










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.25 no.2 Chapingo may./ago. 2019 Epub 19-Feb-2021
https://doi.org/10.5154/r.rchscfa.2018.10.076
Artículo científico
Modelado espacial histórico y actual del oyamel (Abies religiosa [Kunth] Schltdl. & Cham.) en la Faja Volcánica Transmexicana
1Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales. Carretera Nacional kilómetro 145. C. P. 67700. Linares, Nuevo León, México.
Introducción:
Las condiciones climáticas en la Faja Volcánica Transmexicana durante el Holoceno medio fueron diferentes de las actuales, lo que pudo tener un efecto en las áreas de distribución histórica del oyamel (Abies religiosa [Kunth] Schltdl. & Cham.).
Objetivo:
Determinar si los requerimientos ambientales que delimitan la distribución actual del oyamel en la Faja Volcánica Transmexicana han cambiado desde hace 6 000 años.
Materiales y métodos:
Los registros de A. religiosa se obtuvieron de la plataforma Niche Toolbox. Se descargaron las variables WorldClim versión 2.0 para los periodos actual (1970-2000) y Holoceno medio (modelos CNRMCM5 y MIROC_ESM). Los modelos de distribución se generaron en MaxEnt utilizando 75 % de los datos para entrenamiento y 25 % para validación. Las variables más importantes de cada periodo se determinaron con la prueba de Jackknife.
Resultados y discusión:
Las superficies estimadas de oyamel fueron similares en ambos periodos. Aproximadamente, 86.5 % de la distribución del oyamel se encuentra en áreas naturales protegidas de la zona. Las variables ambientales limitantes de la distribución son la altitud, precipitación anual, precipitación de verano, temperatura media anual y oscilación diurna en temperatura. El Nevado de Toluca y la Ciudad de México presentaron superficies mayores de oyamel (+2 658.4 ha y +20 129.7 ha, respectivamente) durante el Holoceno medio. La precipitación anual era 80 a 224 mm mayor que la actual y la temperatura era 1 °C más fría.
Conclusión:
Las variables ambientales de mayor importancia para la distribución del oyamel son similares a las reportadas en la literatura, lo que indica similitud entre el nicho ecológico actual e histórico de A. religiosa.
Palabras clave: Nicho ecológico; variables ambientales; abeto; Holoceno medio; distribución potencial
Introduction:
Climatic conditions in the Trans-Mexican Volcanic Belt during the middle Holocene were different from today's conditions, which may have an effect on the historical distribution areas of the sacred fir (Abies religiosa [Kunth] Schltdl. & Cham.).
Objective:
To determine whether the environmental requirements that delimit the current distribution of the sacred fir in the Trans-Mexican Volcanic Belt have changed since 6 000 years ago.
Materials and methods:
A. religiosa records were obtained from the Niche Toolbox platform. The WorldClim version 2.0 variables for the current (1970-2000) and middle Holocene (CNRMCM5 and MIROC_ESM models) periods were downloaded. The distribution models were generated in MaxEnt using 75 % of the data for training and 25 % for validation. The most important variables of each period were determined with the Jackknife test.
Results and discussion:
The estimated sacred fir areas were similar in both periods. Approximately 86.5 % of the sacred fir distribution is found in protected natural areas of the zone. The limiting environmental variables of its distribution are: elevation, annual precipitation, summer precipitation, annual mean temperature and diurnal temperature oscillation. Nevado de Toluca and Mexico City had larger sacred fir areas (+2 658.4 ha and +20 129.7 ha, respectively) during the Middle Holocene. Annual precipitation was 80 to 224 mm higher than the current level and the temperature was 1 °C colder.
Conclusion:
The most important environmental variables for sacred fir distribution are similar to those reported in the literature, indicating similarity between the current and historical ecological niche of A. religiosa.
Keywords: Ecological niche; environmental variables; fir; Middle Holocene; potential distribution
Introducción
En la actualidad, los modelos de distribución de especies se han convertido en una herramienta útil para el manejo y conservación de la biodiversidad que, al integrarse con los sistemas de información geográfica, permiten construir modelos de distribución potencial. Con estos modelos es posible delimitar, estimar y predecir si la distribución de las especies cambia o se mantiene en un espacio geográfico ambiental (Manzanilla et al., 2019; Martínez-Méndez, Aguirre-Planter, Eguiarte, & Jaramillo-Correa, 2016; Monterrubio-Rico et al., 2016). Además, dichos modelos permiten anticipar posibles efectos del cambio climático en la distribución de las especies (Gutiérrez & Trejo, 2014; Peterson, 2011a; Sáenz-Romero, Rehfeldt, Ortega-Rodríguez, Marín-Togo, & Madrigal-Sánchez, 2015).
Los cambios del clima durante el pasado modificaron la distribución y abundancia de las especies de plantas (Caballero, Lozano-García, Vázquez-Selem, & Ortega, 2010). A partir del último máximo glacial (hace 21 000 años), el clima del planeta ha variado de periodos muy fríos (glaciales -8 °C) a periodos cálidos (interglaciales), donde la temperatura fue de 2 a 3 °C más cálida que la actual (Svensson et al., 2008). Se estima que las condiciones climáticas en la Faja Volcánica Transmexicana durante el Holoceno medio (hace 6 000 años) fueron 2 °C más cálidas que las actuales (Ferrusquía-Villafranca, 1998; Svensson et al., 2008). Estos cambios en el clima modificaron la composición y estructura de la vegetación de las zonas templadas y frías (Lozano-García & Vázquez-Selem, 2005), incluyendo la distribución de los bosques de coníferas (Caballero et al., 2010).
