










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.25 no.2 Chapingo may./ago. 2019 Epub 19-Feb-2021
https://doi.org/10.5154/r.rchscfa.2018.12.096
Artículo científico
Aporte de nutrientes vía precipitación incidente, directa y escurrimiento fustal en especies de matorral en el noreste de México
1Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales. Carretera Nacional núm. 85, km 145. C. P. 67700. Linares, Nuevo León, México.
Introducción:
La precipitación es un factor importante en el aporte de nutrientes a los ecosistemas. En algunos casos, mediante la lluvia pueden agregarse elementos que por las condiciones del área no están disponibles.
Objetivo:
Comparar el aporte nutrimental en la precipitación incidente (Pi) y en el agua de lluvia que llega al suelo por precipitación directa (Pd) y escurrimiento fustal (Ef) de especies de matorral.
Materiales y métodos:
Se evaluaron 41 eventos de lluvia en el noreste de México en el periodo septiembre 2016-septiembre 2017. Se registraron los volúmenes de precipitación y se analizó su composición química: pH, conductividad eléctrica y aporte de macro (Ca, Mg y K) y micronutrientes (Fe, Mn, Zn y Cu).
Resultados y discusión:
La Pi acumuló 508.97 mm. La Pd es la que deposita mayor cantidad de nutrientes al suelo. El aporte neto (Pd+Ef) indicó que el Fe fue el micronutriente más depositado con 2 938.29 g·ha-1·año-1, de los cuales Casimiroa greggii (S. Watson) F. Chiang aportó 31.59 %. El macronutriente con mayor presencia fue el Ca con 319.31 kg·ha-1·año-1, siendo Acacia farnesiana (L.) Willd. la que logró mayor flujo del mineral (22.66 %). El pH y conductividad eléctrica fueron similares (Kruskal-Wallis, P > 0.05) en los tres tipos de precipitación.
Conclusión:
La química de la precipitación se modifica después de pasar el dosel de cada especie, ya sea para enriquecer o remover las concentraciones de nutrientes, jugando un papel importante en los ciclos biogeoquímicos del ecosistema.
Palabras clave: Casimiroa greggii; Acacia farnesiana; matorral espinoso; pH; conductividad eléctrica
Introduction:
Precipitation is an important factor in the input of nutrients to ecosystems. In some cases, rainfall can add elements that are not available due to area conditions.
Objective:
To compare the nutritional contribution in gross rainfall (Pi) and rainwater reaching the soil by throughfall (Pd) and stemflow (Ef) of scrubland species.
Materials and methods:
A total of 41 rainfall events were evaluated in northeastern Mexico in the period September 2016-September 2017. Precipitation volumes were recorded and their chemical composition were analyzed: pH, electrical conductivity and contribution of macronutrients (Ca, Mg and K) and micronutrients (Fe, Mn, Zn and Cu).
Results and discussion:
Pi accumulated 508.97 mm. Pd is the one that deposits more nutrients to the soil. The net contribution (Pd+Ef) indicated that Fe was the most deposited micronutrient with 2 938.29 g·ha-1·year-1, of which Casimiroa greggii (S. Watson) F. Chiang contributed 31.59 %. The macronutrient with greater presence was Ca with 319.31 kg·ha-1·year-1, being Acacia farnesiana (L.) Willd. the one that achieved greater flow of the mineral (22.66 %). pH and electrical conductivity were similar (Kruskal-Wallis, P > 0.05) in the three types of rainfall.
Conclusion:
The precipitation chemistry is modified after passing the canopy of each species, either to enrich or remove nutrient concentrations, playing an important role in the biogeochemical cycles of the ecosystem.
Keywords: Casimiroa greggii; Acacia farnesiana; thornscrub; pH; electrical conductivity
Introducción
La deposición húmeda influye en el ciclo de nutrientes y en la distribución de contaminantes en los ecosistemas. Muchas de las sustancias que llegan a la atmósfera reaccionan entre sí y con el agua; los productos resultantes de esas interacciones son arrastrados por las lluvias como soluciones o partículas insolubles (Duarte, Gioda, Ziolli, & Duó, 2013), provocando desequilibrios en la composición y propiedades del suelo.
La precipitación en los ecosistemas forestales se divide en precipitación directa, escurrimiento fustal y pérdida de intercepción; clasificación que es importante en los estudios de hidrología forestal (Lida, Tanaka, & Sugita, 2005; Yáñez, Cantú, González, & Uvalle, 2014). La precipitación directa se refiere a la lluvia debajo del dosel; es decir, la cantidad de agua que fluye por el dosel a través de claros o por goteo de las hojas y ramas (Cantú & González, 2005). La lluvia interceptada es la parte de la precipitación retenida por el dosel de copas que no alcanza el piso del bosque y es regresada a la atmósfera por el proceso de evaporación, también denominada precipitación incidente (Taghi, Attarod, & Bayramzadeh, 2013; Yáñez et al., 2014). Finalmente, el escurrimiento fustal corresponde a la fracción de la lluvia que llega al piso del bosque, deslizándose por los fustes y ramas de los árboles, la cual es ocasionada por la precipitación interceptada por los componentes del dosel de copas (Staelens, De-Schrijver, Verheyen, & Verhoest, 2008). La precipitación neta es la cantidad de lluvia que llega al suelo del bosque a través de la precipitación directa y el escurrimiento fustal (André, Jonard, & Ponette, 2008; Levia & Herwitz, 2005).
La presente investigación tuvo como objetivo comparar las concentraciones de micro y macronutrientes en la precipitación incidente y en el agua de lluvia que llega al suelo por precipitación directa y escurrimiento fustal de especies de matorral; además, se analizaron las variaciones del pH y conductividad eléctrica. Cabe destacar que no existen antecedentes en lo que refiere al estudio de la composición química de la precipitación dentro de este ecosistema, lo que podría generar información importante que puede ser usada en las decisiones y criterios técnicos para lograr la sustentabilidad de los recursos naturales.
Materiales y métodos
Área de estudio
El estudio se llevó a cabo en el Campus Experimental de la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León (24° 47' N; 99° 32' O), localizado en Linares, Nuevo León. El sitio cuenta con una superficie de 500 ha y una elevación de 350 m. La temperatura media mensual varía de 15 a 22 °C y la precipitación media anual es aproximadamente de 800 mm. El principal tipo de vegetación es denominado matorral espinoso tamaulipeco también conocido como MET. Los suelos dominantes son vertisoles profundos, gris oscuros, limo-grisáceos, limo-arcillosos con montmorillonita (González, Ramírez, Cantú, Gómez, & Uvalle, 2010).
