










Services on Demand
Journal
Article
 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
On-line version ISSN 2007-4018Print version ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.26 n.1 Chapingo Jan./Apr. 2020 Epub Mar 03, 2021
https://doi.org/10.5154/r.rchscfa.2018.09.064
Artículo científico
Crecimiento radial de especies de pino en rodales sujetos a cortas de selección en Santa María Lachixío, Oaxaca, México
1Colegio de Postgraduados, Postgrado en Ciencias Forestales. km 36.5 carretera México-Texcoco. C. P. 56230. Montecillo, Texcoco, Estado de México, México.
2Universidad Autónoma Chapingo, División de Ciencias Forestales. km 38.5 carretera México-Texcoco. C. P. 56230. Chapingo, Texcoco, Estado de México, México.
Introducción:
La evaluación del efecto de actividades de manejo sobre el arbolado es primordial para obtener un mejor conocimiento de los procesos ocurridos en los rodales.
Objetivo:
Evaluar el efecto de las cortas de selección sobre el crecimiento de especies de pino en rodales incoetáneos.
Materiales y métodos:
Se establecieron 30 sitios de muestreo en dos rodales mezclados de pino intervenidos con cortas de selección en el año 2003. Se utilizaron los tocones de dicho aprovechamiento como centros de sitio y se evaluó el crecimiento del arbolado residual en un radio de 10 m; se calcularon variables dasométricas e índices de competencia.
Resultados y discusión:
En el primer quinquenio posterior al año de cosecha, 47 % del arbolado residual mostró crecimiento significativo (P ≤ 0.05). El diámetro de tocón, diámetro normal y la competencia fueron las variables más importantes para explicar el crecimiento. El área de competencia de los individuos aprovechados (tocones) mostró influencia hasta un radio de 15 m. El incremento en área basal fue de 71 % con respecto al crecimiento anterior a la cosecha. Este aumento se relacionó con la suma de área basal del arbolado residual y se estableció 1.3 m2 por sitio (41.3 m2·ha-1) como umbral a partir del cual una liberación se nulifica por la competencia residual.
Conclusión:
Las cortas de selección influyen en el crecimiento del arbolado residual de manera diferenciada; la respuesta puede ser positiva, negativa o nula en función del tamaño y de la competencia local.
Palabras clave: anillos de crecimiento; índices de competencia; bosque incoétaneo; incremento en área basal; manejo forestal
Introduction:
The evaluation of the effect of management activities on trees is essential to gain a better understanding of the processes occurring in the stands.
Objective:
To evaluate the effect of selection cutting on the growth of pine species in uneven-aged stands.
Materials and methods:
Thirty sampling sites were established in two mixed pine stands subjected to selection cutting in 2003. The stumps of this harvest were used as site centers and the growth of the residual trees was evaluated in a 10-m radius; tree-size variables and competition indices were calculated.
Results and discussion:
In the first five years after the harvest, 47 % of residual trees showed significant growth (P ≤ 0.05). Stump diameter, diameter at breast height and competition were the most important variables to explain growth. The competition area of the harvested individuals (stumps) showed influence up to a 15-m radius. The increase in basal area was 71 % with respect to pre-harvest growth. This increase was related to the basal area sum of the residual trees and 1.3 m2 per site (41.3 m2·ha-1) was established as the threshold from which a release is nullified by residual competition.
Conclusion:
Selection cutting influences residual tree growth in a differentiated way; the response may be positive, negative or nil depending on size and local competition.
Keywords: growth rings; competition índices; uneven-aged forest; increase in basal area; forest management
Introducción
Una de las premisas establecidas en el manejo forestal, en particular en el aprovechamiento maderable, es que las actividades realizadas en la masa forestal deben propiciar el incremento del arbolado residual, lo cual no sucede con frecuencia (Powers, Pregitzer, Palik, & Webster, 2009). En México, del total de superficie bajo manejo forestal, aproximadamente 50 % se realizaba bajo métodos selectivos en el 2001 (Comisión Nacional Forestal [CONAFOR], 2001). Desafortunadamente, los datos actualizados a nivel nacional son difíciles de obtener. No obstante, los métodos selectivos han evidenciado deficiencias en sus bases o en sus procedimientos, tales como la aplicación generalizada de fórmulas y métodos sin distinción entre condiciones ecológicas diversas o la suposición del crecimiento exponencial del arbolado residual durante el turno (Hernández-Díaz, Corral-Rivas, Quiñonez-Chávez, Bacon-Sobbe, & Vargas-Larreta., 2008; Torres-Rojo, 2000).
La competencia como factor limitante de crecimiento radial de los árboles ha sido un tema de interés desde hace décadas (Cambrón-Sandoval, Suzán-Azpiri, Vargas-Hernández, Sánchez-Vargas, & Sáenz-Romero, 2013; Looney, D’Amato, Fraver, Palik, & Reinikainen, 2016; Tomé & Burkhart, 1989). Varios autores han evaluado los aclareos aplicados en rodales coetáneos y sus efectos sobre diversas variables dasométricas (Espinosa, García, & Valeria, 1994; Rodríguez-Ortiz et al., 2011; Kang, Shibuya, & Shin, 2014); sin embargo, los trabajos que han incursionado en la evaluación de rodales incoetáneos son pocos, debido principalmente a la complejidad de los factores que intervienen en los procesos de crecimiento y desarrollo de los árboles (Forget, Nolet, Doyon, Delagrange, & Jardon, 2007).
El presente trabajo se enfocó en evaluar el crecimiento del arbolado residual en rodales mixtos e incoetáneos de pino que han sido aprovechados con métodos de selección, para analizar algunas de las variables que influyen en el crecimiento postcosecha y los procesos que ocurren como resultado de las actividades silviculturales.