El modelado de la distribución potencial se ha usado para delimitar y predecir la distribución actual y futura de especies de coníferas (Pinaceae) en México (Manzanilla et al., 2019; Martínez-Méndez et al., 2016; Moreno-Letelier, Ortíz-Medrano, & Pinero, 2013; Sáenz-Romero et al., 2015; Sáenz-Romero, Rehfeldt, Duval, & Lindig-Cisneros, 2012); sin embargo, ha sido poco usado para reconstruir distribuciones históricas de especies de esta familia (Moreno-Letelier et al., 2013), incluyendo al género Abies Mill. De acuerdo con los datos del registro fósil, el género Abies se originó entre el periodo Cretácico tardío y el Eoceno temprano (Xiang, Wei, Shao, Wang, & Zhang, 2015). En la actualidad, existen especies del género con afinidad boreal y templada, distribuidas en Norteamérica y Eurasia, aunque algunos taxones se encuentran en las zonas montañosas y húmedas de Mesoamérica (Farjon & Filer, 2013; Madrigal, 1967; Rzedowski, 2006).
En México, el género Abies está representado por 10 especies (Martínez-Méndez et al., 2016) delimitadas a partir de caracteres morfológicos y con una distribución disyunta y restringida hacia las partes altas y húmedas de las principales cadenas montañosas (Madrigal, 1967; Martínez-Méndez et al., 2016; Rzedowski, 2006). Los bosques de oyamel ocupan alrededor de 144 000 ha (0.5 % de la superficie nacional) y constituyen el cuarto recurso forestal maderable de México (Organización de las Naciones Unidas para la Alimentación y la Agricultura [FAO], 2010; Secretaría del Medio Ambiente y Recursos Naturales [SEMARNAT], 2007). De las 10 especies reportadas, Abies religiosa (Kunth) Schltdl. & Cham. es el abeto u oyamel con mayor distribución (Martínez-Méndez et al., 2016). Las poblaciones de esta especie son pequeñas y aisladas, ubicadas principalmente en las zonas altas y húmedas del centro de México. No obstante, se piensa que, durante el Holoceno medio, dichos bosques ocuparon una superficie mayor que la actual. Bajo este trasfondo se planteó la siguiente pregunta de investigación: ¿Los requerimientos ambientales que delimitan la distribución actual del oyamel en la Faja Volcánica Transmexicana han cambiado en 6 000 años? Para responder esta cuestión se formularon los siguientes objetivos particulares: 1) delimitar y comparar la distribución histórica y actual del bosque de A. religiosa en la Faja Volcánica Transmexicana mediante el modelado de distribución potencial, 2) determinar y comparar las variables ambientales que delimitan el nicho ecológico de la especie en ambos periodos y 3) estimar la superficie boscosa de oyamel histórica y actual en áreas naturales protegidas del área de estudio.
Con base en los cambios climáticos ocurridos entre los periodos actual e histórico (Holoceno medio), y de acuerdo con la teoría del conservadurismo del nicho ecológico (Peterson, 2011b), se podría predecir que la distribución potencial de A. religiosa en la Faja Volcánica Transmexicana es similar para ambos periodos.
Materiales y métodos
Área de estudio
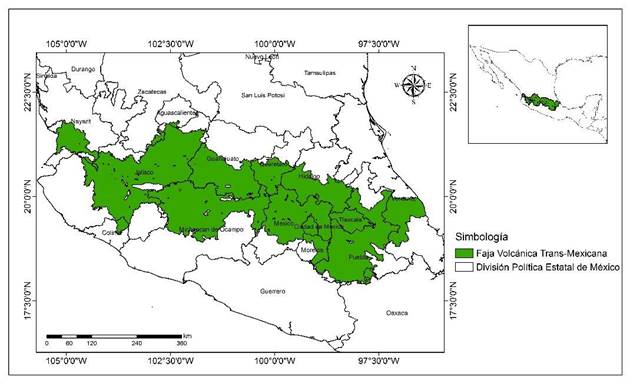
La Faja Volcánica Transmexicana se localiza en las coordenadas 17° 30’ y 20° 25’ LN, y 96° 20’ y 105° 20’ LO (Ferrusquía-Villafranca, 1998) (Figura 1); presenta altitudes de 256 a 5 650 m (Instituto Nacional de Estadística y Geografía [INEGI], 2018). La topografía de la zona presenta relieve accidentado y con mucha pendiente en las montañas (Ferrusquía-Villafranca, 1998). El clima dominante corresponde al tipo templado (húmedo y subhúmedo) con temperatura media anual de 12 a 18 °C y precipitación anual de 600 a 1 500 mm (García, 1998).
Descripción de la especie
Abies religiosa es un árbol monoico, perennifolio, con altura de hasta 60 m y diámetro máximo de 1.80 m. El árbol tiene corteza grisácea, áspera y agrietada con grosor de 18 a 25 mm; ramas en forma de cruz; hojas simples y alternas de 15 a 35 mm de longitud por 1.5 mm de ancho; y conos solitarios de 8 a 16 cm de largo y 4 a 6 cm de ancho (Protectora de Bosques [PROBOSQUE], 2007; Rzedowski, 2006).
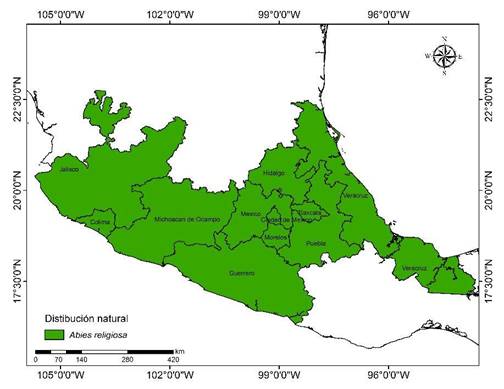
En México, la especie se localiza en Morelos, Estado de México, Hidalgo, Puebla, Michoacán, Jalisco, Colima, Guerrero, Tlaxcala, Veracruz y Ciudad de México (Martínez-Méndez et al., 2016) (Figura 2). En la Faja Volcánica Transmexicana, A. religiosa se distribuye en un gradiente altitudinal de 2 400 a 3 500 m y es la especie dominante del tipo de vegetación conocida en México como bosque de oyamel (Madrigal, 1967; Rzedowski, 2006).