Medición de la precipitación incidente, precipitación directa y escurrimiento fustal
La precipitación incidente (Pi) se midió de acuerdo con la metodología empleada por Cantú y González (2001), la cual se basa en el establecimiento, al azar, de cuatro canaletas de PVC en forma de U en un área abierta sin árboles y contigua a las parcelas experimentales. Cada canaleta representó un área de colecta de 0.1 m2 (10 cm de ancho x 100 cm de largo), la cual se colocó a 1 m de altura sobre la superficie del suelo. Las canaletas se conectaron a recipientes de 20 L por medio de mangueras y se cubrieron con una malla para evitar obstrucción y contaminación de las muestras de precipitación.
Las muestras de precipitación directa (Pd) se recolectaron con el mismo tipo y número de canaletas ya descritas, pero ubicadas debajo del dosel de plantaciones de Acacia farnesiana (L.) Willd. (huizache), Condalia hookeri M. C. Johnst (palo Brasil), Leucaena leucocephala (Lam.) de Wit (huaje), Casimiroa greggii (S. Watson) F. Chiang (chapote amarillo), y de una parcela compuesta por un dosel heterogéneo característico del MET. El Cuadro 1 muestra las características dasométricas promedio de cada especie empleada.
Cuadro 1 Características dasométricas promedio de las especies bajo estudio en el matorral espinoso tamaulipeco.
| Tipo de cobertura | DAP (cm) | Altura (m) | Área de copa (m2) |
|---|---|---|---|
| Acacia farnesiana (L.) Willd. | 28.3 | 5.3 | 42.2 |
| Condalia hookeri M. C. Johnst | 9.5 | 8.5 | 13.5 |
| Casimiroa greggii (S. Watson) F. Chiang | 17.0 | 7.1 | 22.6 |
| Leucaena leucocephala (Lam.) de Wit. | 7.2 | 9.2 | 4.0 |
| Matorral espinoso tamaulipeco | 7.0 | 7.5 | 4.5 |
El componente de escurrimiento fustal (Ef) se midió en cuatro árboles de cada uno de los doseles de A. farnesiana, C. hookeri y C. greggii únicamente, debido a que cumplieron con las dimensiones diamétricas (>15 cm) necesarias para colocar el colector en espiral. El colector es una manguera perforada con una longitud de 2 m aproximadamente, ajustada y sellada con silicón en el fuste, para interceptar el agua que escurre alrededor del tronco y recolectar la muestra en un recipiente de 20 L.
Análisis químico
Las muestras de agua de lluvia se llevaron al laboratorio, para determinar el pH y la conductividad eléctrica (µS·cm-1), usando un electrodo con un potenciómetro-conductivímetro (CORNING, modelo 542, EUA). Después, las muestras se filtraron con papel Whatman número 1 y se refrigeraron a 4 °C con el fin de reducir la actividad microbiana hasta su análisis posterior de nutrientes.
Análisis de nutrientes
Las concentraciones de micronutrientes (Fe, Zn, Mn y Cu) y macronutrientes (K, Ca y Mg), esenciales para el desarrollo de las plantas, se determinaron en Pi, Pd y Ef. Las muestras se analizaron con el uso de un espectrofotómetro de absorción atómica (Varían, modelo SpectrAA-200, EUA). El contenido de nitrógeno (N) no se evaluó, debido a la falta de capacidad operativa para su determinación. En el caso de los macronutrientes fue necesario hacer diluciones para determinar la absorción de radiación a diferentes longitudes de onda según el elemento analizado, utilizando los siguientes supresores con el fin de evitar la ionización durante el análisis: Calcio (Ca), soluciones 1:5 con cloruro de potasio (KCl); magnesio (Mg), soluciones 1:5 con cloruro de lantano (LaCl3); y potasio (K), soluciones 1:5 con cloruro de cesio (CsCl).
Aporte de nutrientes
El aporte de macro y micronutrientes por evento se determinó en la Pi, Pd y Ef de las coberturas. Los flujos de nutrientes se calcularon multiplicando el contenido medio de nutrientes (mg·L-1) obtenido por el espectrofotómetro, por el volumen (L) correspondiente de agua de lluvia recolectada en Pi y Pd, entre el área ocupada por la canaleta (0.10 m2). Para el Ef, el área de la copa de cada árbol se consideró como la superficie receptora de la precipitación, realizando con ello las trasformaciones necesarias para convertir a unidades de mg·m-2. El promedio de las cuatro repeticiones correspondientes a cada muestra de Pi, Pd y Ef recolectada se calculó por evento; para determinar el aporte anual, se sumó la totalidad de los promedios registrados.
El aporte neto de nutrientes se calculó mediante la suma de las concentraciones de Pd y Ef (en caso de registrarse), de acuerdo con el mismo principio que utilizó Béjar et al. (2018). El aporte neto se registró para las especies A. farnesiana, C. greggi y C. hookeri; para el caso deL. leucocephala y MET no se consideró esta suma por falta del componente escurrimiento fustal, por lo que el valor neto fue el encontrado en sus Pd. Cabe señalar que la adición de Ef no causó un cambio significativo en los contenidos de los nutrientes evaluados.
Análisis estadístico
Los datos se analizaron a través del paquete estadístico SPSS versión 22 (International Business Machines [IBM], 2013). Se aplicó la prueba de normalidad y bondad de ajuste de Kolmogorov-Smirnov, la cual rechazó la hipótesis de normalidad de los datos, por lo que se procedió a aplicar pruebas no paramétricas (Kruskal-Wallis), para establecer las diferencias estadísticas significativas (P ≤ 0.05) en el aporte de nutrientes de Pd y Ef por evento y por acumulado anual entre los tipos de dosel vegetal. Asimismo, se aplicaron pruebas de comparación no paramétricas (Kruskal-Wallis), para determinar diferencias significativas (P < 0.05) en los valores de pH y conductividad eléctrica (CE) en la Pi, Pd y Ef de las coberturas por eventos registrados.
Resultados y discusión
Precipitación incidente
En el periodo septiembre 2016-septiembre 2017 se registraron 41 eventos de lluvia (Figura 1), en los cuales se acumularon 508.97 mm. Los meses de noviembre y mayo presentaron mayor cantidad de eventos, cada uno con cinco registros. Estacionalmente, la mayor precipitación se registró en primavera con 228.6 mm; en verano, otoño e invierno, la precipitación fue 152.39, 104.89 y 23.03 mm, respectivamente.