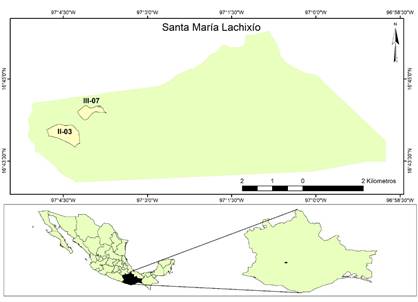
Materiales y métodos
El área de estudio se ubica en el municipio de Santa María Lachixío en el estado de Oaxaca, México (Figura 1). El muestreo se hizo en dos rodales dominados por las especies Pinus douglasiana Martínez, P. herrerae Martínez, P. lawsonii Roezl ex Gordon, P. leiophylla Schiede ex Schltdl. & Cham., P. oocarpa Schiede ex Schltdl, P. pringlei Shaw y P. pseudostrobus Lindl. Los rodales fueron identificados en el programa de manejo como II-03 con superficie de 43.07 ha y III-07 de 20.3 ha. El aprovechamiento comercial de los rodales se realizó en el año 2003 con intensidades de corta de 27.8 y 33.1 %, respectivamente. Para fines de este estudio, se establecieron 15 puntos de muestreo de manera aleatoria en cada rodal, considerando un área interna de 20 m desde la periferia para anular el efecto orilla. En cada punto se ubicó el tocón más cercano que presentara la marca de autorización para aprovechamiento forestal maderable de manera visible y a partir del cual existiera al menos un individuo vivo en un radio de 10 m.
Cada tocón se utilizó como el centro del sitio de muestreo a partir del cual se delimitó un sitio circular con radio de 10 m, para abarcar tanto árboles residuales con influencia de competencia que oscilaran entre 0 y 8.5 m, como aquellos sin influencia (Fraver et al., 2014). Todos los árboles vivos dentro del sitio de muestreo se evaluaron, para lo cual se obtuvieron dos núcleos de madera con taladro de Pressler de 6 mm, se registraron las variables dasométricas diámetro normal (cm) y altura (m), y se determinaron la distancia y el diámetro normal de los árboles competidores. Para clasificar a un árbol como “competidor” se utilizó el método de parcela móvil de Bitterlich con un factor de área basal de 4 (Crecente-Campo, Vázquez-Gómez, Rodríguez-Soalleiro, & Diéguez-Aranda, 2007). En cada sitio de muestreo se registraron además las siguientes variables: coordenadas geográficas del tocón, diámetro del tocón (cm), altitud (m), pendiente promedio (grados) y exposición (azimut). Posteriormente, para cada árbol evaluado, se calcularon tres índices de competencia dependientes de la distancia que se utilizaron como variables explicativas del crecimiento. Los índices fueron los siguientes:
Daniels, Burkhart, y Clason (1986)
donde,
di |
diámetro normal del árbol objetivo (cm) |
dj |
diámetro normal del árbol competidor (cm) |
Distij |
distancia en proyección horizontal entre el árbol objetivo y el árbol competidor (m) |
n |
número de árboles competidores. |
Los núcleos de madera se procesaron mediante técnicas estándar de dendrocronología para su montado, lijado y fechado; este último se realizó con el programa WinDENDRO™ (Regent Instruments Inc., 2009) con apoyo de los programas COFECHA (Grissino-Mayer, 2001) y Microsoft Excel®. Del programa WinDENDRO™ se obtuvieron series de datos de ancho de anillo para cada muestra procesada, las cuales se utilizaron para calcular el incremento en área basal (IAB):
La variable IAB es menos dependiente de la edad que el ancho de anillo, la cual se mantiene estable en árboles maduros como los de este estudio (Biondi, 1999; Martín-Benito, Del Rio, Heinrich, Helle, & Cañellas, 2010).
Las tendencias de crecimiento se analizaron mediante el IAB en los periodos 1993-2002 y 2004-2013; el primer periodo se estableció como testigo o referencia del crecimiento previo a la intervención o aprovechamiento maderable, y el segundo periodo se estableció como el periodo de crecimiento que reflejaría los efectos de tal intervención. El periodo posterior a la intervención de aprovechamiento se dividió en dos quinquenios, de tal forma que la respuesta del crecimiento de los primeros años y a corto plazo (menos de 10 años) se pudiera analizar con mayor detalle (Jones, Domke, & Thomas, 2009; Kang, Shibuya, & Shin, 2014).
El IAB de cada anillo de crecimiento para un mismo año es relativo al diámetro de los individuos; para poder compararlos se calculó el porcentaje de incremento en área basal (PIAB), variable similar a la utilizada en otros estudios (Jones & Thomas, 2004; Powers et al., 2009) y estandarizada e independiente del diámetro. El PIAB se calculó con la fórmula siguiente:
donde,
C1 |
periodo de crecimiento 1, correspondiente al IAB promedio de los 10 años previos a la intervención silvícola (1993-2002). |
C2 |
periodo de crecimiento 2, correspondiente al IAB promedio los 10 años posteriores a la intervención silvícola (2004-2013). |
La prueba de normalidad de Kolmogorov-Smirnov no sustentó la distribución normal de los datos de PIAB; por lo anterior, los valores se transformaron a logaritmo natural. Para evitar números no existentes, matemáticamente hablando, se sumó un valor de 100 a todos los datos, previo a la trasformación.
Análisis estadístico
Las variables de mayor influencia en el crecimiento radial se identificaron con el método de selección stepwise (Hocking, 1976) en el programa SAS (Statistical Analysis Systems Institute, 2014). Este método utiliza un valor crítico de α = 0.15 de manera predeterminada para la discriminación de variables significativas; las variables ingresadas para verificación de significancia fueron diámetro normal, altura, distancia al tocón, edad, índices de competencia, diámetro del tocón y altitud.
