










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.23 no.2 Chapingo may./ago. 2017
https://doi.org/10.5154/r.rchscfa.2017.01.001
Artículos científicos
Supervivencia y crecimiento de tres especies de Quercus bajo condiciones contrastantes de cobertura en el sur de México
1El Colegio de la Frontera Sur. Carretera Panamericana y Periférico Sur s/n. C. P. 29290. San Cristóbal de Las Casas, Chiapas, México.
2Universidad Nacional Autónoma de México, Facultad de Ciencias. Ciudad Universitaria, Circuito exterior s/n. C. P. 04510. Coyoacán, Ciudad de México, México.
Introducción:
El aprovechamiento intensivo modifica la composición y estructura de los bosques del sur de México, limitando la repoblación natural de especies de Quercus.
Objetivos:
Se evaluó la factibilidad de Quercus crispipilis, Q. ocoteifolia y Q. segoviensis en la restauración bajo tres condiciones de dosel: bosque secundario de pino-encino, matorral y pastizal.
Materiales y métodos:
La supervivencia, el crecimiento y la producción de biomasa de plantas jóvenes de las tres especies de Quercus se determinaron en tres condiciones, con tres repeticiones cada una, durante 14 meses. Se trasplantaron 33 individuos de cada especie por repetición.
Resultados y discusión:
La supervivencia fue relativamente alta (> 88 %) en las tres condiciones. La tasa relativa de crecimiento (TRC) en altura de Q. crispipilis y Q. ocoteifolia fue mayor en las condiciones de bosque y matorral. La TRC en diámetro basal de las tres especies fue mayor en el pastizal, al igual que la biomasa de las raíces de Q. crispipilis y Q. segoviensis.
Conclusión:
La presencia de dosel influye en las variables microclimáticas de los sitios. Quercus crispipilis y Q. segoviensis tienen supervivencia y crecimiento mayores en el matorral y pastizal, mientras que Q. ocoteifolia resulta favorecida bajo dosel forestal.
Palabras clave: Altos de Chiapas; bosque de pino-encino; matorral de Baccharis; pastizal inducido; restauración forestal
Introduction:
Intensive use modifies the composition and structure of the forests of southern Mexico, limiting the natural repopulation of Quercus species.
Objective:
The feasibility of Quercus crispipilis, Q. ocoteifolia and Q. segoviensis in forest restoration was evaluated under three canopy conditions: secondary pine-oak forest, shrubland and grassland.
Materials and methods:
Survival, growth and biomass production of young plants of the three Quercus species were determined in three conditions, with three replications each, for 14 months. A total of 33 individuals of each species were transplanted per replication.
Results and discussion:
Survival was relatively high (> 88 %) in all three conditions. The relative growth rate (RGR) in height of Q. crispipilis and Q. ocoteifolia was higher under forest and shrubland conditions. RGR in basal diameter of the three species was higher under grassland conditions, as was the biomass of Q. crispipilis and Q. segoviensis roots.
Conclusions:
The presence of canopy influences the microclimatic variables of the sites. Quercus crispipilis and Q. segoviensis have higher survival and growth under shrubland and grassland conditions, while Q. ocoteifolia is favored under forest canopy.
Keywords: Chiapas highlands; pine-oak forest; Baccharis shrubland; induced grassland; forest restoration
Introducción
La deforestación por cambio de uso del suelo y el aprovechamiento intensivo de algunas especies arbóreas modifican el funcionamiento y la estructura de los ecosistemas forestales, por lo que su recuperación mediante procesos naturales no siempre es posible (Bustamante, Badano, & Pickett, 2012; Cayuela, Rey-Benayas, & Echeverría, 2006; Ramírez-Marcial, Camacho-Cruz, & González-Espinosa, 2008). A escala local, los cambios se perciben tanto a nivel de las condiciones microclimáticas y edáficas como en la modificación de otras interacciones bióticas que afectan conjuntamente la germinación de semillas y el establecimiento de plantas (Bonfil & Soberón, 1999; Ramos-Palacios & Badano, 2014). En la región de Los Altos de Chiapas, el aprovechamiento selectivo de árboles ha provocado la reducción en la densidad y dominancia de árboles adultos, principalmente de encinos (Quercus spp.), promoviendo la regeneración y dominancia de los pinos (Alba-López, González-Espinosa, Ramírez-Marcial, & Castillo-Santiago, 2003; Galindo-Jaimes, González-Espinosa, Quintana-Ascencio, & García-Barrios, 2002).