Aporte de nutrientes vía precipitación incidente
Las concentraciones de nutrientes en la Pi reflejan que el Ca (76.25 kg·ha-1·año-1) es el macronutriente que ingresa en mayor cantidad al suelo, siguiendo en orden descendente el K (20.40 kg·ha-1·año-1) y el Mg (9.39 kg·ha-1·año-1). En cuanto a los micronutrientes, las contribuciones más altas fueron de Fe con 521.55 g·ha-1·año-1, seguido de Mn, Zn y Cu con 180.21, 130.16 y 29.10 g·ha-1·año-1, respectivamente (Cuadro 2). Las tendencias mostradas por los flujos de macronutrientes (Ca>K>Mg) vía Pi coinciden con los obtenidos por Béjar et al. (2018) y Zhang, Wang, Pan, y Hu (2016).
Cuadro 2 Aporte de nutrientes vía precipitación incidente.
| Micronutriente | Aporte (g·ha-1·año-1) | Macronutriente | Aporte (kg·ha-1·año-1) |
|---|---|---|---|
| Fe | 521.55 | Ca | 76.25 |
| Mn | 180.21 | K | 20.40 |
| Zn | 130.16 | Mg | 9.39 |
| Cu | 29.10 | ||
| Total | 861.02 | 106.04 |
Los contenidos altos de Ca pueden estar relacionados con las condiciones del ecosistema, ya que los suelos están desarrollados sobre rocas de origen calcáreo y, por ende, el Ca se halla en mayor movilización sobre el ambiente, a diferencia de los micronutrientes Mn, Zn y Cu. Por otra parte, Segovia et al. (2009) hacen referencia a que el Fe es el cuarto elemento más abundante sobre la corteza terrestre, el cual sigue la vía atmosférica como una de sus principales rutas de acceso, concordando con los resultados encontrados en la presente investigación.
Aporte de nutrientes vía precipitación directa
Los resultados mostraron diferencias en el aporte de nutrientes por evento en al menos una de las coberturas arbóreas, a partir del valor de la asintótica bilateral (P ≤ 0.05). De acuerdo con el Cuadro 3, el aporte de los macronutrientes Ca, K y Mn fue diferente entre las especies en 17, 20 y nueve eventos de lluvia, respectivamente. Con respecto a los micronutrientes, el Cu presentó diferencias significativas en seis de los 41 eventos analizados, mientras que Fe, Mn y Zn en 18, 16 y 16 eventos, respectivamente. Posiblemente estas variaciones temporales estén relacionadas con la adición de partículas de suelo y polvos atmosféricos, y con la composición de la biomasa de las especies en fechas diferentes. En los eventos 26, 27, 28 y 37 en Cu y 24 y 34 en Mg no se muestra el nivel de significación de la prueba estadística, debido a que solo una de las especies evaluadas presentó acumulación de dichos nutrientes, limitando la comparación interespecífica. De acuerdo con los estadísticos de la prueba de Kruskal-Wallis, no existen diferencias (P > 0.05) entre los aportes anuales de micro y macronutrientes entre los cinco doseles arbóreos analizados.
Cuadro 3 Significancias estadísticas (valores P) de la comparación del aporte de nutrientes en la precipitación directa de cinco doseles arbóreos (Acacia farnesiana, Condalia hookeri, Leucaena leucocephala, Casimiroa greggii y matorral espinoso) analizados por evento.
| Evento | Fe | Mn | Zn | Cu | Ca | K | Mg | Precipitación (mm) | Fecha |
|---|---|---|---|---|---|---|---|---|---|
| P | P | P | P | P | P | P | |||
| 1 | 0.01* | 0.52 | 0.05 | 0.08 | 0.04* | 0.19 | 0.04* | 3.83 | 23/09/2016 |
| 2 | 0.06 | 0.02* | 0.05* | 0.40 | 0.58 | 0.05 | 0.61 | 15.43 | 26/09/2016 |
| 3 | 0.04* | 0.02* | 0.05 | 0.03* | 0.01* | 0.11 | 0.09 | 22.04 | 27/09/2016 |
| 4 | 0.01* | 0.01* | 0.76 | 0.01* | 0.32 | 0.01* | 0.07 | 6.60 | 01/10/2016 |
| 5 | 0.02* | 0.07 | 0.33 | 0.26 | 0.01* | 0.37 | 0.77 | 2.97 | 02/10/2016 |
| 6 | 0.62 | 0.31 | 0.17 | 0.85 | 0.23 | 0.69 | 0.53 | 47.32 | 13/10/2016 |
| 7 | 0.44 | 0.03* | 0.04* | 0.04* | 0.07 | 0.03* | 0.27 | 12.76 | 22/10/2016 |
| 8 | 0.04* | 0.31 | 0.49 | 0.05* | 0.02* | 0.76 | 0.02* | 0.69 | 04/11/2016 |
| 9 | 0.05* | 0.01* | 0.39 | 0.07 | 0.01* | 0.01* | 0.12 | 15.54 | 05/11/2016 |
| 10 | 0.22 | 0.22 | 0.17 | 0.55 | 0.14 | 0.13 | 0.52 | 0.59 | 07/11/2016 |
| 11 | 0.03* | 0.01* | 0.02* | 0.26 | 0.03* | 0.55 | 0.12 | 8.47 | 09/11/2016 |
| 12 | 0.03* | 0.02* | 0.53 | 0.73 | 0.09 | 0.18 | 0.02* | 16.17 | 14/11/2016 |
| 13 | 0.76 | 0.07 | 0.39 | 0.24 | 0.69 | 0.01* | 0.85 | 2.33 | 05/12/2016 |
| 14 | 0.32 | 0.27 | 0.69 | 0.54 | 0.24 | 0.01* | 0.64 | 3.22 | 09/12/2016 |
| 15 | 0.76 | 0.03* | 0.01* | 0.66 | 0.67 | 0.31 | 0.15 | 0.78 | 28/12/2017 |