Las variables seleccionadas con el método stepwise se utilizaron para modelar el logaritmo natural del PIAB (Ln_PIAB) mediante regresión lineal múltiple, para lo cual se utilizó un modelo con la estructura siguiente:
donde,
β |
parámetro de intersección |
α |
parámetro de la variable “X” |
x |
variable del modelo |
n |
número de variable |
ɛ |
error del modelo. |
Las variables categóricas (especie y exposición) y de sitio (pendiente) se sometieron a un análisis de varianza de un factor. Posteriormente, para las variables categóricas, se hizo una prueba de comparación de medias de Tukey-Kramer (P ≤ 0.05) debido a que los datos son desbalanceados en los grupos, mientras que la variable pendiente se analizó con regresión, por tratarse de una variable continua y por las tendencias mostradas. El procesamiento y análisis de datos se realizó con el programa SAS (2014).
Resultados
Crecimiento radial a nivel de árbol individual
Se colectó un par de muestras de cada uno de los 204 individuos considerados como árbol objetivo en los 30 sitios de muestreo; sin embargo, las muestras de siete árboles no se pudieron fechar adecuadamente por exceso de resina y falta de contraste en los anillos, y tres árboles presentaron edad menor de 24 años y, por tanto, no aportaban información del periodo 1993-2002. Debido a lo anterior, la muestra final fue de 194 árboles objetivo (Cuadro 1).
Cuadro 1 Resumen del tamaño de muestra por especie y sus características dasométricas.
| Especie | Muestra | Diámetro normal (cm) | Altura (m) |
| Pinus douglasiana | 49 | 37.6 ± 13.96 | 27.9 ± 6.99 |
| Pinus herrerae | 52 | 35.7 ± 16.97 | 26.3 ± 8.64 |
| Pinus lawsonii | 37 | 31.1 ± 10.63 | 23.7 ± 6.52 |
| Pinus leiophylla | 2 | 40.5 ± 4.55 | 35.5 ± 3.43 |
| Pinus oocarpa | 1 | 50.0 | 29.6 |
| Pinus pringlei | 20 | 35.3 ± 18.88 | 26.0 ± 8.89 |
| Pinus pseudostrobus | 33 | 36.2 ± 13.80 | 27.1 ± 8.66 |
| Total | 194 | 35.5 | 26.4 |
± desviación estándar de la media
En virtud de que la especie no resultó ser una variable importante (no significativa con P > 0.05) para explicar el Ln_PIAB, tal y como se detalla en la sección correspondiente a variables del sitio, los análisis posteriores se realizaron para el conjunto total de muestras. De este modo, se obtuvieron tres tipos de respuesta o grupos de crecimiento de los árboles como resultado de las pruebas de comparación de medias de Tukey (α = 0.05) realizadas para cada árbol objetivo. Para ello, el crecimiento promedio (IAB) de los 10 años posteriores al aprovechamiento se contrastó con el crecimiento promedio de la década anterior y las tendencias encontradas fueron: significativamente mayor (G1), no significativo (G2) y significativamente menor (G3) (Cuadro 2). En el grupo G1, el IAB del periodo posterior a la cosecha fue mayor al IAB del periodo anterior; es decir, hubo aumento en el crecimiento. En el grupo G2, el IAB es igual antes y después de la cosecha, por lo tanto, no hubo cambios en el crecimiento. En el grupo G3, el IAB del periodo postcosecha es menor al IAB del periodo anterior; es decir, la tasa de crecimiento disminuyó. El valor anual de IAB de los individuos pertenecientes a cada grupo se promedió en el periodo de evaluación 1993-2013, para conocer la tendencia de crecimiento (Figura 2A).
Cuadro 2 Grupos y subgrupos de árboles de especies de Pinus, de acuerdo con los patrones de crecimiento en el área basal durante dos quinquenios posteriores a la cosecha.
| Grupo | Subgrupo | Quinquenio 1* | Quinquenio 2* | Individuos | Proporción (%) | PIAB |
| G1 | G1.1 | 1 | 1 | 9 | 5 | 248 |
| G1 | G1.2 | 1 | 0 | 50 | 26 | 145 |
| G1 | G1.3 | 1 | -1 | 18 | 9 | 73 |
| G1 | G1.4 | 0 | 0 | 6 | 3 | 47 |
| Subtotal | --- | --- | 83 | 43 | 133 | |
| G2 | G2.1 | 1 | 0 | 3 | 2 | 27 |
| G2 | G2.2 | 1 | -1 | 9 | 5 | 12 |
| G2 | G2.3 | 0 | 1 | 4 | 2 | 22 |
| G2 | G2.4 | 0 | 0 | 45 | 23 | 3 |
| G2 | G2.5 | 0 | -1 | 11 | 6 | -7 |
| G2 | G2.6 | -1 | 1 | 2 | 1 | -13 |
| Subtotal | --- | --- | 74 | 38 | 4 | |
| G3 | G3.1 | 0 | 0 | 5 | 3 | -36 |
| G3 | G3.2 | 0 | -1 | 7 | 4 | -36 |
| G3 | G3.3 | -1 | 0 | 18 | 9 | -46 |
| G3 | G3.4 | -1 | -1 | 7 | 4 | -46 |
| Subtotal | --- | --- | 37 | 19 | -43 | |
| Total | 194 | 100 | 71 | |||
*Los números representan la diferencia con respecto al grupo anterior: 1 = crecimiento significativo positivo, 0 = crecimiento no significativo, y -1= crecimiento significativo negativo; PIAB = porcentaje de incremento en área basal.

Figura 2 Tendencias de crecimiento en grupos y subgrupos de crecimiento de área basal (IAB) en especies de Pinus, antes y después del aprovechamiento comercial de los rodales en el 2003 (línea roja).