Registros de Abies religiosa
Se descargaron 1 042 registros de A. religiosa de la plataforma Niche Toolbox (Osorio-Olvera, Vijay, Narayani, Soberón, & Falconi, 2017). Con el objetivo de evitar la autocorrelación espacial y el sobreajuste de los modelos (Monterrubio-Rico et al., 2016; Peterson & Nakazawa, 2008), se suprimieron las coordenadas duplicadas, mal georreferenciadas y cercanas entre sí (distancia >1 km), dejando un registro por cada celda de 1 km2. Al final del proceso de depuración, se obtuvieron 341 registros de A. religiosa distribuidos en el área de estudio, con los cuales se generaron los modelos de distribución potencial para los periodos actual e histórico.
Información bioclimática actual e histórica
En el Cuadro 1 se muestran las 19 variables bioclimáticas disponibles en WorldClim versión 2.0 para el periodo actual (1970-2000) (Fick & Hijmans, 2017) y de los modelos de circulación global (MCG) CNRMCM5 (Centro Nacional de Investigaciones Meteorológicas de Francia) y MIROC_ESM (Instituto de Investigaciones Oceánicas y Atmosféricas, Instituto Nacional de Estudios Ambientales y Agencia Japonesa para la Ciencia y Tecnología Marina y Terrestre) para el Holoceno medio. Estos últimos fueron generados a partir de Modelos Regionales del Proyecto CMIP5 (Coupled Model Intercomparison Project Phase 5, 2013) del Panel Intergubernamental de Cambio Climático (IPCC). Todas las variables tuvieron resolución espacial de 1 km2 y presentaron formato Geotiff (TIF). Además, se incluyeron las variables tipo de suelo (SUE; escala 1: 250 000) y altitud (DEM; modelo digital de elevación, resolución de 90 m), las cuales se descargaron en formato vectorial y ráster desde la plataforma del INEGI (INEGI, 2014, 2018). Al final, todas las variables se homogeneizaron a formato ASCII con escala de 1 km2.
Cuadro 1 Variables bioclimáticas utilizadas en la generación de los modelos actuales e históricos de distribución potencial de Abies religiosa en la Faja Volcánica Transmexicana.
| Variables bioclimáticas | Clave |
|---|---|
| Temperatura promedio anual (°C) | BIO1 |
| Oscilación diurna de la temperatura (°C) | BIO2 |
| Isotermalidad [(BIO2/BIO7)*100] | BIO3 |
| Estacionalidad de la temperatura (coeficiente de variación, %) | BIO4 |
| Temperatura máxima promedio del periodo más cálido (°C) | BIO5 |
| Temperatura mínima promedio del periodo más frío (°C) | BIO6 |
| Oscilación anual de la temperatura (°C) | BIO7 |
| Temperatura promedio del trimestre más lluvioso (°C) | BIO8 |
| Temperatura promedio del trimestre más seco (°C) | BIO9 |
| Temperatura promedio del trimestre más cálido (°C) | BIO10 |
| Temperatura promedio del trimestre más frío (°C) | BIO11 |
| Precipitación acumulada anual (mm) | BIO12 |
| Precipitación del periodo más lluvioso (mm) | BIO13 |
| Precipitación del periodo más seco (mm) | BIO14 |
| Estacionalidad de la precipitación (coeficiente de variación, %) | BIO15 |
| Precipitación del trimestre más lluvioso (mm) | BIO16 |
| Precipitación del trimestre más seco (mm) | BIO17 |
| Precipitación del trimestre más cálido (mm) | BIO18 |
| Precipitación del trimestre más frío (mm) | BIO19 |
Selección de variables
La autocorrelación espacial se eliminó mediante un análisis de multicolinearidad (Monterrubio-Rico et al., 2016; Peterson & Nakazawa, 2008), donde se descartaron aquellas variables con coeficiente de correlación ?#8805;0.85 (Manzanilla et al., 2019; Monterrubio-Rico et al., 2016), para maximizar la contribución de las variables en los modelos de distribución (Martínez-Méndez et al., 2016; Monterrubio-Rico et al., 2016; Peterson & Nakazawa, 2008).
Delimitación del área M
El área de modelamiento M es el espacio geográfico ambiental donde una especie se ha reportado o donde se supone que podría estar de acuerdo con el conocimiento biológico disponible (Martínez-Méndez et al., 2016; Soberón & Peterson, 2005). Para delimitar el área M se utilizaron las subprovincias fisiográficas donde se localizaban los registros de A. religiosa: Llanuras y Sierras de Querétaro e Hidalgo, Chapala, Mil Cumbres, Lagos y Volcanes de Anáhuac, Neovolcánica Tarasca y Volcanes de Colima (INEGI, 2001). El área M sirvió para recortar las variables a un mismo tamaño y con ello evitar la generación de áreas de distribución sobrestimadas (Monterrubio-Rico et al., 2016).
Generación de los modelos de distribución
Los modelos de distribución se generaron con los registros de presencia de A. religiosa en formato CSV y con las variables BIO1, BIO2, BIO3, BIO4, BIO12, BIO14, BIO15, BIO18, BIO19 en formato ASCII (Cuadro 1), tipo de suelo (SUE) y altitud (DEM).
Las variables fueron introducidas al programa de máxima entropía (MaxEnt), donde se utilizó 75 % de los registros de la especie para realizar la prueba de entrenamiento y el 25 % restante para la prueba de validación de los modelos (Martínez-Méndez et al., 2016; Monterrubio-Rico et al., 2016; Phillips, Anderson, & Schaphire, 2006). Se aplicó un replicado interno por validación cruzada y se desactivaron las casillas Extrapolate y Do clamping para evitar el sobreajuste de los modelos (Elith et al., 2011). Las salidas de los modelos fueron de tipo logística, la cual representa un índice ambiental de probabilidades con valores de 0 a 1. Los valores cercanos a 0 señalan condiciones ambientales no aptas, mientras que los valores cercanos a 1 sugieren condiciones ambientales excelentes para el crecimiento y desarrollo de la especie (Coitiño, Montenegro, Fallabrino, González, & Hernández, 2013; Phillips et al., 2006).