| 16 | 0.01* | 0.01* | 0.01* | 0.01* | 0.01* | 0.01* | 0.01* | 0.72 | 06/01/2017 |
| 17 | 0.13 | 0.02* | 0.41 | 0.15 | 0.54 | 0.07 | 0.10 | 0.68 | 07/01/2017 |
| 18 | 0.17 | 0.01* | 0.01* | 0.01* | 0.01* | 0.01* | 0.01* | 11.28 | 28/01/2017 |
| 19 | 0.48 | 0.36 | 0.12 | 0.73 | 0.02* | 0.01* | 0.95 | 3.03 | 04/02/2017 |
| 20 | 0.13 | 0.04* | 0.02* | 0.22 | 0.18 | 0.01* | 0.39 | 0.99 | 25/02/2017 |
| 21 | 0.11 | 0.07 | 0.02* | 0.17 | 0.03* | 0.05* | 0.53 | 7.13 | 05/03/2017 |
| 22 | 0.02* | 0.31 | 0.01* | 0.54 | 0.07 | 0.23 | 0.52 | 7.75 | 08/03/2017 |
| 23 | 0.47 | 0.08 | 0.70 | 0.41 | 0.13 | 0.04* | 0.24 | 11.00 | 10/03/2017 |
| 24 | 0.75 | 0.19 | 0.01* | 0.19 | 0.15 | 0.01* | - | 19.88 | 11/03/2017 |
| 25 | 0.61 | 0.05 | 0.01* | 0.66 | 0.02* | 0.03* | 0.25 | 19.00 | 18/03/2017 |
| 26 | 0.02* | 0.53 | 0.48 | - | 0.17 | 0.01* | 0.93 | 56.50 | 12/04/2017 |
| 27 | 0.43 | 0.08 | 0.29 | - | 0.67 | 0.01* | 0.02* | 3.50 | 23/04/2017 |
| 28 | 0.03* | 0.20 | 0.06 | - | 0.15 | 0.01* | 0.05* | 3.00 | 04/05/2017 |
| 29 | 0.11 | 0.06 | 0.35 | 0.37 | 0.03* | 0.01* | 0.02* | 0.88 | 07/05/2017 |
| 30 | 0.06 | 0.11 | 0.20 | 0.44 | 0.01* | 0.01* | 0.11 | 1.93 | 11/05/2017 |
| 31 | 0.01* | 0.01* | 0.40 | 0.42 | 0.05* | 0.37 | 0.11 | 11.50 | 12/05/2017 |
| 32 | 0.01* | 0.01* | 0.53 | 0.47 | 0.05 | 0.06 | 0.34 | 0.99 | 23/05/2017 |
| 33 | 0.01* | 0.01* | 0.07 | 0.81 | 0.38 | 0.11 | 0.72 | 5.13 | 24/05/2017 |
| 34 | 0.01* | 0.01* | 0.05* | 0.09 | 0.07 | 0.22 | - | 80.50 | 30/05/2017 |
| 35 | 0.01* | 0.11 | 0.01* | 0.19 | 0.60 | 0.07 | 0.03* | 0.62 | 20/06/2017 |
| 36 | 0.01* | 0.11 | 0.01* | 0.09 | 0.01* | 0.08 | 0.10 | 10.90 | 26/06/2017 |
| 37 | 0.05 | 0.06 | 0.33 | - | 0.18 | 0.39 | 0.08 | 53.38 | 28/06/2017 |
| 38 | 0.88 | 0.57 | 0.47 | 0.17 | 0.10 | 0.05 | 0.36 | 10.28 | 07/07/2017 |
| 39 | 0.15 | 0.70 | 0.84 | 0.47 | 0.03* | 0.01* | 0.06 | 1.50 | 09/07/2017 |
| 40 | 0.12 | 0.21 | 0.01* | 0.55 | 0.04* | 0.01* | 0.15 | 23.13 | 04/08/2017 |
| 41 | 0.27 | 0.69 | 0.05* | 0.54 | 0.06 | 0.25 | 0.30 | 5.10 | 05/08/2017 |
*Valores con diferencias significativas (Kruskal-Wallis P ≤ 0.05) en el aporte de nutrientes entre especies.
La Figura 2 muestra que A. farnesiana fue la especie con mayores aportes de Ca y K (72.33 y 33.10 kg·ha-1·año-1, respectivamente) en la Pd, mientras que el contenido más alto de Mg se encontró en la parcela correspondiente a MET con 13.55 kg·ha-1·año-1. En general, una parte del aporte de Ca vía lluvia es retenido por todos los doseles arbóreos, mientras que el Mg es ligeramente retenido solo en C. hookeri y L. leucocephala con aportes de 9.03 y 8.31 kg·ha-1·año-1 respecto a los arrojados por Pi (9.62 kg·ha-1·año-1). Esto coincide con Béjar (2018), Jiménez, Calvo, y Arias (2006), y Jaramillo (2003), quienes describen una remoción de estos nutrientes con el paso de la precipitación por los doseles.

Figura 2 Aporte de macronutrientes vía precipitación directa para cada dosel vegetal y vía precipitación incidente (Pi).
La concentración de K fue enriquecida significativamente por los cinco doseles evaluados. Los enriquecimientos promedios mostraron 1.38 veces más que el registrado en Pi; por lo tanto, K es un nutriente móvil en las plantas y se lixivia más fácilmente. Este enriquecimiento puede ser efecto de las interacciones químicas entre la lluvia, la deposición seca, las exudaciones y la lixiviación del dosel (Návar, González, & González, 2009). Otros estudios han manifestado este mismo comportamiento; por ejemplo, Jaramillo (2003) encontró que el K incrementó al pasar por la parte aérea hasta 9.6 veces más que la estimada en el agua de lluvia, variación provocada principalmente por el lavado foliar; Shen, Ren, Jenerette, Hui, y Ren (2013) encontraron incrementos de aproximadamente 20 veces en Pd en plantaciones de Dimocarpus longan Lour y Acacia mangium Wild.
De acuerdo con el Cuadro 4, la vía Pd siguió una misma tendencia en el aporte de micronutrientes para todas las coberturas analizadas: Fe>Zn>Mn>Cu. En el dosel de C. greggii se depositó la mayor cantidad de Fe con 889.53 g·ha-1·año-1, en el MET se cuantificaron los valores más altos de Zn y Mn con 203.38 y 118.93 g·ha-1·año-1, respectivamente y, por último, en A. farnesiana se encontró la mayor cantidad de Cu con 65.03 g·ha-1·año-1.