Posteriormente, los individuos pertenecientes a cada grupo (G1, G2 y G3) se analizaron nuevamente para dos periodos de cinco años posteriores a la cosecha (2004-2008 y 2009-2013), comparándolos con el periodo anterior inmediato respectivamente. Al comportamiento del crecimiento de los individuos dentro de cada grupo se le denominó subgrupos. Los valores crecientes estadísticamente significativos fueron caracterizados con el valor de 1. Además, están los individuos que no presentaron crecimiento significativo; es decir, presentaron crecimiento constante antes y después de los periodos analizados y fueron caracterizados con 0. Finalmente, otros presentaron crecimiento negativo; es decir, su crecimiento fue significativamente menor en el periodo analizado en comparación con el del periodo previo y se caracterizaron como -1. La combinación de las tendencias de crecimiento (1, 0, -1) de los individuos en cada grupo en el quinquenio 1 (2004-2008) con el quinquenio 2 (2009-2013) produjo 14 subgrupos (Cuadro 3).
Cuadro 3 Parámetros estimados y estadísticos de la regresión lineal de las variables significativas en el crecimiento del arbolado residual de Pinus, seleccionadas con el procedimiento stepwise.
| Variable | Parámetros estimados (α) | Error estándar | Pr > F |
| Intercepto | 7.56019 | 1.24559 | <.0001 |
| Diámetro de tocón (X1) | 0.00915 | 0.00257 | 0.0005 |
| Distancia al tocón (X2) | -0.03267 | 0.01567 | 0.03840 |
| Índice de competencia de Hegyi (X3) | -0.18319 | 0.04653 | 0.0001 |
| Diámetro normal (X4) | -0.01326 | 0.00338 | 0.0001 |
| Elevación (X5) | -0.00085381 | 0.00049929 | 0.0889 |
modelo: Ln_PIAB = α + α1x1 + α2x2 + α3x3 + α4x4 + α5x5 + ε. α = 0.05; R2 ajustada = 0.156.
Los subgrupos con valor 1 en el primer quinquenio fueron cinco (tres del grupo G1 y dos en el G2) que representan 45.9 % (n = 89) del total de la muestra; seis subgrupos presentaron 0 en el primer quinquenio postcosecha y representan 40.2 % (n = 78) del total de la muestra; por último, solo tres subgrupos mostraron valor de -1 y representan 13.29 % (n = 27) del total de la muestra (Cuadro 2).
El crecimiento de los cuatro subgrupos del G1 se presenta en la Figura 2B. El subgrupo G1.1 aumentó su crecimiento en ambos quinquenios, a pesar de que el crecimiento anterior a la cosecha fue el más bajo de todos los subgrupos. El subgrupo G1.2 posee el mayor número de individuos del total de muestras con 26 % (Cuadro 2) y presenta un comportamiento positivo de crecimiento en el primer quinquenio y no significativo en el segundo quinquenio. En el subgrupo G1.3 se observa una respuesta al crecimiento que dura aproximadamente cuatro años, pero en el quinto año presenta una reducción. Finalmente, el subgrupo G1.4 muestra crecimiento irregular, aunque constante en ambos quinquenios.
Los subgrupos incluidos en el grupo G2 se muestran en la Figura 2C. El grupo G2.4 representa 23 % del total de muestras y es el de mayor representación en el grupo G2 (Cuadro 2). Los subgrupos G2.1 y G2.2 aumentaron su crecimiento durante los primeros cuatro o cinco años posteriores a la cosecha y después la tasa disminuyó. El subgrupo G2.6, aunque solo es representado por dos individuos, presentó un descenso significativo del crecimiento en el primer quinquenio postcosecha del cual se recupera hasta el sexto año. El resto de los subgrupos de crecimiento presentaron una tendencia relativamente estable en el crecimiento antes y después de la intervención.
Los subgrupos del grupo G3 se presentan en la Figura 2D; se observa que el patrón general de este conjunto es de disminución en el crecimiento. El subgrupo G3.2 incrementó el crecimiento en los tres primeros años posteriores a la intervención, aunque no resultó significativo estadísticamente (P > 0.05). Por otro lado, el subgrupo G3.4 se diferenció del resto por los valores altos de IAB en el periodo precosecha, los cuales se pierden en los años posteriores hasta alcanzar valores semejantes al resto de los subgrupos.
Selección de variables y regresión
Se utilizaron nueve variables para realizar el proceso de selección stepwise en el programa SAS; cinco de ellas fueron importantes para explicar la respuesta en crecimiento del arbolado residual, mismas que se usaron para elaborar un modelo de regresión lineal múltiple (Cuadro 3). Los resultados de la regresión mostraron que la elevación es la única variable no significativa.
Se observa que, si bien el modelo de regresión múltiple proporcionó resultados que permiten discernir la influencia de cada variable sobre el PIAB, el porcentaje de la variabilidad total que este modelo explica es bajo (R2 = 0.16). Por lo anterior, se hizo un análisis gráfico de las variables más importantes en el modelo general de manera individual.
La Figura 3 presenta el comportamiento del logaritmo del PIAB con respecto a las cuatro variables de mayor importancia para explicar la respuesta del crecimiento. En las cuatro gráficas se presenta la línea de regresión de cada una de las variables (línea punteada), la ecuación de regresión, el coeficiente de determinación (R2) y la línea de crecimiento cero (PIAB = 0) que se ubica en la escala de logaritmo con valor 4.605.
