











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Today, species distribution models have become a useful tool for biodiversity management and conservation that, when integrated with geographic information systems, enable the construction of potential distribution models. With these models it is possible to delimit, estimate, and predict whether the distribution of species changes or remains the same in an environmental geographic space (Manzanilla et al., 2019; Martínez-Méndez, Aguirre-Planter, Eguiarte, & Jaramillo-Correa, 2016; Monterrubio-Rico et al., 2016). In addition, these models allow us to anticipate possible effects of climate change on the distribution of species (Gutiérrez & Trejo, 2014; Peterson, 2011a; Sáenz-Romero, Rehfeldt, Ortega-Rodríguez, Marín-Togo, & Madrigal-Sánchez, 2015).
Climate changes during the past have modified the distribution and abundance of plant species (Caballero, Lozano-García, Vázquez-Selem, & Ortega, 2010). Since the last glacial maximum (21 000 years ago), the planet's climate has varied from very cold periods (glacial -8 °C) to warm periods (interglacial), where the temperature was 2 to 3 °C warmer than today (Svensson et al., 2008). It is estimated that the climatic conditions in the Trans-Mexican Volcanic Belt during the Middle Holocene (6,000 years ago) were 2 °C warmer than today (Ferrusquía-Villafranca, 1998; Svensson et al., 2008). These climate changes modified the composition and structure of vegetation in temperate and cold zones (Lozano-García & Vázquez-Selem, 2005), including the distribution of coniferous forests (Caballero et al., 2010).
Potential distribution modeling has been used to delimit and predict the current and future distribution of coniferous species (Pinaceae) in Mexico (Manzanilla et al., 2019; Martínez-Méndez et al., 2016; Moreno-Letelier, Ortíz-Medrano, & Pinero, 2013; Sáenz-Romero et al., 2015; Sáenz-Romero, Rehfeldt, Duval, & Lindig-Cisneros, 2012). However, it has been little used to reconstruct historical distributions of species of this family (Moreno-Letelier et al., 2013), including the genus Abies Mill. According to fossil record data, the genus Abies originated between the late Cretaceous period and the early Eocene (Xiang, Wei, Shao, Wang, & Zhang, 2015). At present, there are species of the genus with boreal and temperate affinity, distributed in North America and Eurasia, although some taxa are found in the mountainous and humid areas of Mesoamerica (Farjon & Filer, 2013; Madrigal, 1967; Rzedowski, 2006).
In Mexico, the genus Abies is represented by 10 species (Martínez-Méndez et al., 2016) delimited by morphological characters and with a disjunct and restricted distribution towards the high and humid parts of the main mountain ranges (Madrigal, 1967; Martínez-Méndez et al., 2016; Rzedowski, 2006). Sacred fir forests occupy about 144 000 ha (0.5 % of the national area) and constitute the fourth largest timber resource in Mexico (Organización de las Naciones Unidas para la Alimentación y la Agricultura [FAO], 2010; Secretaría del Medio Ambiente y Recursos Naturales [SEMARNAT], 2007). Of the 10 species reported, Abies religiosa (Kunth) Schltdl. & Cham. is the most widely distributed fir (Martínez-Méndez et al., 2016). The populations of this species are small and isolated, located mainly in the high and humid areas of central Mexico. Despite that, it is thought that during the Middle Holocene these forests occupied a larger area than today. Against this background, the following research question was posed: Have the environmental requirements that delimit the current distribution of sacred fir in the Trans-Mexican Volcanic Belt changed in 6 000 years? In order to answer this question, the following particular objectives were formulated: 1) to delimit and compare the historical and current distribution of A. religiosa forest in the Trans-Mexican Volcanic Belt using potential distribution modeling, 2) to determine and compare the environmental variables that delimit the ecological niche of the species in both periods, and 3) to estimate the historical and current sacred fir forest area in the protected natural areas of the study area.
Based on the climatic changes that occurred between the current and historical (middle Holocene) periods, and according to the ecological niche conservatism theory (Peterson, 2011b), it could be predicted that the potential distribution of A. religiosa in the Trans-Mexican Volcanic Belt is similar for both periods.