La restauración de los ecosistemas forestales degradados se hace con la intención de recuperar su estructura, función y productividad, así como restablecer los procesos ecológicos y los servicios ecosistémicos que representan (Lamb, Stanturf, & Madsen, 2012; Stanturf, Palik, Williams, Dumroese, & Madsen, 2014; Torres-Miranda, Luna-Vega, & Oyama, 2011). La reforestación con múltiples taxones es una opción viable cuando las especies leñosas no pueden establecerse de manera natural; este proceso permite reconstruir la estructura de la flora leñosa (Ramírez-Marcial et al., 2008).
La densidad de la vegetación, las condiciones del medio abiótico local y el régimen de disturbio existente son algunos factores que limitan el establecimiento de las especies (Guo, Wang, Zhu, Wang, & Guo, 2011). Por lo anterior, para fines de restauración, es necesario entender el efecto de las condiciones climáticas, edáficas y bióticas del sitio sobre la supervivencia y el crecimiento de las especies utilizadas (Cardillo & Bernal, 2006; Pulsford, Lindenmayer, & Driscoll, 2016). Algunas especies de Quercus, debido a su plasticidad fenotípica, tienen potencial para ser utilizadas en proyectos de restauración forestal bajo diferentes condiciones edáficas (González-Espinosa et al., 2012) y de dosel (Cardillo & Bernal, 2006; Ramírez-Marcial, Camacho-Cruz, González-Espinosa, & López-Barrera, 2006; Sánchez-Velásquez, Ramírez-Bamonde, Andrade-Torres, & Rodríguez-Torres, 2008). No obstante, los daños físicos causados por el pastoreo (Ramírez-Marcial, González-Espinosa, & García-Moya, 1996; Sánchez-Velásquez, Domínguez-Hernández, Pineda López, & Lara-González, 2011), el estrés hídrico, el grado de perturbación ambiental, las condiciones del sustrato (Bonfil & Soberón, 1999; Flores-Cano, Badano, & Flores, 2012), y el tamaño y vigor de las plántulas (Bonfil, Rodríguez de la Vega, & Peña, 2000; Ramírez-Contreras & Rodríguez-Trejo, 2004) son factores que afectan la supervivencia y crecimiento.
Una alternativa que facilita el establecimiento de las plantas es el uso de arbustos o árboles como nodrizas, ya que mejoran las condiciones microclimáticas y favorecen el crecimiento inicial (Bonfil & Soberón, 1999; Ramírez-Contreras & Rodríguez-Trejo, 2009; Ramírez-Marcial et al., 1996). Por tanto, el análisis del desempeño de las plantas a lo largo de un gradiente ambiental, asociado al disturbio humano, permite identificar algunas barreras que impiden el establecimiento en sitios con disponibilidad limitada de propágulos (Ramírez-Marcial et al., 2008; Ramos-Palacios & Badano, 2014).
El objetivo de esta investigación fue evaluar la supervivencia y el crecimiento de tres especies de encinos introducidas en tres condiciones de dosel: bosque, matorral y pastizal. Las tres especies, características de los ecosistemas de montaña del sur de México y Guatemala, se encuentran en alguna categoría de riesgo de extinción (González-Espinosa, Meave, Lorea-Hernández, Ibarra-Manríquez, & Newton, 2011; Ramírez-Marcial et al., 2010). Quercus ocoteifolia Liebm., a diferencia de Q. crispipilis Trel. y Q. segoviensis Liebm., es perennifolia y requiere condiciones de cobertura de dosel, temperaturas frescas y mayor humedad para el establecimiento (González-Espinosa et al., 2011; Gutiérrez & Trejo, 2014; Ramírez-Marcial et al., 2010). Por ello se espera que la supervivencia de Q. ocoteifolia sea mayor bajo el dosel del bosque, mientras que la de Q. crispipilis y Q. segoviensis lo sea en condiciones más abiertas sin dosel.