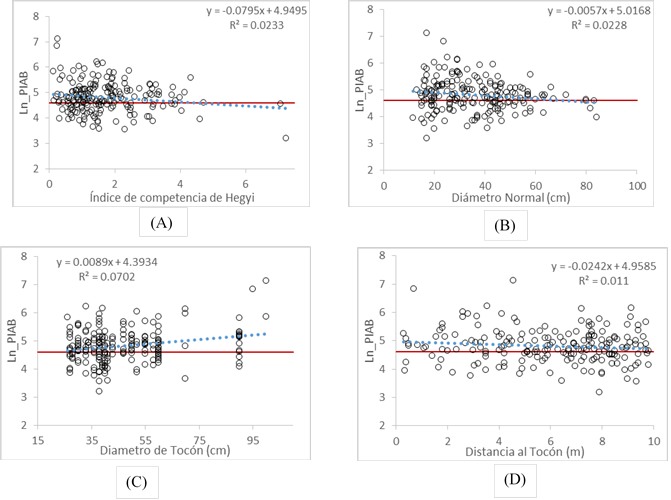
Figura 3 Regresión lineal de las variables más importantes en la respuesta del crecimiento del arbolado residual de Pinus, después de las cortas de selección.
En este estudio, el diámetro del tocón fue la variable más importante para explicar la respuesta del crecimiento a las cortas (Figura 3C), seguida del índice de competencia de Hegyi (Figura 3A) y del diámetro normal (Figura 3B). En el caso del índice de Hegyi, la línea de regresión interseca a la de crecimiento cero en un valor aproximado de 4.3, lo que indica que, en este nivel de competencia, el arbolado residual ya no responde a la liberación por cortas de selección.
La relación entre el Ln_PIAB con el diámetro normal del arbolado muestra una tendencia negativa (Figura 3B). La línea de regresión interseca a la línea de crecimiento cero en 72 cm, valor promedio después del cual es probable que el individuo no responda a la liberación de espacio derivada de las cortas realizadas.
Se observa una relación positiva entre el Ln_PIAB y el diámetro del tocón (Figura 3C); en este caso, la línea de regresión interseca al crecimiento cero a los 23.7 cm, diámetro por debajo del cual es poco probable que el espacio de crecimiento que libera un ejemplar cortado impacte de manera positiva en el crecimiento del arbolado residual.
La distancia al tocón se relaciona con el Ln_PIAB de manera negativa. La línea de regresión no interseca al valor de crecimiento cero dentro del radio de muestreo establecido en la evaluación (10 m); sin embargo, fue posible proyectar la línea con los parámetros obtenidos y se pudo ubicar dicha intersección a los 14.6 m, lo cual se interpreta como la distancia límite en la cual los árboles remanentes se ven influenciados por la corta de un individuo competidor.
Variables a nivel de sitio
La influencia de la exposición del sitio sobre Ln_PIAB resultó significativa (P = 0.014). El efecto de la exposición SE difiere en la respuesta del crecimiento (Ln_PIAB), ya que fue menor que el resto de las exposiciones (Figura 4). En el caso de la especie, la prueba no mostró variación significativa en el Ln_PIAB (P > 0.1854).

Figura 4 Porcentaje de incremento de área basal (Ln_PIAB) en especies de Pinus con respecto a la exposición del sitio. Medias con letras distintas indican diferencias significativas de acuerdo con la prueba de Tukey-Kramer (P ≤ 0.05).
La Figura 5 muestra que la tendencia decreciente del Ln_PIAB conforme el valor de la pendiente aumenta es notable. Se observa también una línea de regresión (azul) y otra de valor de referencia de crecimiento cero (rojo) en la escala de logaritmos, las cuales no se intersecan.

Figura 5 Regresión del logaritmo del porcentaje de incremento de área basal (Ln_PIAB) en especies de Pinus contra los valores promedio de la pendiente del sitio.
La Figura 6 muestra el logaritmo del PIAB en función del área basal del sitio (suma del área basal de cada individuo). La línea de regresión interseca al valor de crecimiento cero en 1.33 m2; después de dicho valor, el crecimiento promedio del sitio se nulifica por la competencia remanente. El promedio de crecimiento general de los 30 sitios es de 71 %.

Discusión
Crecimiento radial a nivel de árbol individual
Existe una respuesta significativa del crecimiento posterior a las actividades de cosecha en 47 % del arbolado residual; el 53 % restante muestra crecimiento no significativo, lo cual no necesariamente implica que no haya respuesta al tratamiento, sino que, en algunos subgrupos como G1.3 y G2.2, el crecimiento se presenta en un periodo limitado de solo cuatro a cinco años. Esto coincide con lo reportado por Kang et al. (2014) en un experimento de aclareos en P. koraiensis Siebold & Zucc. en Corea del Sur, donde el crecimiento se observó en un periodo de tres a cinco años después de los aclareos. Lo anterior sugiere que los individuos responden positivamente a la corta durante un lapso limitado y que el efecto se termina posiblemente por la competencia remanente en el sitio. Por otra parte, el subgrupo G1.2 mostró efecto positivo en el crecimiento de los individuos durante un periodo igual o mayor de 10 años después de la intervención (Cuadro 2); el mismo periodo fue reportado para Acer saccharum Marshall y Fagus grandifolia Ehrhart en Canadá (Jones et al., 2009). El patrón de crecimiento precosecha del grupo G1, significativamente menor al de los otros dos grupos, podría ser explicado principalmente por la competencia, lo que implica que los árboles suprimidos hayan respondido de mejor manera a la liberación del espacio (Figura 2A). Esto mismo sucedió con el subgrupo G1.2, ya que los individuos presentaron el menor crecimiento precosecha de todo el grupo y fueron los que más se beneficiaron con las cortas realizadas. Aunque no se tiene registro de investigaciones similares, estudios a nivel rodal han concluido que la respuesta en crecimiento se relaciona de forma positiva con la intensidad de la liberación (Espinosa et al., 1994; Kariuki, 2008), lo cual coincide con lo encontrado en este trabajo.