Se generaron y probaron cinco modelos de distribución con el fin de determinar por medio de las pruebas ROC (análisis de las características de funcionamiento del receptor del área bajo la curva [ABC]), Roc Parcial y Z, cuál modelo ajustaba con la distribución actual de la especie (Cuadro 2). Al modelo con mejor desempeño estadístico se sobrepuso el shapefile áreas naturales protegidas federales decretadas para la región (Comisión Nacional de Áreas Naturales Protegidas [CONANP], 2017). La superficie actual de oyamel dentro de la poligonal de las áreas naturales protegidas se estimó con el programa ArcGIS 10.3 (Environmental Systems Research Institute [ESRI], 2014).
Cuadro 2 Parámetros utilizados en la generación de los modelos potenciales de distribución de Abies religiosa en la Faja Volcánica Transmexicana.
| Modelo | Replicado interno | Regla de aplicación de umbral | Réplicas |
|---|---|---|---|
| M1 | Validación cruzada | Sensibilidad y especificidad de entrenamiento iguales | 1 000 |
| M2 | Validación cruzada | Máxima sensibilidad de entrenamiento más especificidad | 1 000 |
| M3 | Validación cruzada | Prueba de sensibilidad y especificidad iguales | 1 000 |
| M4 | Validación cruzada | Prueba de máxima sensibilidad más especificidad | 1 000 |
| M5 | Validación cruzada | Sin aplicación de regla de umbral | 500 |
Los parámetros del modelo de distribución actual con el mejor ajuste estadístico (Morrone & Escalante, 2016) se transfirieron al programa de máxima entropía (MaxEnt), para generar los modelos de distribución en el pasado.
Validación de los modelos
Los modelos se evaluaron por medio de los valores del ABC de ROC, donde los valores entre 0.7 a 0.9 se consideran buenos y los mayores de 0.9 excelentes (Coitiño et al., 2013; Peterson et al., 2011). No obstante, este tipo de validación ha sido cuestionado por no considerar ausencias verdaderas (Lobo, Jiménez, & Real, 2008; Peterson, Papes, & Soberón, 2008), por lo cual fue necesario efectuar un análisis Roc Parcial en el programa Tool for Partial Roc versión 1.0 (Narayani, 2008), para contrarrestar las deficiencias del ABC (Peterson et al., 2008).
Se siguieron las recomendaciones de Peterson et al. (2008) usando 1 000 réplicas por bootstrap entre los archivos de presencia e idoneidad de hábitat y estableciendo un error de omisión de 5 %. La prueba genera valores de 1 a 2, donde un valor con promedio de radio 1.0 equivale a un modelo al azar (Garza-López et al., 2016; Lobo et al., 2008; Peterson et al., 2008). Para determinar si los modelos eran válidos estadísticamente se hizo una prueba de Z entre las proporciones del ABC de Roc Parcial (Martínez-Méndez et al., 2016). El mejor modelo se eligió con base en el valor más alto de las pruebas de Roc Parcial, desviación estándar menor y un valor confiable de Z (P < 0.01).
Los valores de las salidas logísticas de los modelos se reclasificaron en tres categorías de calidad del hábitat con intervalos iguales (baja, moderada y alta) con la herramienta reclass del programa ArcMap 10.3 (ESRI, 2014). Los valores de la categoría de calidad de hábitat alta se emplearon para transformar los modelos continuos a binarios (apto-no apto). Con base en la reclasificación, se estimó la superficie de la distribución potencial de A. religiosa en la Faja Volcánica Transmexicana para ambos periodos analizados.
Las proyecciones de los modelos con el mejor desempeño estadístico para cada periodo se visualizaron y plasmaron en mapas de distribución potencial con el programa ArcMap 10.3 (ESRI, 2014). Por último, las variables más importantes de cada periodo analizado se determinaron con la prueba de Jackknife, la cual permitió cuantificar la contribución de las variables ambientales en los modelos de distribución (Phillips et al., 2006).
Resultados y discusión
Los resultados del ABC de la prueba ROC de los modelos actuales obtuvieron valores mayores de 0.9 (datos de entrenamiento y validación) para ambas pruebas, al igual que los modelos para el Holoceno medio. Estos valores indican que el desempeño de los modelos generados para ambos periodos fue excelente (>0.9; Coitiño et al., 2013; Monterrubio-Rico et al., 2016; Peterson et al., 2011); por lo tanto, se consideró que los modelos de distribución fueron confiables.
El desempeño de los cinco modelos fue similar para la prueba de ROC; sin embargo, al aplicar la prueba de Roc Parcial, el modelo con el mejor ajuste estadístico y menor desviación estándar fue el modelo 2 (Cuadro 3).
Cuadro 3 Resultados Roc Parcial y Z de los modelos de distribución potencial histórica y actual de Abies religiosa en la Faja Volcánica Transmexicana.
| Periodo analizado | Modelo | Promedio radios Roc Parcial | Desviación estándar | Prueba de Z |
|---|---|---|---|---|
| Presente | M1 | 1.494 | 0.129 | P < 0.01 |
| M2 | 1.500 | 0.123 | P < 0.01 | |
| M3 | 1.494 | 0.128 | P < 0.01 | |
| M4 | 1.492 | 0.130 | P < 0.01 | |
| M5 | 1.475 | 0.134 | P < 0.01 | |
| Histórico | CRNMCM5 | 1.551 | 0.114 | P < 0.01 |
| MIROC-ESM | 1.496 | 0.138 | P < 0.01 |
Los modelos indicaron que la distribución potencial actual (1970-2000) de A. religiosa en la Faja Volcánica Transmexicana es de 194 387.3 ha. La FAO (2010) y SEMARNAT (2007) reportan 144 000 ha, lo cual indica que, posterior al año 2000, hubo una reducción de 50 387.3 ha de la superficie boscosa de oyamel. Además, se encontró que 86.5 % (168 148 ha) de la distribución potencial actual del oyamel se encuentra dentro de áreas naturales protegidas de la Faja Volcánica Transmexicana. Este resultado concuerda con lo mencionado por Rzedowski (2006) y Martínez-Méndez et al. (2016), quienes señalan que A. religiosa es la única especie de abeto mexicano que presenta la mayor parte de su distribución en las zonas altas y húmedas dentro de áreas naturales de la Faja Volcánica Transmexicana, lo cual ha proporcionado (hasta cierto punto) una protección legal a la especie.