Materiales y métodos
Área de estudio
El estudio se realizó en el Parque Ecológico El Encuentro (PEE) ubicado en la porción noreste de San Cristóbal de Las Casas, Chiapas (16° 43’ 54.72” - 16° 44’ 08.38” LN y 92° 38’ 52.59” - 92° 38’ 24.52” LO) a una altitud promedio de 2,270 m. La precipitación promedio anual es 1090.5 mm con temperatura media anual de 15.0 °C (Comisión Nacional del Agua [CONAGUA], 2017). El PEE estuvo bajo aprovechamiento forestal de Pinus para producción de madera y de Quercus spp. y otras especies latifoliadas para leña; también fue sitio de apacentamiento de ovinos hasta el año 2010. El suelo es moderadamente profundo, derivado de rocas calcáreas y correspondiente a los grupos Rendzina y Luvisol. La vegetación actual comprende bosques secundarios dominados por Pinus pseudostrobus Lindl., P. tecunumanii F. Schwerdtf. ex Eguiluz & J. P. Perry, Q. segoviensis, Q. crispipilis y Q. rugosa Née (De la Mora-Estrada, Ruiz-Montoya, Ramírez-Marcial, Morón-Ríos, & Mayorga-Martínez, 2017).
En el estudio se evaluaron tres condiciones de cobertura de dosel: bosque de pino-encino, matorral de Baccharis vaccinioides Kunth y pastizal. En cada condición se establecieron tres parcelas de 100 a 150 m2 que se consideraron repeticiones experimentales. El tamaño varió en función de la disponibilidad del terreno, dependiendo de la homogeneidad de la cobertura; las parcelas del pastizal fueron las más pequeñas y las del bosque las más grandes. En cada parcela se caracterizaron las condiciones de cobertura del dosel, humedad del suelo y temperatura del suelo y aire. La cobertura de dosel se obtuvo del análisis de seis fotografías hemisféricas por parcela, tomadas a 1 m de altura con un lente hemisférico (OptekaTM 0.20x, EUA) adherido a una cámara digital (Nikkon® modelo D5200, Singapur) y procesadas con el programa HemiView (Rich, Wood, Vieglais, Burek, & Webb, 1999). La humedad y temperatura del suelo se registraron con 20 lecturas por parcela en la temporada de lluvias (octubre 2015) y 20 en la temporada de sequía (abril 2016). La humedad se midió con el Theta Meter® (modelo HH1, EUA) y la temperatura con un termómetro digital de doble salida tipo J/K Extech Instruments® (modelo 421502, EUA). La temperatura del aire de cada condición se registró continuamente a un metro de altura del suelo con un sensor HOBO TM® (Onset Computer Corporations, EUA).
Diseño de la plantación
Se utilizaron plantas de Q. crispipilis, Q. ocoteifolia y Q. segoviensis provenientes de semillas colectadas en noviembre y diciembre de 2013 y germinadas en los viveros forestales de El Colegio de la Frontera Sur (ECOSUR) en San Cristóbal de Las Casas. Quercus crispipilis y Q. segoviensis se distribuyen en Chiapas y Guatemala formando asociaciones de bosques de pino-encino entre los 1,800 y 2,400 m de altitud, en tanto que Q. ocoteifolia se distribuye en Oaxaca y Chiapas formando asociaciones de pino-encino y bosque mesófilo de montaña en lugares más húmedos y frescos, entre los 2,000 y 2,600 m (Ramírez-Marcial et al., 2010). Las plantas se mantuvieron durante 15 meses en el vivero y dos meses en el exterior para su aclimatación. No se aplicó poda de raíz en vivero. Las tres especies se trasplantaron con cepellón durante la última semana de julio de 2015, para aprovechar el periodo de crecimiento (Ramírez-Marcial et al., 2006). En cada parcela se colocaron 33 plantas por especie, distribuidas aleatoriamente a 1 m de equidistancia. La cantidad por parcela se determinó con base en la disponibilidad de plantas de las tres especies estudiadas.
Variables de medición
Una semana después del trasplante se midieron la altura y diámetro de cada planta; estos valores se consideraron como el punto de inicio del experimento. Las evaluaciones de supervivencia y crecimiento se repitieron a los 2, 4, 7, 9, 12 y 14 meses posteriores al trasplante. El crecimiento individual se midió a través de la altura máxima y el diámetro basal del tallo. Con estos valores se calcularon las tasas relativas de crecimiento (TRC) de ambas variables mediante la fórmula utilizada por Hunt et al. (2002): TRC = [ln crecimiento final (cm) - ln crecimiento inicial (cm)] / tiempo de evaluación (meses).
