historical and future periods for North America. PLoS ONE, 11(6), e0156720. doi: 10.1371/journal.pone.0156720 [ Links ]
Wilkinson, S., Ogée, J., Domec, J. C., Rayment, M., & Wingate, L. (2015). Biophysical modelling of intra-ring variations in tracheid features and wood density of Pinus pinaster trees exposed to seasonal droughts. Tree Physiology, 35(3), 305-318. doi: 10.1093/treephys/tpv010 [ Links ]
Zhang, J., Gou, X., Pederson, N., Zhang, F., Niu, H., Zhao, S., & Wang, F. (2018). Cambial phenology in Juniperus przewalskii along different altitudinal gradients in a cold and arid region. Tree Physiology, 38(6), 840-852. doi: 10.1093/treephys/tpx160 [ Links ]
Zhu, J. Y., Scott, C. T., Scallon, K. L., & Myers, G. C. (2007). Effects of plantation density on wood density and anatomical properties of red pine (Pinus resinosa Ait.). Wood and Fiber Science, 39(3), 502-512. Retrieved from https://www.fs.usda.gov/ treesearch/pubs/29256 [ Links ]
Żywiec, M., Muter, E., Zielonka, T., Delibes, M., Calvo, G., & Fedriani, J. M. (2017). Long-term effect of temperature and precipitation on radial growth in a threatened thermo-Mediterranean tree population. Trees - Structure and Function, 31(2), 491-501. doi: 10.1007/s00468-016-1472-8 [ Links ]
Recibido: 22 de Abril de 2019; Aprobado: 27 de Noviembre de 2019
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