De acuerdo con los resultados de los modelos históricos, la superficie boscosa estimada de A. religiosa fue de 190 466.2 ha (CNRMCM5) a 193 563.2 ha (MIROC_ESM), lo que se traduce en 49 563.2 a 46 466.2 ha más que la superficie actual reportada por SEMANART (2007) y FAO (2010), respectivamente, pero 824.1 ha (MIROC_ESM) y 3 921.1 ha (CNRMCM5) menores a lo estimado en el presente estudio (Figuras 3 y 4). Con base en los resultados de las pruebas binomiales de ROC, Roc parcial y Z (P < 0.01) se seleccionó el modelo CRNMCM5, el cual presentó el mejor ajuste (Cuadro 3), donde los valores cercanos a 2 indicaron que el ajuste estadístico del modelo fue confiable según lo mencionado por Garza-López et al. (2016), Narayani (2008) y Peterson et al. (2008).
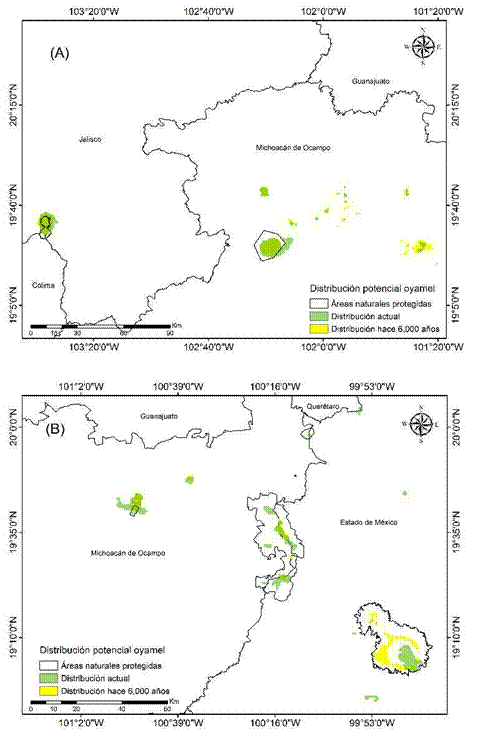
Figura 3 Distribución potencial actual (1970-2000) e histórica del oyamel (Abies religiosa) en la Faja Volcánica Transmexicana: zona Occidente (A) y Centro I (B).

Figura 4 Distribución potencial actual (1970-2000) e histórica del oyamel (Abies religiosa) en la Faja Volcánica Transmexicana: zona Centro II (C) y Oriente (D).
Distribución potencial histórica y actual
Dada la diversificación reciente del género Abies en el mundo (Xiang et al., 2015) es muy probable que las condiciones climáticas de hace 6 000 a 12 000 años fuesen más cálidas (+2 °C) en zonas templadas y frías de Norteamérica (Caballero et al., 2010; Svensson et al., 2008). Esto posiblemente delimitó la distribución de los bosques de coníferas hacia las zonas templadas y frías de la Faja Volcánica Transmexicana (Caballero et al., 2010; Madrigal, 1967; Martínez-Méndez et al., 2016; Rzedowski, 2006).
De acuerdo con la prueba de Jackknife, las variables ambientales limitantes con mayor importancia en la distribución de A. religiosa en la Faja Volcánica Transmexicana fueron altitud (DEM con 56.5 %), precipitación anual acumulada (BIO12 con 18.4 %) y precipitación del trimestre más cálido (BIO18 con 8.3 %). Estas tres variables explicaron 83.2 % de la variabilidad del modelo. Estos resultados son similares a los encontrados por Farjon y Filer (2013), Madrigal (1967) y Rzedowski (2006), quienes indican que la humedad anual constante y lluvias abundantes en verano son esenciales para la distribución de este abeto. En el presente estudio se observó que el gradiente altitudinal idóneo para el crecimiento y desarrollo de A. religiosa fue de 2 129 hasta 3 687 m. Estos resultados son similares a los que se reportan en la literatura, ya que trabajos previos mencionan que la distribución de A. religiosa oscila de 2 400 hasta 3 660 m (Calderón de Rzedowski & Rzedowski, 2001; Madrigal, 1967; Nieto de Pascual-Pola, 1995, Rzedowski, 2006). Cabe resaltar que, de acuerdo con los resultados de este estudio, la distribución del bosque de oyamel se puede encontrar 271 m por debajo de la altitud mínima y 27 m por encima de la altitud máxima reportada en la literatura. En cuanto a la precipitación anual acumulada, los modelos actuales estimaron un promedio de 1 125 mm. Este valor fue ligeramente mayor que el reportado por Madrigal (1967) y similar a lo mencionado por Rzedowski (2006) (1 000 mm y >1 000 mm, respectivamente). Martínez-Méndez et al. (2016) y Sáenz-Romero et al. (2012) mencionan que la precipitación de los meses de verano es determinante en la distribución de la especie, lo que concuerda con lo obtenido en el presente estudio; sin embargo, los estudios dendrocronológicos realizados para la especie, dentro de la zona de estudio, indican que la precipitación de invierno-primavera es de alta relevancia para el crecimiento de A. religiosa (Cerano-Paredes et al., 2014; Huante, Rincón, & Swetnam, 1991). Lo anterior difiere con lo obtenido en este estudio, pues los modelos evaluados no consideraron a la precipitación de invierno-primavera como un factor clave en la distribución del oyamel. No obstante, dicha variable podría ser de mayor relevancia que la precipitación en los meses de verano, durante la etapa de crecimiento radial de la especie.