Al término de la última evaluación (septiembre de 2016), la biomasa acumulada en tallos, hojas y raíces se cuantificó en una muestra aleatoria de cinco plantas por especie y repetición (135 plantas en total). Las plantas extraídas se pesaron por componente; hojas, tallo y raíz. Posteriormente, las fracciones se colocaron en estufa a 70 °C durante 72 h para su desecación. Al final de este periodo, las muestras se pesaron nuevamente en una balanza con precisión de 0.01 g, para obtener el peso seco.
Análisis de datos
Se utilizó un diseño completamente al azar con arreglo factorial 3 x 3 (tipo de cobertura y especies). El cumplimiento de los supuestos estadísticos de normalidad, homocedasticidad e independencia se verificó. Las variables ambientales entre condiciones se analizaron mediante la prueba de Kruskal-Wallis; y la comparación entre pares de tipos de cobertura, con la prueba de rangos de Wilcoxon. La proporción de plantas supervivientes en cada condición se analizó con la prueba no paramétrica de rangos logarítmicos de Kaplan-Meier (Crawley, 2013). Las TRC, 14 meses después del trasplante, se analizaron con ANOVA considerando a las especies y la condición como factores principales. Una vez que se mostraron diferencias significativas entre especies (P < 0.05), se utilizó otro ANOVA y se evaluó el efecto de la condición en cada especie. Las diferencias en la biomasa acumulada por fracción de cada especie, entre condiciones, también se evaluaron con ANOVA. Todos los análisis se realizaron con el programa R versión 3.2.2 (R Development Core Team, 2015).
Resultados y discusión
Características microclimáticas de los sitios
El Cuadro 1 presenta las condiciones microclimáticas en las tres coberturas de estudio. Las variables difirieron entre los sitios de trasplante y entre temporadas del año; las temperaturas del suelo fueron significativamente (P < 0.001) más frescas en el bosque. En el otoño, la humedad del suelo fue similar (P = 0.32) en los tres sitios; sin embargo, al inicio de la primavera se observaron diferencias significativas (P < 0.001) con menor humedad en el matorral que en el bosque y el pastizal.
Cuadro 1 Temperatura y porcentaje de humedad del suelo en tres condiciones de cobertura en San Cristóbal de Las Casas, Chiapas.
| Variable microclimática | Periodo | Cobertura | Kruskal-Wallis X 2 | Valor P | ||
|---|---|---|---|---|---|---|
| Bosque | Matorral | Pastizal | ||||
| Temperatura del suelo (°C) | Octubre 2015 | 15.9 ± 0.05 a | 16.6 ± 0.12 b | 18.8 ± 0.08 c | 122.03 | P < 0.001 |
| Abril 2016 | 18.2 ± 0.18 a | 20.2 ± 0.20 b | 22 ± 0.24 c | 94.53 | P < 0.001 | |
| Humedad del suelo (%) | Octubre 2015 | 64 ± 1.59 a | 61 ± 1.83 a | 65 ± 1.63 a | 2.24 | P = 0.32 |
| Abril 2016 | 31 ± 1.43 a | 23 ± 1.03 b | 28 ± 1.47 a | 12.93 | P < 0.001 | |
Letras diferentes denotan diferencias significativas entre sitios de cobertura para cada periodo de evaluación (P < 0.001). ± error estándar de la media.
Por otra parte, la temperatura media del aire fue significativamente más baja (P < 0.05) en el bosque y el matorral que en el pastizal (Figura 1). Con respecto a la radiación fotosintéticamente activa (RFA), los valores fueron significativamente (P = 0.05) mayores en el pastizal, intermedios en el matorral e inferiores en el bosque (Figura 2).