Las variables más importantes para explicar el PIAB fueron el diámetro del tocón, la competencia y el diámetro normal. Al respecto, Dolezâl, Ishii, Vetrova, Sumida, y Hara (2004) encontraron que la competencia y el tamaño fueron las variables de mayor importancia en los géneros Betula y Larix. La competencia es un factor de gran importancia para el crecimiento, tal como lo han demostrado diversos autores (Das, 2012; King, Davies, Nur-Supardi, & Tan, 2005; Pedersen, Bollandsås, Gobakken, & Næsset, 2012; Sánchez-Salguero et al., 2015).
El tamaño del tocón y la distancia al mismo son variables que están relacionadas con la intensidad de la competencia que ejercía el árbol derribado y que fue aprovechada por el arbolado residual en el sitio. En México, no se han reportado estudios con metodología similar al presente trabajo; sin embargo, algunos han evaluado el crecimiento en función de la competencia y sus resultados son comparables a los de esta investigación, ya que en ambos casos se determinan áreas de influencia de la competencia de los individuos en vida y derribados. Los 14.6 m de radio de influencia encontrados en este estudio superan lo reportado en otros trabajos con intervalos de 5.5 a 6.5 m (von Oheimb et al., 2011), 6.7 a 8.5 m (Fraver et al., 2014) y 7 m en otros (Looney et al., 2016). La diferencia en los resultados descritos puede deberse al efecto del tamaño (diámetro y altura) de los individuos evaluados; Fraver et al. (2014) no precisan datos, aunque describen que los árboles evaluados exceden de manera excepcional los 20 m de altura; von Oheimb et al. (2011) reportan 9.5 cm de diámetro y 11.5 m de altura, mientras que Looney et al. (2016) reportan un diámetro promedio de 22.2 ± 15 cm. Dicha medidas difieren en los individuos evaluados en el presente estudio, los cuales tuvieron diámetro de tocón de 48.4 ± 19.7 cm. Es posible que el radio de influencia de competencia obtenido en este trabajo difiera con los obtenidos en los estudios antes citados, debido a las diferencias de altura del arbolado, además de las discrepancias imputables a las variables usadas (i. e. diámetro del tocón a 0.3 m contrastado con el diámetro normal a 1.3 m en otros trabajos).
El resultado de la elevación del sitio contrasta con los estudios de Dang, Jiang, Zhang, y Zhang (2007) y Liu, Liang, Liu, y Camarero (2018), quienes sí encontraron influencia de la altitud sobre el crecimiento. La discrepancia con el presente estudio puede atribuirse al intervalo limitado de datos evaluados que tiene aproximadamente 300 m de diferencia, contra los 1 950 y 700 m de gradiente altitudinal en los estudios referidos.
Los resultados por especie varían en cada estudio; algunos coinciden con lo obtenido en el presente trabajo (Das, 2012; Huang, Stadt, Dawson, & Comeau, 2013), mientras que otros reportan variaciones significativas (King et al., 2005), lo cual posiblemente esté relacionado con la fisiología de las especies investigadas.
Crecimiento a nivel sitio
Los sitios mostraron un aumento en crecimiento promedio de 71 %, el cual es semejante a lo reportado por Forget et al. (2007), quienes obtuvieron incrementos de entre 45 y 50 % en rodales con aclareo de 29 % de intensidad de corta. Aunque los datos de incremento son comparables, el presente estudio acota el PIAB al área evaluada en los 30 sitios y no a nivel rodal.
Se identificó un umbral de 1.3 m2 de área basal residual como límite de competencia en el sitio (41.3 m2·ha-1), después del cual la liberación de competencia por efecto de cortas no tiene influencia en el crecimiento del arbolado residual. Debido a la falta de estudios comparables, se postula la hipótesis de que la competencia remanente impide el desarrollo promedio de los individuos residuales.
Los resultados negativos de crecimiento en exposición SE contrastan con los de Li et al. (2014), donde la exposición sur sí lo beneficia. La diferencia puede atribuirse al nivel latitudinal, ya que a 29° N (estudio citado), la radiación solar se recibe principalmente en la exposición sur y es lo que genera mejores condiciones de crecimiento; en cambio, a los 16° N (presente estudio), la radiación solar se recibe en exposiciones N y S dependiendo de la época del año.
La relación lineal negativa entre la pendiente del sitio y el crecimiento de los árboles (Figura 5) sugiere que este alcanza la línea de crecimiento cero aproximadamente en los 45º de pendiente. En un estudio similar se reportó un parámetro negativo débil (i. e. con valor de probabilidad cercano al valor α establecido), resultado de la regresión entre la pendiente y el crecimiento radial promedio de Quercus fabri Hance, aunque para Castanopsis fargesii Franch. resultó no significativo (von Oheimb et al., 2011). Estos resultados concuerdan con los reportados en el presente trabajo. La influencia negativa de la pendiente sobre el crecimiento posiblemente no se deba a la condición física, sino a las condiciones edafoclimáticas que genera, pues se ha encontrado que el aumento de la pendiente se relaciona con la disminución en la calidad del sitio (Wanshnong, Thakuria, Sangma, Ram, & Bora, 2013).
Conclusiones
Solo la mitad del arbolado residual aumentó su crecimiento como respuesta a las cortas de selección; los individuos suprimidos en precosecha fueron los más beneficiados con la liberación de espacio. La competencia fue la variable más influyente en el crecimiento, en conjunto con el tamaño del individuo evaluado. Este estudio presenta datos de tendencias, límites y umbrales que pueden ser útiles en la toma de decisiones del manejo forestal; por ejemplo, a partir del umbral de respuesta en crecimiento se puede modular el manejo de rodales, de acuerdo con el estrés hídrico y los cambios climáticos esperados. La metodología fue útil para evaluar el efecto de las cortas de selección, aunque es posible implementar mejoras en los criterios de selección de árboles competidores y en la elección de índices de competencia. Asimismo, es necesario establecer parcelas permanentes de muestreo para concluir de manera más sólida sobre los efectos de aclareos y cortas en los rodales analizados.