Por otro lado, las variables más importantes en la predicción de distribución histórica (modelo MIROC_ESM) fueron temperatura media anual (BIO1 con 65.9 %), precipitación anual acumulada (BIO12 con 18.6 %) y oscilación diurna en temperatura (BIO2 con 4.6 %); estas tres variables contribuyeron con 89.1 % en la variación del modelo. Para el modelo CNRMCM5, las variables de mayor importancia fueron la temperatura media anual (BIO1 con 62.4 %), la precipitación anual acumulada (BIO12 con 16.2 %) y la oscilación diurna en temperatura (BIO2 con 6.5 %), las cuales contribuyeron con 85.1 % del modelo. Los resultados del presente estudio concuerdan con los de Madrigal (1967) y Rzedowski (2006), quienes mencionan que la oscilación diurna en la temperatura (11 a 16 °C) es determinante para el crecimiento de A. religiosa. Por lo tanto, se corroboró que esta variable ha sido un factor limitante desde hace 6 000 años (10.6 a 17.2 °C) en la distribución del oyamel en la Faja Volcánica Transmexicana. Con respecto a la temperatura media anual, los valores entre 4.6 y 15.7 °C son congruentes con lo reportado por Rzedowski (2006), quien menciona que fluctúan entre 7 y 15 °C. Por otra parte, según las condiciones climáticas estimadas por los modelos de circulación global, la precipitación anual durante el Holoceno medio fue de 80 a 224 mm mayor que la actual y la temperatura promedio anual fue 1 °C más fría y no más cálida como lo mencionan Svensson et al. (2008) y Caballero et al. (2010).
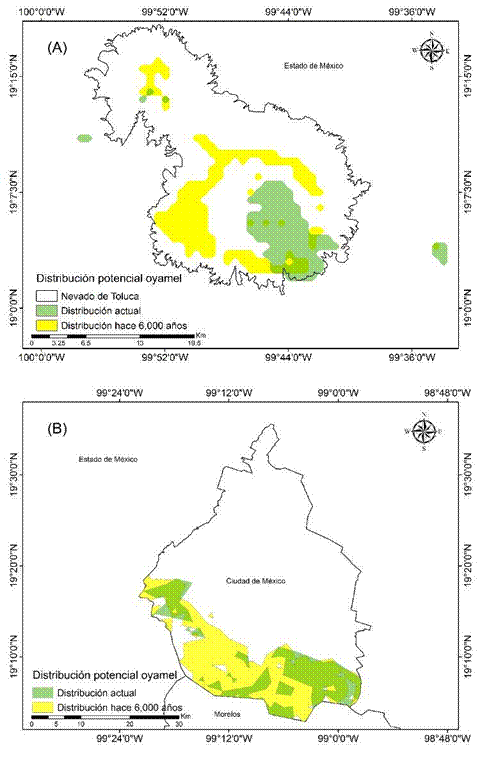
Debido a que las superficies estimadas y las condiciones ambientales descritas en la literatura y en este estudio presentaron similitudes, se puede suponer que la distribución potencial de A. religiosa ha permanecido estable a través del tiempo (según la teoría del conservadurismo de nicho ecológico; Peterson, 2011b). Así, en el presente estudio, las evidencias encontradas por los modelos actual e histórico parecen indicar que la distribución de la especie en la Faja Volcánica Transmexicana no ha cambiado en 6 000 años. Esto probaría (con la limitante de que no se cuentan con registros palinológicos) que el nicho ecológico de A. religiosa se ha mantenido durante los periodos evaluados. Cabe resaltar que, aunado a que la distribución actual e histórica no cambió significativamente, se logró identificar que sitios como el Nevado de Toluca y la Ciudad de México presentaron superficies mayores del oyamel en el pasado: +2 658.4 ha y +20 129.7 ha, respectivamente (Cuadro 4; Figura 5). Por lo tanto, se puede inferir que las condiciones climáticas de hace 6 000 años fueron más frías y húmedas en las zonas montañosas templadas y frías de la Faja Volcánica Transmexicana, lo que provocó que la distribución del oyamel se mantuviera ahí hasta el presente (Madrigal, 1967; Martínez-Méndez et al., 2016; Ramírez-Barahona & Eguiarte, 2013; Rzedowski, 2006).
Cuadro 4 Sitios con mayor distribución potencial histórica de bosque de Abies religiosa en la Faja Volcánica Transmexicana.
| Sitio | Distribución potencial actual (ha) | Distribución potencial pasada (ha) | Cambio (%) |
|---|---|---|---|
| Nevado de Toluca | 6 847.8 | 9 506.2 | -28 |
| Ciudad de México | 19 340.9 | 39 470.6 | -51 |
Conclusiones
El presente estudio representa una primera aproximación en la estimación de la distribución potencial de Abies religiosa hace 6 000 años en la Faja Volcánica Transmexicana. Estos resultados aportan información valiosa sobre las variables ambientales (altitud, precipitación anual, precipitación de verano, temperatura media anual y oscilación diurna en temperatura) que han limitado la distribución de esta especie. Hace 6 000 años, las condiciones climáticas correspondientes a la distribución del oyamel dentro de la Faja Volcánica Transmexicana fueron más frías y húmedas que las actuales. Aproximadamente, 86.5 % de la distribución de A. religiosa se encuentra dentro de áreas naturales protegidas de la zona; sin embargo, esta protección no la hace inmune a disturbios naturales y antropogénicos. Pese a que la distribución histórica y actual del oyamel en la Faja Volcánica Transmexicana fue similar, se detectó que sitios como el Nevado de Toluca y la Ciudad de México presentaron superficies más amplias en el pasado.
Agradecimientos
A la Universidad Autónoma de Nuevo León, Facultad de Ciencias Forestales. Al Consejo Nacional de Ciencia y Tecnología (CONACYT) por el apoyo de beca de posgrado al primer autor. A los revisores anónimos por sus comentarios y sugerencias, los cuales ayudaron a mejorar la calidad del presente manuscrito.