Materials and methods
Study area
The Trans-Mexican Volcanic Belt is located at coordinates 17° 30’ and 20° 25’ N, and 96° 20’ and 105° 20’ W (Ferrusquía-Villafranca, 1998) (Figure 1); it has elevations ranging from 256 to 5 650 m (Instituto Nacional de Estadística y Geografía [INEGI], 2018). The area’s topography is rugged and steep in the mountains (Ferrusquía-Villafranca, 1998). The dominant climate is temperate (humid and sub-humid) with an annual mean temperature of 12 to 18 °C and annual precipitation of 600 to 1 500 mm (García, 1998).

Species description
Abies religiosa is a monoecious, evergreen tree, with a height of up to 60 m and maximum diameters of 1.80 m. The tree has the following characteristics: greyish, rough and cracked bark 18-25 mm thick; cross-shaped branches; simple and alternate leaves 15-35 mm long by 1.5 mm wide; and solitary cones 8-16 cm long and 4-6 cm wide (Protectora de Bosques [PROBOSQUE], 2007; Rzedowski, 2006).
In Mexico, the species is located in Morelos, Estado de Mexico, Hidalgo, Puebla, Michoacán, Jalisco, Colima, Guerrero, Tlaxcala, Veracruz and Mexico City (Martínez-Méndez et al., 2016) (Figure 2). In the Trans-Mexican Volcanic Belt, A. religiosa is distributed in an altitudinal gradient from 2 400 to 3 500 m and is the dominant species of the vegetation type known in Mexico as oyamel (sacred fir) forest (Madrigal, 1967; Rzedowski, 2006).

Abies religiosa records
A total of 1 042 A. religiosa records were downloaded from the Niche Toolbox platform (Osorio-Olvera, Vijay, Narayani, Soberón, & Falconi, 2017). In order to avoid spatial autocorrelation and overfitting of the models (Monterrubio-Rico et al., 2016; Peterson & Nakazawa, 2008), duplicate, poorly georeferenced, and close coordinates (distance >1 km) were removed, leaving one record for each cell of 1 km2. At the end of the depuration process, 341 records of A. religiosa distributed in the study area were obtained. These were used to generate the potential distribution models for the current and historical periods.
Current and historical bioclimatic information
Table 1 shows the 19 bioclimatic variables available in WorldClim version 2.0 for the current period (1970-2000) (Fick & Hijmans, 2017) and the global circulation models (GCMs) CNRMCM5 (National Center for Meteorological Research of France) and MIROC_ESM (Institute of Oceanic and Atmospheric Research, National Institute of Environmental Studies and the Japanese Agency for Marine-Earth Science and Technology) for the Middle Holocene. The latter were generated from the CMIP5 (Coupled Model Intercomparison Project Phase 5, 2013) Regional Models of the Intergovernmental Panel on Climate Change (IPCC). All variables had a spatial resolution of 1 km2 and were in Geotiff (TIF) format. In addition, the variables soil type (SUE; scale 1: 250 000) and elevation (DEM; digital elevation model, 90-m resolution) were included, which were downloaded in vector and raster format from the INEGI platform (INEGI, 2014, 2018). At the end, all variables were homogenized to ASCII format with a scale of 1 km2.
Table 1 Bioclimatic variables used in the generation of current and historical potential distribution models of Abies religiosa in the Trans-Mexican Volcanic Belt.
| Bioclimatic variables | Key |
|---|---|
| Annual mean temperature (°C) | BIO1 |
| Diurnal temperature oscillation (°C) | BIO2 |
| Isothermality [(BIO2/BIO7)*100] | BIO3 |
| Temperature seasonality (coefficient of variation, %) | BIO4 |
| Mean maximum temperature of warmest period (°C) | BIO5 |
| Mean minimum temperature of coldest period (°C) | BIO6 |
| Temperature annual oscillation (°C) | BIO7 |
| Mean temperature of wettest quarter (°C) | BIO8 |
| Mean temperature of driest quarter (°C) | BIO9 |
| Mean temperature of warmest quarter (°C) | BIO10 |
| Mean temperature of coldest quarter (°C) | BIO11 |
| Cumulative annual precipitation (mm) | BIO12 |
| Precipitation of wettest period (mm) | BIO13 |
| Precipitation of driest period (mm) | BIO14 |
| Precipitation seasonality (coefficient of variation, %) | BIO15 |
| Precipitation of wettest quarter (mm) | BIO16 |
| Precipitation of driest quarter (mm) | BIO17 |
| Precipitation of warmest quarter (mm) | BIO18 |
| Precipitation of coldest quarter (mm) | BIO19 |
Selection of variables
Spatial autocorrelation was eliminated through a multicollinearity analysis (Monterrubio-Rico et al., 2016; Peterson & Nakazawa, 2008), where those variables with a correlation coefficient ?#8805;0.85 (Manzanilla et al., 2019; Monterrubio-Rico et al., 2016) were discarded to maximize the contribution of variables in the distribution models (Martínez-Méndez et al., 2016; Monterrubio-Rico et al., 2016; Peterson & Nakazawa, 2008).