Agradecimientos
Los autores agradecen a las autoridades comunales de Santa María Lachixío, Oaxaca por el apoyo proporcionado en la planeación y colecta de datos en campo. Se agradece al CONACYT por el apoyo otorgado al primer autor a través de la beca 428413, indispensable para el desarrollo de la investigación. Al Colegio de Postgraduados por el financiamiento y apoyo logístico.
REFERENCIAS
Biondi, F. (1999). Comparing tree-ring chronologies and repeated timber inventories as forest monitoring tools. Ecological Applications, 9(1), 216-227. Retrieved from https://wolfweb.unr.edu/homepage/fbiondi/EcolApplic1999.pdf [ Links ]
Cambrón-Sandoval, V. H., Suzán-Azpiri, H., Vargas-Hernández, J. J., Sánchez-Vargas, N. M., & Sáenz-Romero, C. (2013). Estrategias de crecimiento y distribución de biomasa en Pinus pseudostrobus bajo diferentes condiciones de competencia. Revista Fitotecnia Mexicana, 36(1), 71-79. [ Links ]
Comisión Nacional Forestal (CONAFOR). (2001). Programa estratégico forestal para México 2025. Retrieved from http://era-mx.org/biblio/PEF_2025.pdf [ Links ]
Crecente-Campo, F., Vázquez-Gómez, E., Rodríguez-Soalleiro, R., & Diéguez-Aranda, U. (2007). Influencia de la competencia en el crecimiento individual de Pinus radiata D. Don en Galicia. Sociedad Española de Ciencias Forestales, 23, 167-174. Retrieved from http://secforestales.org/publicaciones/index.php/cuadernos_secf/article/view/9620/9538 [ Links ]
Dang, H., Jiang, M., Zhang, Q., & Zhang, Y. (2007). Growth responses of subalpine fir (Abies fargesii) to climate variability in the Qinling Mountain, China. Forest Ecology and Management, 240(1-3 ), 143-150. doi: 10.1016/j.foreco.2006.12.021 [ Links ]
Daniels, F., Burkhart, H. E., & Clason, T. R. (1986). A comparison of competition measures for predicting growth of loblolly pine trees. Canadian Journal of Forest Research,16(6), 1230-1237. doi: 10.1139/x86-218 [ Links ]
Das, A. (2012). The effect of size and competition on tree growth rate in old-growth coniferous forests. Canadian Journal of Forest Research, 42(11), 1983-1995. doi: 10.1139/x2012-142 [ Links ]
Dolezâl, J., Ishii, H., Vetrova, V. P., Sumida, A., & Hara, T. (2004). Tree growth and competition in a Betula platyphylla-Larix cajanderi post-fire forest in Central Kamchatka. Annals of Botany, 94(3), 333-343. doi: 10.1093/aob/mch149 [ Links ]
Espinosa, B. M., García, S. M., & Valeria, E. O. (1994). Efecto de intensidades diferentes de raleo en el crecimiento de un rodal de pino radiata. Bosque, 15(1), 55-65. doi: 10.4206/bosque.1994.v15n1-07 [ Links ]
Forget, E., Nolet, P., Doyon, F., Delagrange, S., & Jardon, Y. (2007). Ten-year response of northern hardwood stands to commercial selection cutting in southern Quebec, Canada. Forest Ecology and Management, 242(2-3), 764-765. doi: 10.1016/j.foreco.2007.02.010 [ Links ]
Fraver, S., D’Amato, A. W., Bradford, J. B., Jonsson, B. G., Jönsson, M., & Esseen, P-A. (2014). Tree growth and competition in an old-growth Picea abies forest of boreal Sweden: influence of tree spatial patterning. Journal of Vegetation Science, 25(2), 374-385. doi: 10.1111/jvs.12096 [ Links ]
Grissino-Mayer, H. D. (2001). Evaluating cross dating accuracy: A manual and tutorial for the computer program COFECHA. Tree-Ring Research 57(2), 205-221. Retrieved from https://www.researchgate.net/publication/244461190_Evaluating_crossdating_accuracy_a_manual_and_tutorial_for_the_computer_program_COFECHA [ Links ]
Hegyi, F. (1974). A simulation model for managing jack pine stands. In J. Fries (Ed.), Growth models for tree and stand simulation (pp. 74-90). Stockholm, Sweden: Royal Coll. of For. [ Links ]
Hernández-Díaz, J. C., Corral-Rivas, J. J., Quiñonez-Chávez, A., Bacon-Sobbe, J. R., & Vargas-Larreta, B. (2008). Evaluación del manejo forestal regular e irregular en bosques de la Sierra Madre Occidental. Madera y Bosques, 14(3), 25-41. Retrieved from http://www.scielo.org.mx/pdf/mb/v14n3/v14n3a2.pdf [ Links ]
Hocking, R. R. (1976). The analysis and selection of variables in linear regression. Biometrics, 32(1), 1-49. doi: 10.2307/2529336 [ Links ]
Huang, J-G., Stadt, K. J., Dawson, A., & Comeau, P. G. (2013). Modelling growth-competition relationships in trembling aspen and white spruce mixed boreal forests of western Canada. PloS ONE, 8(10), 1-14. doi: 10.1371/journal.pone.0077607 [ Links ]
Jones, T. A., Domke, G. M., & Thomas, S. C. (2009). Canopy tree growth responses following selection harvest in seven species varying in shade tolerance. Canadian Journal of Forest Research, 39(2), 430-440. doi: 10.1139/X08-186 [ Links ]
Jones, T. A., & Thomas, S. C. (2004). The time course of diameter increment responses to selection harvests in Acer saccharum. Canadian Journal of Forest Research, 34, 1525-1533. doi: 10.1139/X04-034 [ Links ]