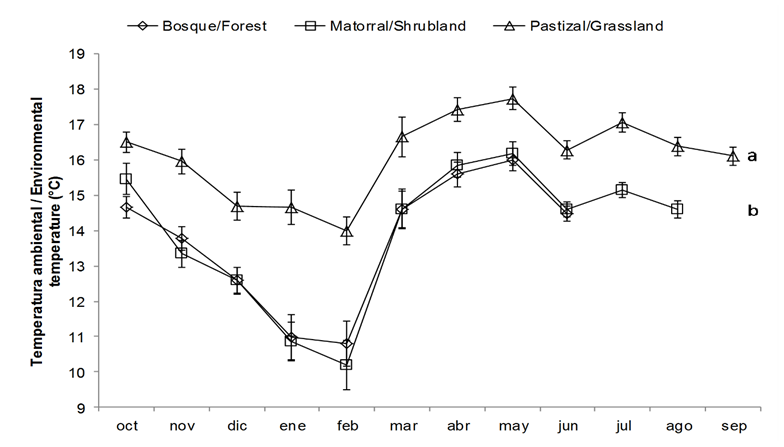
Figura 1 Temperatura del aire registrada a 1 m sobre el nivel del suelo (octubre 2015-septiembre 2016) en San Cristóbal de Las Casas, Chiapas. Cada punto corresponde al valor medio mensual (± error estándar). Letras diferentes denotan diferencias significativas entre coberturas mediante la prueba de Wilcoxon (P < 0.05), posterior al análisis de Kruskal-Wallis.

Figura 2 Perfil de la distribución mensual de la radiación fotosintéticamente activa en tres condiciones de cobertura (enero 2016-diciembre 2016) en San Cristóbal de Las Casas, Chiapas. Letras diferentes denotan diferencias significativas entre coberturas mediante la prueba de Wilcoxon (P < 0.05). ± error estándar de la media.
Los resultados indican que las condiciones microclimáticas varían en función del tipo de cobertura, lo cual puede afectar el desempeño de las plantas (Dickson, 1990). La ausencia de dosel se refleja en temperatura, radiación y evapotranspiración mayores, propiciando considerable estrés y desecación en el suelo, y menor crecimiento de las plantas (Arosa, Ceia, Costa, & Freitas, 2015; Ramírez-Marcial et al., 2008).
Supervivencia de Quercus
La supervivencia de las especies después de 14 meses de evaluación fue alta en las tres condiciones de estudio (> 90 %), con excepción de Q. ocoteifolia (88 %) en el pastizal (Figura 3). El periodo de seguimiento del estudio permite afirmar que las especies de Quercus tuvieron la habilidad para establecerse efectivamente en las tres condiciones; sin embargo, estas tendencias pueden modificarse con base en la estacionalidad o la etapa de crecimiento de las plantas utilizadas (Alvarez-Aquino & Williams-Linera, 2012; Espelta, Riba, & Retana, 1995).
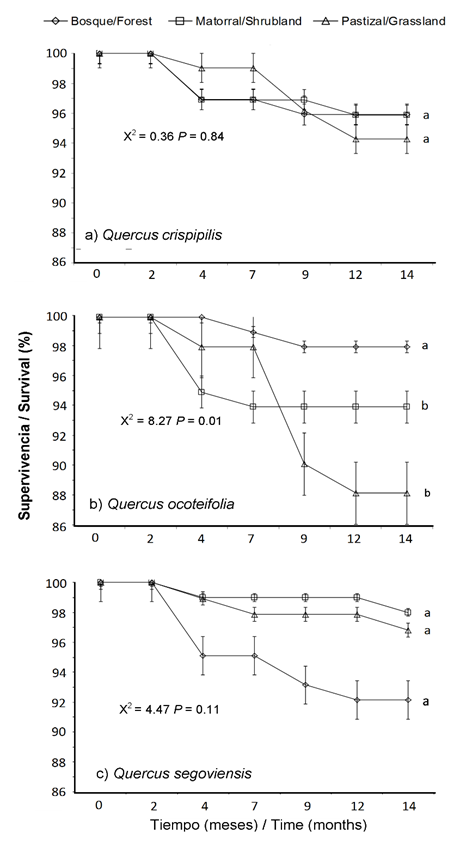
Figura 3 Curvas de supervivencia de tres especies de Quercus durante un periodo de 14 meses a partir del trasplante en condiciones de bosque, matorral y pastizal en San Cristóbal de Las Casas, Chiapas. Letras diferentes indican diferencias significativas entre las curvas de supervivencia mediante la prueba no paramétrica de rangos logarítmicos de Kaplan-Meier (P < 0.05).