Delimitation of area M
Modeling area M is the environmental geographic space where a species has been reported or where it is supposed to be in accordance with the available biological knowledge (Martínez-Méndez et al., 2016; Soberón & Peterson, 2005). To delimit area M, the physiographic subprovinces were used where A. religiosa records were located: Plains and Sierras of Querétaro and Hidalgo, Chapala, Mil Cumbres, Lakes and Volcanoes of Anáhuac, Neovolcanica Tarasca and Volcanoes of Colima (INEGI, 2001). Area M was used to cut the variables to the same size and thus avoid the generation of overestimated distribution areas (Monterrubio-Rico et al., 2016).
Generation of distribution models
The distribution models were generated with the presence records of A. religiosa in CSV format and with the variables BIO1, BIO2, BIO3, BIO4, BIO12, BIO14, BIO15, BIO18, BIO19 in ASCII format (Table 1), soil type (SUE), and elevation (DEM).
The variables were introduced into the maximum entropy program (MaxEnt), where 75 % of the species records were used to perform the training test and the remaining 25 % for the validation test of the models (Martínez-Méndez et al., 2016; Monterrubio-Rico et al., 2016; Phillips, Anderson, & Schaphire, 2006). An internal replication was applied by cross validation and the Extrapolate and Do clamping boxes were disabled to avoid overfitting of the models (Elith et al., 2011). The outputs of the models were of the logistic type, which represents an environmental probability index with values from 0 to 1. Values close to 0 indicate unsuitable environmental conditions, while values close to 1 suggest excellent environmental conditions for the growth and development of the species (Coitiño, Montenegro, Fallabrino, González, & Hernández, 2013; Phillips et al., 2006).
Five distribution models were generated and tested, in order to determine by means of ROC (analysis of the performance characteristics of the receiver of the area under the curve [AUC]), Partial Roc and Z tests, which model fitted the current distribution of the species (Table 2). The model with best statistical performance was overlapped on the federal protected natural areas shapefile decreed for the region (Comisión Nacional de Áreas Naturales Protegidas [CONANP], 2017). The current sacred fir area within the polygonal of the protected natural areas was estimated with ArcGIS 10.3 software (Environmental Systems Research Institute [ESRI], 2014).
The parameters of the current distribution model with the best statistical fit (Morrone & Escalante, 2016) were transferred to the maximum entropy (MaxEnt) program to generate distribution models in the past.
Table 2 Parameters used in the generation of potential distribution models of Abies religiosa in the Trans-Mexican Volcanic Belt.
| Model | Internal replication | Threshold application rule | Replicates |
|---|---|---|---|
| M1 | Cross validation | Equal training sensitivity and specificity | 1 000 |
| M2 | Cross validation | Maximum training sensitivity plus specificity | 1 000 |
| M3 | Cross validation | Test of equal sensitivity and specificity | 1 000 |
| M4 | Cross validation | Maximum test sensitivity plus specificity | 1 000 |
| M5 | Cross validation | No application of threshold rule | 500 |
Validation of the models
Models were evaluated using the AUC values of the ROC, where values between 0.7 to 0.9 are considered good and those greater than 0.9 are considered excellent (Coitiño et al., 2013; Peterson et al., 2011). Nevertheless, this type of validation has been questioned for not considering true absences (Lobo, Jiménez, & Real, 2008; Peterson, Papes, & Soberón, 2008), so it was necessary to perform a Partial Roc analysis in the Tool for Partial Roc version 1.0 software (Narayani, 2008) to counteract AUC shortcomings (Peterson et al., 2008).