REFERENCIAS
Caballero, M., Lozano-García, S., Vázquez-Selem, L., & Ortega, B. (2010). Evidencias de cambio climático y ambiental en registros glaciales y en cuencas lacustres del centro de México durante el último máximo glacial. Boletín de la Sociedad Geológica Méxicana, 62(3), 359-377. Retrieved from http://www.redalyc.org/articulo.oa?id=94319371005 [ Links ]
Calderón de Rzedowski, G., & Rzedowski, J. (2001). Flora fanerogámica del Valle de México (2.ª ed.). Michoacán, México: Instituto de Ecología A. C. & Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. [ Links ]
Cerano-Paredes, J., Villanueva-Díaz, J., Cervantes-Martínez, R., Vázquez-Selem, L., Trucios-Caciano, R., & Guerra de la Cruz, V. (2014). Reconstrucción de precipitación invierno-primavera para el Parque Nacional Pico de Tancítaro, Michoacán. Boletín Investigaciones Geográficas, 83, 42-55. doi: 10.14350/rig.35190 [ Links ]
Coitiño, H. I., Montenegro, F., Fallabrino, A., González, E. M., & Hernández, D. (2013). Distribución actual y potencial de Cabassous tatouay y Tamandua tetradactyla en el límite sur de su distribución: implicancias para su conservación en Uruguay. Edentata, 14(1), 23-34. doi: 10.5537/020.014.0104 [ Links ]
Comisión Nacional de Áreas Naturales Protegidas (CONANP). (2017). Áreas Naturales Protegidas Federales de México. Archivo vectorial shapefile. Retrieved November 11, 2017 from http://www.conabio.gob.mx/informacion/metadata/gis/anpmay17gw.xml?_httpcache=yes&_xsl=/db/metadata/xsl/fgdc_html.xsl&_indent=no [ Links ]
Coupled Model Intercomparison Project Phase 5 (CMIP5). (2013). Retrieved January 15, 2017 from http://www.worldclim.org/paleo-climate1 [ Links ]
Elith, J., Phillips, S. J., Hastie, T., Dudík, M., Chee, Y. E., & Yates, C. J. (2011). A statistical explanation of MaxEnt for ecologists. Diversity and Distributions, 17(1), 43-57. doi: 10.1111/j.1472-4642.2010.00725.x [ Links ]
Environmental Systems Research Institute (ESRI). (2014). ArcGIS desktop, version 10.3. Redlands, California, Virginia, USA: Author. [ Links ]
Farjon, A., & Filer, D (2013). An atlas of the world´s conifers: An analysis of their distribution, biogeography, diversity, and conservation status. Netherlands: Brill Academic Publishers. [ Links ]
Ferrusquía-Villafranca, I. (1998). Geología de México: una sinopsis. In T. P. Ramamoorthy, R. Bye, A. Lot, & J. Fa. (Eds.), Diversidad biológica de México (pp. 3-108). México: Universidad Nacional Autónoma de México. [ Links ]
Fick, S. E., & Hijmans, R. J. (2017). WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology, 37(12), 4302-4317. doi: 10.1002/joc.5086 [ Links ]
García, E. (1998). Modificaciones al sistema de clasificación climática de Köppen (para adaptarlo a las condiciones de la República Mexicana). México: Instituto de Geografía, Universidad Nacional Autónoma de México. [ Links ]
Garza-López, M., Ortega-Rodríguez, J. M., Zamudio-Sánchez, F. J., López-Toledo, J. F., Domínguez-Álvarez, F. A., & Sáenz-Romero, C. (2016). Calakmul como refugio de Swietenia macrophylla King ante el cambio climático. Botanical Sciences, 94(1), 43-50. doi: 10.17129/botsci.500 [ Links ]
Gutiérrez, E., & Trejo, I. (2014). Efecto del cambio climático en la distribución potencial de cinco especies arbóreas de bosque templado en México. Revista Mexicana de Biodiversidad, 85(1), 179-188. doi: 10.7550/rmb.37737 [ Links ]
Huante, P., Rincón, E., & Swetnam, T. W. (1991). Dendrochronology of Abies religiosa in Michoacan, Mexico. Tree-Ring Bulletin, 51, 15-28. Retrieved from https://www.researchgate.net/publication/228116883_Dendrochronology_of_Abies_religiosa_in_Michoacan_Mexico [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI). (2001). Conjunto de datos vectoriales fisiográficos. Continuo Nacional. Escala 1:1 000 000. Serie I. Subprovincias Fisiográficas de México. Archivo vectorial shapefile. Retrieved November 11, 2017 from http://www.beta.inegi.org.mx/temas/mapas/fisiografia/ [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI). (2014). Conjunto de datos vectoriales edafológicos. Continuo Nacional. Escala 1: 250 000. Serie II. Archivo vectorial shapefile. Retrieved November 11, 2017 from http://www.conabio.gob.mx/informacion/metadata/gis/eda250s2gw.xml?_httpcache=yes&_xsl=/db/metadata/xsl/fgdc_html.xsl&_indent=no [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI). (2018). Continuo de Elevaciones Mexicano. Archivo ráster. Retrieved September 8, 2018 from http://www.beta.inegi.org.mx/app/geo2/elevacionesmex/index.jsp [ Links ]
Lobo, J. M., Jiménez, V. A., & Real, R. (2008). AUC: A misleading measure of the performance of predictive distribution models. Global Ecology and Biogeography, 17(2), 145-151. doi: 10.1111/j.1466-8238.2007.00358.x [ Links ]
Lozano-García, M. S., & Vázquez-Selem, L. (2005). A high-elevation Holocene pollen record from Iztaccihuatl volcano, central Mexico. The Holocene, 15(3), 329-338. doi: 10.1191/0959683605hl814rp [ Links ]
Madrigal, S. X. (1967). Contribución al conocimiento de la ecología de los bosques de oyamel (Abies religiosa (HBK) Schl. & Cham.) en el Valle de México. México: Instituto Nacional de Investigaciones Forestales. [ Links ]
Manzanilla, Q. U., Delgado, V. P., Hernández, R. J., Molina, S. A., García, M. J. J., & Rocha, G. M. del C. (2019). Similaridad del nicho ecológico de Pinus montezumae y P. pseudostrobus (Pinaceae) en México: implicaciones para la selección de áreas productoras de semillas y de conservación. Acta Botánica Mexicana, 126, e1398. doi: 10.21829/abm126.2019.1398 [ Links ]