Cuadro 4 Aporte anual de micronutrientes vía precipitación directa por tipo de dosel vegetal.
| Tipo de cobertura | Aporte (g·ha-1·año-1) | |||
|---|---|---|---|---|
| Fe | Mn | Zn | Cu | |
| Acacia farnesiana | 655.96 | 102.46 | 150.8 | 65.03 |
| Condalia hookeri | 613.43 | 55.5 | 130.7 | 58.31 |
| Casimiroa greggii | 889.53 | 73.13 | 160.97 | 45.1 |
| Leucaena leucocephala | 394.18 | 101.86 | 115.43 | 26.59 |
| Matorral espinoso tamaulipeco | 342.11 | 118.93 | 203.38 | 31.64 |
| Total | 2 895.21 | 451.88 | 761.28 | 226.67 |
Aporte de nutrientes vía escurrimiento fustal
El aporte de calcio en la vía Ef de las tres especies analizadas mostró diferencias significativas en nueve de los 24 eventos de lluvia analizados, mientras que K y Mg presentaron diferencias en seis eventos. En cuanto a Cu, Fe, Mn y Zn se encontraron diferencias en cuatro, seis, cinco y 10 eventos de lluvia, respectivamente. En el caso del Cu, los eventos 14, 23, 24, 26 y 27 no se compararon debido a la contribución nula de dos de las especies evaluadas para esta vía, repitiéndose esta situación en el evento 34 en Mg (Cuadro 5).
Cuadro 5 Significancias estadísticas (valores P) de la comparación del aporte de nutrientes en el escurrimiento fustal de tres coberturas (Acacia farnesiana, Condalia hookeri, Casimiroa greggii) analizadas por evento.
| Evento | Cu | Fe | Mn | Zn | Ca | K | Mg |
|---|---|---|---|---|---|---|---|
| P | P | P | P | P | P | P | |
| 13 | 0.18 | 0.08 | 0.10 | 0.08 | 0.08 | 0.08 | 0.08 |
| 14 | - | 1.00 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| 15 | 0.18 | 0.18 | 0.18 | 0.18 | 0.18 | 0.32 | 0.22 |
| 16 | 0.49 | 0.08 | 0.20 | 0.34 | 0.30 | 0.57 | 0.25 |
| 17 | 0.08 | 0.04* | 0.08 | 0.30 | 0.20 | 0.05* | 0.61 |
| 18 | 0.08 | 0.03* | 0.02* | 0.02* | 0.02* | 0.02* | 0.04* |
| 19 | 0.23 | 0.06 | 0.03* | 0.03* | 0.04* | 0.23 | 0.04* |
| 21 | 0.02* | 0.02* | 0.08 | 0.02* | 0.02* | 0.04* | 0.03* |
| 22 | 0.22 | 0.03* | 0.03* | 0.03* | 0.03* | 0.03* | 0.03* |
| 23 | - | 0.62 | 0.06 | 0.03* | 0.74 | 0.07 | 0.72 |
| 24 | - | 0.15 | 0.04* | 0.04* | 0.02* | 0.03* | 0.12 |
| 25 | 0.32 | 0.20 | 0.02* | 0.04* | 0.11 | 0.12 | 0.26 |
| 26 | - | 0.38 | 0.41 | 0.11 | 0.63 | 0.38 | 0.74 |
| 27 | - | 0.86 | 0.34 | 0.08 | 0.86 | 0.34 | 0.50 |
| 28 | 0.22 | 0.08 | 0.23 | 0.09 | 0.09 | 0.08 | 0.20 |
| 30 | 0.21 | 0.11 | 0.18 | 0.34 | 0.04* | 0.15 | 0.41 |
| 31 | 0.11 | 0.02* | 0.02* | 0.10 | 0.03* | 0.12 | 0.18 |
| 33 | 0.04* | 0.03* | 0.59 | 0.55 | 0.41 | 0.30 | 0.04* |
| 34 | 0.02* | 0.32 | 0.16 | 0.05* | 0.30 | 0.15 | - |
| 36 | 0.37 | 0.07 | 0.20 | 0.19 | 0.07 | 0.08 | 0.38 |
| 37 | 0.30 | 0.57 | 0.90 | 0.75 | 0.44 | 0.93 | 0.17 |
| 38 | 0.70 | 0.44 | 0.68 | 0.42 | 0.53 | 0.45 | 0.82 |
| 40 | 0.03* | 0.14 | 0.28 | 0.02* | 0.03* | 0.02* | 0.03* |
| 41 | 0.16 | 0.46 | 0.18 | 0.03* | 0.05* | 0.23 | 0.08 |
*Valores con diferencias significativas (Kruskal-Wallis P ≤ 0.05) en el aporte de nutrientes entre especies.
Los nutrientes encontrados en esta vía fueron menores que los depositados vía Pi y Pd. De acuerdo con el Cuadro 6, el macronutriente y micronutriente con mayor aporte medio anual fueron el Mg y Fe con 0.52 kg·ha-1·año-1 y 43.08 g·ha-1·año-1, respectivamente. De acuerdo con los resultados, C. greggii presentó los valores más altos de Mg, Ca Fe, Zn, Cu y Mn, y C. hookeri aportó K en mayor cantidad.
Cuadro 6 Aporte de macronutrientes y micronutrientes vía escurrimiento fustal por tipo de dosel vegetal.
| Dosel vegetal | Macronutrientes (kg·ha-1·año-1) | Micronutrientes (g·ha-1·año-1) | |||||
|---|---|---|---|---|---|---|---|
| Ca | K | Mg | Fe | Mn | Zn | Cu | |
| Acacia farnesiana | 0.02 | 0.01 | 0.07 | 5.66 | 0.52 | 0.47 | 0.18 |
| Condalia hookeri | 0.07 | 0.06 | 0.19 | 10.29 | 0.72 | 1.38 | 0.49 |
| Casimiroa greggii | 0.09 | 0.04 | 0.26 | 27.13 | 2.27 | 3.34 | 3.10 |
| Total | 0.18 | 0.11 | 0.52 | 43.08 | 3.51 | 5.19 | 3.77 |
Los estudios que estiman los flujos de nutrientes no consideran rutinariamente el análisis del Ef porque es volumétricamente menor en comparación con la Pd por evento (Germer, Zimmermann, Neill, Krusche, & Elsenbeer, 2012; Zhang et al., 2016); sin embargo, en el presente estudio, la contribución anual acumulada de nutrientes vía Ef es considerable, representando 0.80 y 6.4 % de macro y micronutrientes, respectivamente, en relación con el aportado por la Pi. Una explicación para las bajas concentraciones de nutrientes en el Ef es que, durante la circulación del agua a través de los fustes, sucedió una absorción considerable por briofitas epífitas (musgos y líquenes). Estas crecen abundantemente en los tallos y en los troncos de los árboles, y contribuyen a la captación y liberación de nutrientes (Dawoe, Barnes, & Oppong, 2018; Liu, Fox, & Xu, 2003).