Kang, J-S., Shibuya, M., & Shin, C-S. (2014). The effect of forest-thinning works on tree growth and forest environment. Forest Science and Technology, 10(1), 33-39. doi: 10.1080/21580103.2013.821958 [ Links ]
Kariuki, M. (2008). Modelling the impacts of various thinning intensities on tree growth and survival in a mixed species eucalypt forest in central Gippsland, Victoria, Australia. Forest Ecology and Management, 256(12), 2007-2017. doi: 10.1016/j.foreco.2008.07.035 [ Links ]
King, D. A., Davies, S. J., Nur-Supardi, M. N., & Tan, S. (2005). Tree growth is related to light interception and wood density in two mixed dipterocarp forests of Malaysia. Functional Ecology, 19(3), 445-453. doi: 10.1111/j.1365-2435.2005.00982.x [ Links ]
Li, Y., Härdtle, W., Bruelheide, H., Nadrowski, K., Scholten, T., von Wehrden, H., & von Oheimb, G. (2014). Site and neighborhood effects on growth of tree saplings in subtropical plantations (China). Forest Ecology and Management, 327, 118-127. doi: 10.1016/j.foreco.2014.04.039 [ Links ]
Liu, B., Liang, E., Liu, K., & Camarero, J. J. (2018). Species- and elevation-dependent growth responses to climate warming of Mountain Forests in the Qinling Mountains, Central China. Forests, 9(5), 1-11. doi: 10.3390/f9050248 [ Links ]
Looney, C. E., D’Amato, A. W., Fraver, S., Palik, B. J., & Reinikainen, M. R. (2016). Examining the influences of tree-to-tree competition and climate on size-growth relationships in hydric, multi-aged Fraxinus nigra stands. Forest Ecology and Management, 375, 238-248. doi: 10.1016/j.foreco.2016.05.050 [ Links ]
Martín-Benito, D., Del Rio, M., Heinrich, I., Helle, G., & Cañellas, I. (2010). Response of climate-growth relationships and water use efficiency to thinning in a Pinus nigra afforestation. Forest Ecology and Management, 259(5), 967-975. doi: 10.1016/j.foreco.2009.12.001 [ Links ]
Martin, L., & Ek, A. R. (1984). A comparison of competition measures and growth models for predicting plantation red pine diameter growth. Forest Science, 30(3), 731-743. doi: 10.1093/forestscience/30.3.731 [ Links ]
Pedersen, R. O., Bollandsås, O. M., Gobakken, T., & Næsset, E. (2012). Deriving individual tree competition indices from airborne laser scanning. Forest Ecology and Management, 280, 150-165. doi: 10.1016/j.foreco.2013.07.040 [ Links ]
Powers, M. D., Pregitzer, K. S., Palik, B. J., & Webster, C. R. (2009). Wood d13C, d18O and radial growth responses of residual red pine to variable retention harvesting. Tree Physiology, 30, 326-334. doi: 10.1093/treephys/tpp119 [ Links ]
Regent Instruments Inc. (2009). WinDENDRO™ for tree-ring analysis. Quebec, Canada: Author. [ Links ]
Rodríguez-Ortiz, G., González-Hernández, V. A., Aldrete, A., De los Santos-Posadas, H. M., Gómez-Guerrero, A., & Fierros-González, A. M. (2011). Modelos para estimar crecimiento y eficiencia de crecimiento en plantaciones de Pinus patula en respuesta al aclareo. Revista Fitotecnia Mexicana, 34(3), 205-212. Retrieved from http://www.scielo.org.mx/pdf/rfm/v34n3/v34n3a12.pdf [ Links ]
Sánchez-Salguero, R., Linares, J. C., Camarero, J. J., Madrigal-González, J., Hevia, A., Sánchez-Miranda, A., …Rigling, A. (2015). Disentangling the effects of competition and climate on individual tree growth: A retrospective and dynamic approach in Scots pine. Forest Ecology and Management, 358, 12-25. doi: 10.1016/j.foreco.2015.08.034 [ Links ]
Statistical Analysis Systems Institute (2014). The SAS system for windows, release 9.4. Cary, NC, USA: Author. [ Links ]
Tomé, M., & Burkhart, H. E. (1989). Distance-dependent competition measures for predicting growth of individual trees. Forest Science, 35(3), 816-831. Retrieved from https://www.researchgate.net/publication/233645214_Distance-Dependent_Competition_Measures_for_Predicting_Growth_of_Individual_Trees [ Links ]
Torres-Rojo, J. M. (2000). Sostenibilidad del volumen de cosecha calculado con el Método Mexicano de Ordenación de Montes. Madera y Bosques, 6(2), 57-72. doi: 10.21829/myb.2000.621335 [ Links ]
Von Oheimb, G., Lang, A. C., Bruelheide, H., Forrester, D. I., Wäschea, I., Yu, M., & Härdtlea, W. (2011). Individual-tree radial growth in a subtropical broad-leaved forest: The role of local neighbourhood competition. Forest Ecology and Management, 261(3), 499-507. doi: 10.1016/j.foreco.2010.10.035 [ Links ]
Wanshnong, R. K., Thakuria, D., Sangma, C. B., Ram, V., & Bora, P. K. (2013). Influence of hill slope on biological pools of carbon, nitrogen, and phosphorus in acidic alfisols of citrus orchard. CATENA, 111, 1-8. doi: 10.1016/j.catena.2013.07.009 [ Links ]
Recibido: 02 de Septiembre de 2018; Aprobado: 29 de Octubre de 2019
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