Martínez-Méndez, N., Aguirre-Planter, E., Eguiarte, E. L., & Jaramillo-Correa, J. P. (2016). Modelado de nicho ecológico de las especies del género Abies (Pinaceae) en México: algunas implicaciones taxonómicas y para la conservación. Botanical Sciences, 94(1), 5-24. doi: 10.17129/botsci.508 [ Links ]
Monterrubio-Rico, T. C., Charre-Medellín, J. F., Pacheco-Figueroa, C., Arriaga-Weiss, S., Valdez-Leal, J. D., Cancino-Murillo, R., …Rubio-Rocha, A. (2016). Distribución potencial histórica y contemporánea de la familia Psittacidae en México. Revista Mexicana de Biodiversidad, 87(3), 1103-1117. doi: 10.1016/j.rmb.2016.06.004 [ Links ]
Moreno-Letelier, A., Ortíz-Medrano, A., & Pinero, D. (2013). Niche divergence versus neutral processes: Combined environmental and genetic analyses identify contrasting patterns of differentiation in recently diverged pine species. PLoS ONE, 8(10), e78228. doi: 10.1371/journal.pone.0078228 [ Links ]
Morrone, J. J., & Escalante, T. (2016). Introducción a la biogeografía (1.a ed.). México: Universidad Nacional Autónoma de México . [ Links ]
Narayani, B. (2008). Tool for Partial ROC version 1.0. Lawrence, KS, USA: University of Kansas-CONABIO. Retrieved January 7, 2018 from http://nicho.conabio.gob.mx/home/proposito-y-guia-del-usuario/validacion-del-modelo [ Links ]
Nieto de Pascual-Pola, M. C. (1995). Estudio sinecológico del bosque de oyamel de la cañada de Contreras, Distrito Federal. Revista Ciencia Forestal en México, 20, 3-34. [ Links ]
Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO). (2010). Evaluación de los recursos forestales mundiales 2010: Informe nacional, México. Roma, Italia: Author. Retrieved from https://www.researchgate.net/publication/263274117_Evaluacion_De_Los_Recursos_Forestales_Mundiales_2010_Informe_Nacional_Mexico [ Links ]
Osorio-Olvera, L., Vijay, B., Narayani, B., Soberón, J., & Falconi, M. (2017). Ntbox: From getting biodiversity data to evaluating species distributions models in a friendly GUI environment. R package version 0.2.5.4. Retrieved January 15, 2017 from https://github.com/luismurao/ntbox [ Links ]
Peterson, A. T. (2011a). Paleoclimates: Understanding climate change past and present. Quarterly Review of Biology, 86(4), 342-343. doi: 10.1086/662496 [ Links ]
Peterson, A. T. (2011b). Ecological niche conservatism: a time-structured review of evidence. Journal of Biogeography, 38(5), 817-827. doi: 10.1111/j.1365-2699,2010.02456.x [ Links ]
Peterson, A. T., & Nakazawa, Y. (2008). Environmental data sets matter in ecological niche modelling: an example with Solenopsis invicta and Solenopsis richteri. Global Ecology and Biogeography, 17(1), 135-144. doi: 10.1111/j.1466-8238.2007.00347.x [ Links ]
Peterson, A. T., Papes, M., & Soberón, J. (2008). Rethinking receiver operating characteristic analysis applications in ecological niche modeling. Ecological Modelling, 213(1), 63-72. doi: 10.1016/j.ecolmodel.2007.11.008 [ Links ]
Peterson, A. T., Soberón, J., Pearson, R. G., Anderson, R. P., Martinez-Meyer, E., Nakamura, M., & Araujo, M. B. (2011). Ecological niches and geographic distributions. USA: Princeton University Press. [ Links ]
Phillips, S. J., Anderson, R. P., & Schaphire, R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecological Modelling, 190(3-4), 231-259. doi: 10.1016/j.ecolmodel.2005.03.026 [ Links ]
Protectora de Bosques (PROBOSQUE). (2007). Manual de producción de planta forestal. Clima templado frío. Retrieved from http://www.earthgonomic.com/biblioteca/2007_SEDAGRO_Manual_de_Produccion_Forestal.pdf [ Links ]
Ramírez-Barahona, S., & Eguiarte, F. L. E. (2013). The role of glacial cycles in promoting genetic diversity in the Neotropics. The case of cloud forests during the Last Glacial Maximum. Ecology and Evolution, 3(3), 725-738. doi: 10.1002/ece3.483 [ Links ]
Rzedowski, J. (2006). Vegetación de México (1.a ed. digital). México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO). [ Links ]
Sáenz-Romero, C., Rehfeldt, G. E., Duval, P., & Lindig-Cisneros, R. A. (2012). Abies religiosa habitat prediction in climatic change scenarios and implications for monarch butterfly conservation in Mexico. Forest Ecology and Management, 275, 98-106. doi: 10.1016/j.foreco.2012.03.004 [ Links ]
Sáenz-Romero, C., Rehfeldt, G. E., Ortega-Rodríguez, J. M., Marín-Togo, M. C., & Madrigal-Sánchez, X. (2015). Pinus leiophylla suitable habitat for 1961-1990 and future climate. Botanical Sciences, 93(4), 709-718. doi: 10.17129/botsci.86 [ Links ]
Secretaría de Medio Ambiente y Recursos Naturales (SEMARNAT). (2007). Anuario estadístico de la producción forestal. México: Autor. Retrieved from https://www.gob.mx/cms/uploads/attachment/file/282952/2007.pdf [ Links ]
Soberón, J., & Peterson, A. T. (2005). Interpretation of models of fundamental ecological niches and species distribution areas. Biodiversity Information, 2, 1-10. doi: 10.17161/bi.v2i0.4 [ Links ]
Svensson, A., Andersen, K. K., Bigler, M., Clausen, H. B., Dahl-Jensen, D., Davies, S. M., …Vinter, B. M. (2008). A 60 000 year Greenland stratigraphic ice core chronology. Climate of the Past, 4, 47-57. doi: 10.5194/cp-4-47-2008 [ Links ]
Xiang, Q. P., Wei, R., Shao, Z. Y., Wang, X. Q., & Zhang, X. C. (2015). Phylogenetic relationships, possible ancient hybridization and biogeography history of Abies (Pinaceae) based on data from nuclear, plastid and mitochondrial genomes. Molecular Phylogenetics and Evolution, 82(A), 1-14. doi: 10.1016/j.ympev.2014.10.008 [ Links ]
Recibido: 30 de Octubre de 2018; Aprobado: 28 de Febrero de 2019
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