Aporte neto de nutrientes
Los valores netos para macronutrientes presentaron el orden Ca>K>Mg, donde la estructura arbórea de A. farnesiana aportó los contenidos máximos de Ca y K, mientras que el tipo de dosel del MET agregó mayor cantidad de Mg. Las contribuciones totales de micronutrientes mostraron la siguiente tendencia: Fe>Zn>Mn>Cu, donde los valores más altos de Zn y Mn se depositaron en la parcela correspondiente a MET, Fe en C. hookeri, y Cu en A. farnesiana (Cuadro 7).
Cuadro 7 Aporte neto de macro y micronutrientes por tipo de dosel vegetal.
| Tipo de cobertura | Macronutrientes (kg·ha-1·año-1) | Micronutrientes (g·ha-1·año-1) | |||||
|---|---|---|---|---|---|---|---|
| Ca | K | Mg | Fe | Mn | Zn | Cu | |
| Acacia farnesiana | 72.35 | 33.16 | 11.42 | 661.62 | 102.98 | 151.27 | 65.21 |
| Condalia hookeri | 57.47 | 28.57 | 9.09 | 623.72 | 56.22 | 132.08 | 58.8 |
| Casimiroa greggii | 66.44 | 28.68 | 9.67 | 916.66 | 75.40 | 164.31 | 48.2 |
| Leucaena leucocephala | 55.21 | 27.45 | 8.31 | 394.18 | 101.86 | 115.43 | 26.59 |
| Matorral espinoso tamaulipeco | 67.85 | 23.78 | 13.55 | 342.11 | 118.93 | 203.38 | 31.64 |
| Total | 319.31 | 141.63 | 52.05 | 2 938.29 | 455.38 | 766.47 | 230.44 |
Durante el periodo de evaluación, entre las concentraciones de Pi y las cinco coberturas analizadas (Pd y Ef), se logró incorporar al suelo un promedio total de 675.16, 260.96, 857.32 y 3458.15 g·ha-1·año-1 de Mn, Cu, Zn y Fe, respectivamente, y 217.88, 72.45 y 328.70 kg·ha-1·año-1 de K, Mg y Ca, respectivamente.
Los resultados obtenidos en la presente investigación presentan variaciones en los aportes de nutrientes entre los doseles, confirmando que muchos de los aspectos en el ciclo de los nutrientes son afectados directamente por la morfología de las especies y por la fenología (Cantú & González, 2001; Germer, Elsenbeer, & Moraes, 2006; Jaramillo, 2003; Lilienfein & Wilcke, 2004; Návar et al., 2009).
pH
La prueba de Kruskal-Wallis no evidenció diferencias estadísticas significativas (P > 0.05) del pH entre las vías (Pd y Ef) que circulan a través de las coberturas analizadas y la precipitación incidente en los eventos analizados.
Durante el periodo de evaluación se observaron variaciones entre el pH de la Pi y de las Pd de las especies estudiadas. De acuerdo con los resultados, es razonable suponer que la lluvia, al fluir por las coberturas arbóreas, se recarga de sales minerales depositadas en el follaje, lo que ocasiona variaciones del pH en Pd respecto de Pi. El cambio más significativo se presentó en la Pd del dosel de A. farnesiana, con un promedio general de 7.19, con respecto a la Pi con valor promedio de 6.88, mientras que las Pd del MET y de L. leucocephala mostraron menos alteraciones.
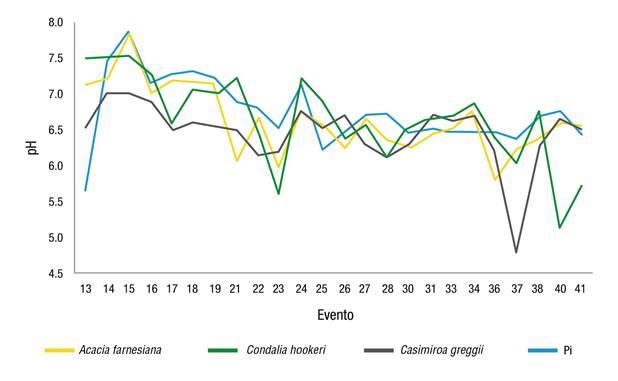
En la Figura 3 se observa que en el evento 13, la Pi presentó pH de 5.65 (ligeramente ácido), el cual fue menor con respecto a las determinadas en las Pd, donde el agua de lluvia, al fluir por el dosel de cada especie, sufrió un cambio significativo reduciendo su acidez a consecuencia del arrastre de cationes como el K y Mn. Por otra parte, en los eventos 32 y 35 ocurrió lo contrario, específicamente en la parcela de MET se observó que la Pd, al momento de fluir por el dosel de los árboles, presentaba acidificación con diferencia de 1.4 con relación al pH de Pi; cabe señalar que ambos eventos mostraron precipitaciones menores de 5 mm.

Figura 3 Variación del pH en la precipitación directa de cada dosel vegetal y en la precipitación incidente (Pi). MET: matorral espinoso tamaulipeco.
La tendencia de decremento del pH al pasar por el dosel vegetal también ha sido documentada en otros ecosistemas y se ha atribuido principalmente a la presencia de ácidos orgánicos y otras partículas volátiles en las estructuras arbóreas, así como al volumen de la Pd y Ef, lo cual pudo haber ocurrido en la parcela de MET en los eventos 32 y 35. Por ejemplo, Jiménez et al. (2006) mostraron que el pH de la Pi presentó un valor de 5.58 con ligero aumento a su paso por las copas de los árboles, alcanzando valores de 5.83 en Vochysia ferruginea Mart., 5.98 en Vochysia guatemalensis Standl. y 5.96 en un bosque secundario. Similarmente, Abbasian, Attarod, Sadeghi, Van Stan, y Hojjati (2015) evaluaron el aporte de nutrientes en la Pd de una plantación de Picea abies (L.) Karst y un bosque de Fagus orientalis Lipsky en el norte de Irán, y reportaron valores de pH entre 7 y 7.5 para ambas especies. Estos valores son similares a los resultados de A. farnesiana y C. hookeri considerados ligeramente alcalinos, mientras que C. greggii, el MET y L. leucocephala presentaron una acidez ligera. En el noreste de México, Cantú y González (2001) observaron que los valores de pH de la Pd en los doseles de pino (6.2), encino (6.3) y pino-encino (6.3) fueron más ácidos que la precipitación incidente (6.6).