The recommendations of Peterson et al. (2008) were followed by using 1 000 replicates per bootstrap between presence and habitat suitability files and establishing a 5 % omission error. The test generates values from 1 to 2, where a value with an average radius of 1.0 is equivalent to a random model (Garza-López et al., 2016; Lobo et al., 2008; Peterson et al., 2008). To determine whether the models were statistically valid, a Z test was performed between the AUC ratios of Partial ROC (Martínez-Méndez et al., 2016). The best model was chosen based on the highest value of the Partial ROC tests, lowest standard deviation, and a reliable Z value (P < 0.01).
The logistic output values of the models were reclassified into three habitat quality categories with equal intervals (low, moderate, and high), with the ArcMap 10.3 program reclassify tool (ESRI, 2014). The values of the high habitat quality category were used to transform the continuous models to binary models (suitable-unsuitable). Based on the reclassification, the potential distribution area of A. religiosa in the Trans-Mexican Volcanic Belt was estimated for both analyzed periods.
Projections of the models with the best statistical performance for each period were visualized and drawn in potential distribution maps with the ArcMap 10.3 program (ESRI, 2014). Finally, the most important variables of each analyzed period were determined with the Jackknife test, which allowed quantifying the contribution of the environmental variables in the distribution models (Phillips et al., 2006).
Results and discussion
The AUC results from the ROC test of the current models obtained values greater than 0.9 (training and validation data) for both tests, like the models for the middle Holocene. These values indicate that the performance of the models generated for both periods was excellent (>0.9; Coitiño et al., 2013; Monterrubio-Rico et al., 2016; Peterson et al., 2011); therefore, the distribution models were considered reliable.
The performance of the five models was similar for the ROC test; however, when applying the Partial Roc test, the model with the best statistical fit and lowest standard deviation was model 2 (Table 3).
Table 3 Partial ROC and Z results of the historical and current potential distribution models of Abies religiosa in the Trans-Mexican Volcanic Belt.
| Period analyzed | Model | Partial ROC average ratios | Standard deviation | Z test |
|---|---|---|---|---|
| Current | M1 | 1.494 | 0.129 | P < 0.01 |
| M2 | 1.500 | 0.123 | P < 0.01 | |
| M3 | 1.494 | 0.128 | P < 0.01 | |
| M4 | 1.492 | 0.130 | P < 0.01 | |
| M5 | 1.475 | 0.134 | P < 0.01 | |
| Historical | CRNMCM5 | 1.551 | 0.114 | P < 0.01 |
| MIROC-ESM | 1.496 | 0.138 | P < 0.01 |
The models indicated that the current potential distribution (1970-2000) of A. religiosa in the Trans-Mexican Volcanic Belt is 194 387.3 ha. FAO (2010) and SEMARNAT (2007) report 144 000 ha, which indicates that, after 2000, there was a reduction of 50 387.3 ha of sacred fir forest area. In addition, it was found that 86.5 % (168 148 ha) of the current potential distribution of sacred fir is within protected natural areas of the Trans-Mexican Volcanic Belt. This result coincides with what was mentioned by Rzedowski (2006) and Martínez-Méndez et al. (2016), who suggest that A. religiosa is the only Mexican fir species that presents most of its distribution in the high and humid zones within natural areas of the Trans-Mexican Volcanic Belt, which has provided (in some measure) the legal protection to this species.
According to the results of the historical models, the estimated forest area of A. religiosa was 190 466.2 ha (CNRMCM5) to 193 563.2 ha (MIROC_ESM), which translates into 49 563.2 to 46 466.2 ha more than the current area reported by SEMANART (2007) and FAO (2010), respectively, but 824.1 ha (MIROC_ESM) and 3 921.1 ha (CNRMCM5) less than estimated in the present study (Figures 3 and 4). Based on the results of the binomial ROC, Partial ROC and Z tests (P < 0.01), we selected the CRNMCM5 model, which presented the best fit (Table 3), where values close to 2 indicated that the statistical fit of the model was reliable as mentioned by Garza-López et al. (2016), Narayani (2008) and Peterson et al. (2008).