La vía de Ef registró un total de 24 eventos, la cual se midió solo en A. farnesiana, C. hookeri y C. greggii, mostrando valores medios de pH de 6.63, 6.64 y 6.45, respectivamente; presentando ligero aumento en la acidez en comparación con Pi (6.74) (Figura 4). Este resultado coincide con el de Yáñez et al. (2014), quienes evaluaron las propiedades hidrológicas de un fragmento de MET e indicaron que el pH para Pi (7.4) disminuyó al circular por esta vía de redistribución de la precipitación (Ef = 6.9), lo cual pudo haber sido provocado por la presencia y arrastre de ácidos orgánicos y otras partículas presentes en el componente fustal.
Conductividad eléctrica
La prueba de Kruskal-Wallis no evidenció diferencias estadísticas significativas (P > 0.05) en la CE entre las vías que circulan a través de las coberturas analizadas (Pd y Ef) y la Pi en los eventos analizados.
Los resultados indican incremento de la CE al paso del agua por los doseles en relación con la observada en Pi (59.93 µS·cm-1), a causa del depósito de sólidos y sales minerales. La Pd con el valor medio más alto de CE en los 41 eventos evaluados corresponde a C. hookeri con 191.36 µS·cm-1, seguido del dosel de MET, A. farnesiana, L. leucocephala y C. greggii con 166.73, 155.13, 129.10 y 125.34 µS·cm-1, respectivamente (Figura 5).
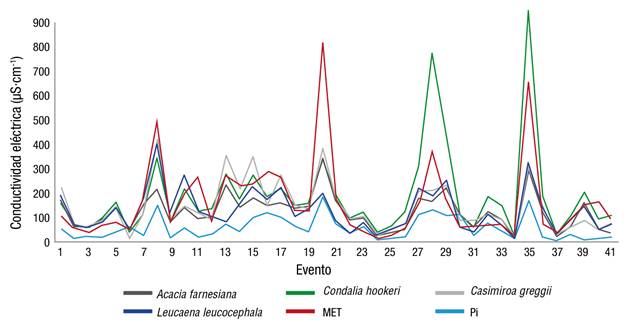
Figura 5 Comportamiento de la conductividad eléctrica en la precipitación directa para cada dosel vegetal y en la precipitación incidente (Pi). MET: matorral espinoso tamaulipeco.
El análisis de la CE en el Ef mostró un comportamiento similar a Pd, registrando valores de 56.49 µS·cm-1 en Pi, mientras que A. farnesiana presentó los registros más elevados con 219.16 µS·cm-1, seguida de C. hookeri y C. greggii con 155.57 y 92.01 µS·cm-1, respectivamente (Figura 6). Los resultados son similares a los encontrados por Yáñez et al. (2014) en la evaluación de MET, donde registraron valores de 71.7 µS·cm-1 para Pi, 145 µS·cm-1 para Pd y 126.8 µS·cm-1 para Ef.
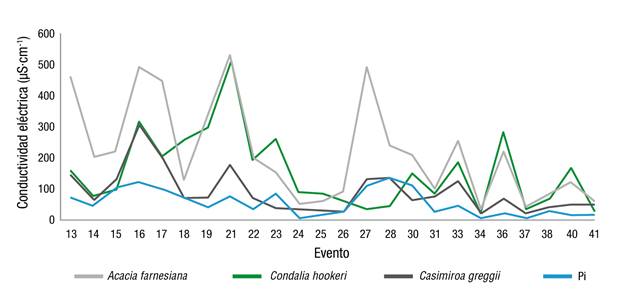
Figura 6 Comportamiento de la conductividad eléctrica en el escurrimiento fustal en tres especies y en la precipitación incidente (Pi).
El arrastre de aniones corticales y la poca cesión de cationes son responsables del aumento en la conductividad eléctrica. En general, Pérez, Cetina, Aldrete, Fenn, y Landois (2006) mencionan que el incremento de la CE de Ef y Pd, respecto a Pi, puede justificarse por el efecto de concentración, ya que al disminuir la cantidad y ocurrencia de eventos de lluvia hay acumulación mayor de iones en la atmósfera, los cuales son lavados por la lluvia posterior.
Conclusiones
La acumulación anual permitió detectar variaciones en los aportes entre precipitación incidente (Pi), precipitación directa (Pd) y escurrimiento fustal (Ef), manifestando claramente que la vía de Pd es la que deposita nutrientes al suelo en mayor medida. En general, no se encontraron diferencias estadísticas del aporte anual total de micro y macronutrientes entre las cinco coberturas en 41 eventos de lluvia. Los beneficios que ofrecen la precipitación y la vegetación de matorral en el ciclaje y enriquecimiento de nutrientes al suelo han quedado establecidos, permitiendo proponer estrategias de conservación, restauración y aprovechamientos para este tipo de ecosistema.
Agradecimientos
Los autores desean expresar su reconocimiento a la Facultad de Ciencias Forestales de la UANL y al CONACYT por el financiamiento de esta investigación, a través del proyecto Fondo Ciencia Básica SEP-CONACYT, 250732.
REFERENCIAS
Abbasian, P., Attarod, P., Sadeghi, S. M. M., Van Stan, J. T., & Hojjati, S. M. (2015). Throughfall nutrients in a degraded indigenous Fagus orientalis forest and a Picea abies plantation in the North of Iran. Forest Systems, 24(3), 2171-9845. doi: 10.5424/fs/2015243-06764 [ Links ]
André, F., Jonard, M., & Ponette, Q. (2008). Effects of biological and meteorological factors on stemflow chemistry within a temperate mixed Oak-beech stand. Science of the Total Environment, 393(1), 72-83. doi: 10.1016/j.scitotenv.2007.12.002 [ Links ]
Béjar, P. S., Cantú, S. I., Domínguez, G. T., González, R. H., Marmolejo, M. J., Yáñez, D. M., & Luna, R. E. (2018). Redistribución de la precipitación y aporte de nutrimentos en Pinus cooperi C.E. Blanco. Revista Mexicana de Ciencias Forestales, 9(50), 94-120. doi: 10.29298/rmcf.v9i50.237 [ Links ]
Cantú, S. I., & González, R. H. (2001). Interception loss, throughfall and stemflow chemistry in pine and oak forests in northeastern Mexico. Tree Physiology, 21(12-13), 1009-1013. Retrieved from https://www.researchgate.net/publication/11847169 [ Links ]
Dawoe, E. K., Barnes, V. R., & Oppong, S. K. (2018). Spatio-temporal dynamics of gross rainfall partitioning and nutrient fluxes in shaded-cocoa (Theobroma cocoa) systems in a tropical semi-deciduous forest. Agroforestry Systems, 92(2), 397-413. doi: 10.1007/s10457-017-0108-3 [ Links ]
Duarte, A. F., Gioda, A., Ziolli, R., & Duó, D. (2013). Contaminación atmosférica y deposición húmeda en la Amazonia brasileña. Revista Cubana de Salud Pública, 39(4), 627-639. Retrieved from http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S0864-34662013000400002 [ Links ]
Germer, S., Elsenbeer, H., & Moraes, J. M. (2006). Throughfall and temporal trends of rainfall redistribution in an open tropical rainforest, south-western Amazonia (Rondônia, Brazil). Hydrology and Earth System Sciences Discussions, 10(3), 383-393. doi: 10.5194/hess-10-383-2006. [ Links ]
Germer, S., Zimmermann, A., Neill, C., Krusche, A. V., & Elsenbeer, H. (2012). Disproportionate single-species contribution to canopy-soil nutrient flux in an Amazonian rainforest. Forest Ecology and Management, 267, 40-49. doi: 10.1016/j.foreco.2011.11.041 [ Links ]
González, R. H., Ramírez, L. R. G., Cantú, S. I., Gómez, M. M. V., & Uvalle, S.J. I. (2010). Composición y estructura de la vegetación en tres sitios del estado de Nuevo León, México. Polibotánica , ( 29) 91-106. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S1405-27682010000100004 [ Links ]
International Business Machines (IBM). (2013). IBM SPSS Statistics for Windows, version 22.0. Armonk, NY, USA: IBM Corp. [ Links ]
Jaramillo, R. A. (2003). La lluvia y el transporte de nutrimentos dentro de ecosistemas de bosque y cafetales. Cenicafé, 54(2), 134-144. Retrieved from http://hdl.handle.net/10778/249 [ Links ]
Jiménez, R. C., Calvo, A. J. C., & Arias, A. D. (2006). Lavado de nutrientes en plantaciones forestales de Vochysia ferruginea Mart. y Vochysia guatemalensis Donn. Sm., Sarapiquí, Costa Rica. Revista Forestal Mesoamericana Kurú, 3(8), 16-26. Retrieved from http://revistas.tec.ac.cr/index.php/kuru/article/view/512/439 [ Links ]
Levia, D. F., & Herwitz, S. R. (2005). Interspecific variation of bark water storage capacity of three deciduous tree species in relation to stemflow yield and solute flux to forest soils. CATENA, 64(1), 117-137. doi: 10.1016/j.catena.2005.08.001 [ Links ]
Lida, S. I., Tanaka, T., & Sugita, M. (2005). Change of interception process due to the succession from Japanese red pine to evergreen oak. Journal of Hydrology, 315, 154-166. doi: 10.1016/j.jhydrol.2005.03.024 [ Links ]
Lilienfein, J., & Wilcke, W. (2004). Water and element input into native, agri-and silvicultural ecosystems of the Brazilian savanna.Biogeochemistry,67(2), 183-212. doi: 10.1023/B:BIOG.0000015279.48813.9d [ Links ]
Liu, W., Fox, J. E., & Xu, Z. (2003). Nutrient budget of a montane evergreen broad‐leaved forest at Ailao Mountain National Nature Reserve, Yunnan, southwest China.Hydrological Processes , 17(6), 1119-1134. doi: 10.1002/hyp.1184 [ Links ]
Návar, J., González, J. M., & González, H. (2009). Gross precipitation and throughfall chemistry in legume species planted in Northeastern México.Plant and Soil , 318(1-2), 15-26. doi: 10.1007/s11104-008-9812-0 [ Links ]
Pérez, S. M., Cetina, A. V. M., Aldrete, A., Fenn, M. E., & Landois, L. L. (2006). Química de la precipitación pluvial en dos bosques de la cuenca de la Ciudad de México.Agrociencia , 40(2), 239-248. Retrieved from http://www.redalyc.org/pdf/302/30240209.pdf [ Links ]
Segovia, Z. J. A., Delgadillo, H. F., Lares, R. M. L., Huerta, D. M. A., Muñoz, B. A., & Torres, D. E. V. (2009). Aporte atmosférico y concentración de hierro disuelto en la capa superficial del Golfo de California. Ciencias Marinas, 35(1), 75-90. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S0185-38802009000100006 [ Links ]
Shen, W., Ren, H., Jenerette, G. D., Hui, D., & Ren, H. (2013). Atmospheric deposition and canopy exchange of anions and cations in two plantation forests under acid rain influence. Atmospheric Environment, 64, 242-250. doi: 10.1016/j.atmosenv.2012.10.015 [ Links ]
Staelens, J., De-Schrijver, A., Verheyen, K., & Verhoest, N. E. (2008). Rainfall partitioning into throughfall, stemflow, and interception within a single beech (Fagus sylvaticaL.) canopy: Influence of foliation, rain event characteristics, and meteorology. Hydrological Processes, 22(1), 33-45. doi: 10.1002/hyp.6610 [ Links ]
Taghi, A. M., Attarod, P., & Bayramzadeh, V. (2013). The role of rainfall size in canopy interception loss: An observational study in a typical beech forest. Middle East Journal of Scientific Research, 13(7), 876-882. doi: 10.5829/idosi.mejsr.2013.13.7.2721 [ Links ]
Yáñez, D. M. I., Cantú, S. I., González, R. H., & Uvalle, S. J. I. (2014). Redistribución de la precipitación en tres especies arbustivas nativas y una plantación de eucalipto del noreste de México. Tecnología y Ciencias del Agua, 5(2), 71-84. Retrieved from http://www.scielo.org.mx/scielo.php?script=sci_arttext&pid=S2007-24222014000200005&lng=es&nrm=iso&tlng=es [ Links ]
Zhang, Y. F., Wang, X. P., Pan, Y. X., & Hu, R. (2016). Variations of nutrients in gross rainfall, stemflow, and throughfall within revegetated desert ecosystems. Water, Air, & Soil Pollution, 227, 183. doi: 10.1007/s11270-016-2878-z [ Links ]
Recibido: 24 de Diciembre de 2018; Aprobado: 11 de Marzo de 2019
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