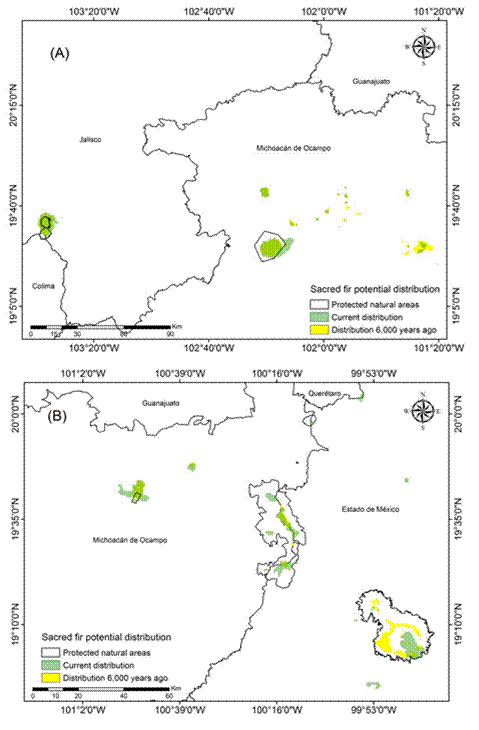
Figure 3 Current (1970-2000) and historical potential distribution of the sacred fir (Abies religiosa) in the Trans-Mexican Volcanic Belt: West zone (A) and Center I zone (B).
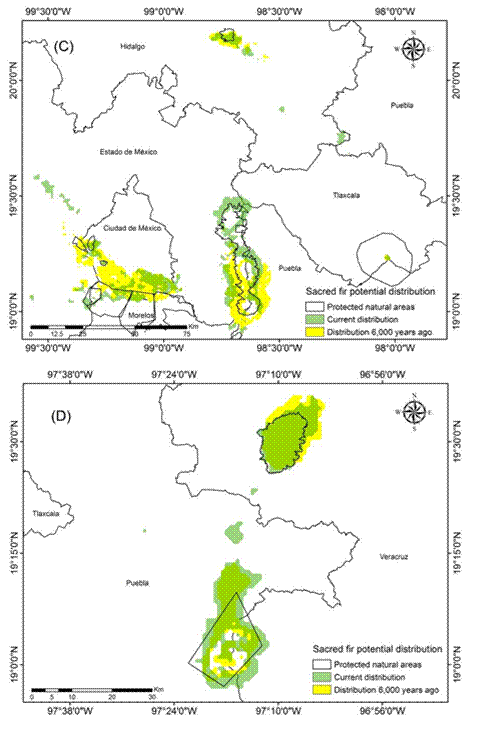
Figure 4 Current (1970-2000) and historical potential distribution of the sacred fir (Abies religiosa) in the Trans-Mexican Volcanic Belt: Center II zone (C) and East zone (D).
Historical and current potential distribution
Given the recent diversification of the genus Abies in the world (Xiang et al., 2015), it is very likely that climatic conditions of 6 000 to 12 000 years ago were warmer (+2 °C) in North America’s temperate and cold areas (Caballero et al., 2010; Svensson et al., 2008). This possibly delimited the distribution of coniferous forests to the temperate and cold zones of the Trans-Mexican Volcanic Belt (Caballero et al., 2010; Madrigal, 1967; Martínez-Méndez et al., 2016; Rzedowski, 2006).
According to the Jackknife test, the most important limiting environmental variables in the distribution of A. religiosa in the Trans-Mexican Volcanic Belt were the elevation (DEM with 56.5 %), cumulative annual precipitation (BIO12 with 18.4 %) and precipitation of the warmest quarter (BIO18 with 8.3 %). These three variables explained 83.2 % of the model variability. These results are similar to those found by Farjon and Filer (2013), Madrigal (1967), and Rzedowski (2006), who indicate that constant annual humidity and abundant rainfall in summer are essential for the distribution of this fir. The present study found that the ideal altitudinal gradient for the growth and development of A. religiosa was from 2 129 to 3 687 m. These results are similar to those reported in the literature, since previous studies mention that the distribution of A. religiosa ranges from 2 400 to 3 660 m (Calderón de Rzedowski & Rzedowski, 2001; Madrigal, 1967; Nieto de Pascual-Pola, 1995, Rzedowski, 2006). It should be noted that, according to the results of this study, the distribution of the sacred fir forest can be found 271 m below the minimum elevation and 27 m above the maximum elevation reported in the literature. Regarding cumulative annual precipitation, the current models estimated an average of 1 125 mm. This value was slightly higher than that reported by Madrigal (1967) and similar to that mentioned by Rzedowski (2006) (1 000 mm and >1 000 mm, respectively). Martínez-Méndez et al. (2016) and Sáenz-Romero et al. (2012) mention that precipitation during the summer months is a determining factor in the distribution of the species, which agrees with what was obtained in the present study. However, dendrochronological studies carried out for the species within the study area indicate that winter-spring precipitation is of high relevance for the growth of A. religiosa (Cerano-Paredes et al., 2014; Huante, Rincón, & Swetnam, 1991). This differs from what was obtained in this study, since the evaluated models did not consider the winter-spring precipitation as a key factor in sacred fir distribution. However, this variable could be of greater relevance than precipitation in the summer months during the radial growth stage of the species.
On the other hand, the most important variables in the prediction of historical distribution (MIROC_ESM model) were annual mean temperature (BIO1 with 65.9 %), cumulative annual precipitation (BIO12 with 18.6 %), and diurnal temperature oscillation (BIO2 with 4.6 %); these three variables accounted for 89.1 % of the model’s variation. For the CNRMCM5 model, the most important variables were annual mean temperature (BIO1 with 62.4 %), cumulative annual precipitation (BIO12 with 16.2 %), and diurnal temperature range (BIO2 with 6.5 %), which contributed with 85.1 % of the model. The results of the present study agree with those of Madrigal (1967) and Rzedowski (2006), who mention that diurnal temperature range (11 to 16 °C) is a determining factor for the growth of A. religiosa. Therefore, it was corroborated that this variable has been a limiting factor for 6 000 years (10.6 to 17.2 °C) in the distribution of sacred fir in the Trans-Mexican Volcanic Belt. With respect to the annual mean temperature, the values between 4.6 and 15.7 °C are congruent with those reported by Rzedowski (2006), who mentions that they fluctuate between 7 and 15 °C. Otherwise, according to the climatic conditions estimated by the global circulation models, annual precipitation during the middle Holocene was 80 to 224 mm higher than today’s and the annual mean temperature was 1 °C colder and not warmer as mentioned by Svensson et al. (2008) and Caballero et al. (2010).
Because the estimated areas and environmental conditions described in the literature and in this study showed similarities, it can be assumed that the potential distribution of A. religiosa has remained stable over time (according to the theory of ecological niche conservatism; Peterson, 2011b). Thus, the evidence found in this study seems to indicate that the distribution of the species in the Trans-Mexican Volcanic Belt has not changed in 6,000 years. This would prove (with the limitation that there are no palynological records) that the ecological niche of A. religiosa has been maintained during the evaluated periods. It should be noted that, added to the fact that the current and historical distribution did not change significantly, it was possible to determine that sites such as Nevado de Toluca and Mexico City had larger sacred fir areas during the past: +2 658.4 ha and +20 129.7 ha, respectively (Table 4; Figure 5A). Consequently, it can be inferred that the climatic conditions of 6,000 years ago were colder and wetter in the temperate and cold mountainous areas of the Trans-Mexican Volcanic Belt, which caused the sacred fir distribution to remain there until the present (Madrigal, 1967; Martínez-Méndez et al., 2016; Ramírez-Barahona & Eguiarte, 2013; Rzedowski, 2006).
Table 4 Sites with the greatest historical potential distribution of A. religiosa forest in the Trans-Mexican Volcanic Belt.
| Site | Current potential distribution (ha) | Past potential distribution (ha) | Change (%) |
|---|---|---|---|
| Nevado de Toluca | 6 847.8 | 9 506.2 | -28 |
| Mexico City | 19 340.9 | 39 470.6 | -51 |

Conclusions
This study represents a first attempt at estimating the potential distribution of Abies religiosa 6 000 years ago in the Trans-Mexican Volcanic Belt. These results provide valuable information on the environmental variables (elevation, annual precipitation, summer precipitation, annual mean temperature, and diurnal temperature oscillation) that have limited the distribution of this species. Six thousand years ago, the climatic conditions corresponding to the distribution of the sacred fir within the Trans-Mexican Volcanic Belt were colder and wetter than today. Approximately 86.5 % of the distribution of A. religiosa is found within protected natural areas of the zone; however, this protection does not make it immune to natural and anthropogenic disturbances. Although the historical and current distribution of sacred fir in the Trans-Mexican Volcanic Belt was similar, sites such as Nevado de Toluca and Mexico City were found to have wider areas in the past.