En estudios de corto plazo se ha demostrado que la supervivencia está influenciada por las condiciones microclimáticas creadas por la presencia de árboles, arbustos y especies nativas de sucesión temprana. Tales circunstancias pueden modificar las condiciones microbianas y microclimáticas del suelo (Castro, Zamora, & Hódar, 2006; Ramírez-Contreras & Rodríguez-Trejo, 2009), mejorando el desempeño de las plantas introducidas bajo el dosel (Avendaño-Yáñez, Sánchez-Velázquez, Meave, & Pineda-López, 2014; Bonfil et al., 2000; Camacho-Cruz, González-Espinosa, Wolf, & de Jong, 2000; Castro et al., 2006).
Crecimiento de Quercus
Las TRC y la biomasa difirieron significativamente (P < 0.001) entre especies y entre las coberturas del dosel (Cuadro 2), lo cual sugiere un efecto diferenciado de las situaciones microambientales de cada condición.
Cuadro 2 Análisis de varianza de la tasa relativa de crecimiento (TRC) de altura y diámetro y de la biomasa acumulada de tres especies de Quercus (Q. ocoteifolia, Q. crispipilis y Q. segoviensis) establecidas en tres sitios con diferente condición microambiental (bosque, matorral y pastizal) en San Cristóbal de Las Casas, Chiapas.
| Factor | Grados de libertad | TRC altura | TRC diámetro | Biomasa | |||
|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | ||
| Especie | 2 | 15.52 | <0.001 | 13.82 | <0.001 | 19.22 | <0.001 |
| Sitio | 2 | 11.09 | <0.001 | 71.53 | < 0.001 | 17.85 | <0.001 |
| Especie*Sitio | 4 | 1.59 | 0.17 | 4.45 | < 0.01 | 1.88 | 0.11 |
La Figura 4 presenta las TRC en altura y diámetro de las tres especies de Quercus por cobertura. Las TRC en altura de Q. crispipilis y Q. ocoteifolia fueron mayores en condiciones con radiación y temperatura intermedia y baja (matorral y bosque). Quercus segoviensis presentó el incremento más bajo de TRC en altura y no mostró diferencias (P < 0.05) entre sitios. Con respecto a la TRC en diámetro, las tres especies incrementaron en el sitio con mayor radiación (pastizal) seguido por el matorral y el bosque. Este patrón se repitió en la acumulación de biomasa en Q. crispipilis y Q. segoviensis, principalmente en las raíces (Figura 5).
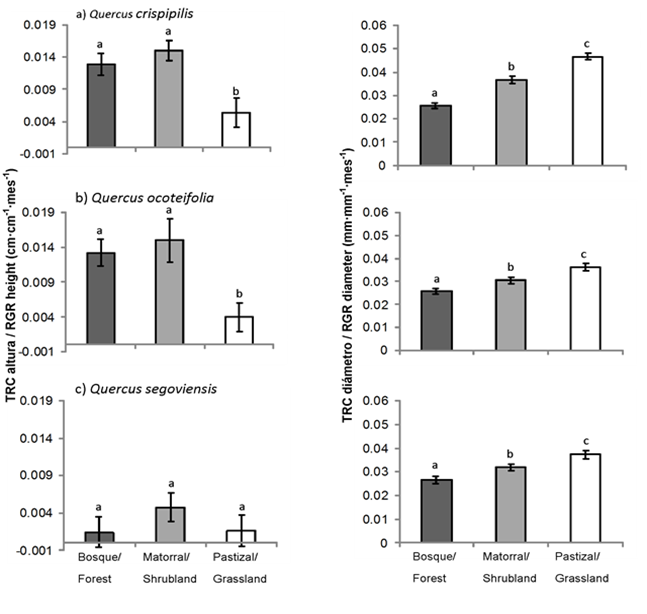
Figura 4 Tasa relativa de crecimiento (TRC) en altura y diámetro de especies de Quercus después de 14 meses de crecimiento en campo en San Cristóbal de Las Casas, Chiapas. Letras diferentes indican diferencias significativas entre sitios mediante la prueba de Tukey (P < 0.05). ± error estándar de la media.
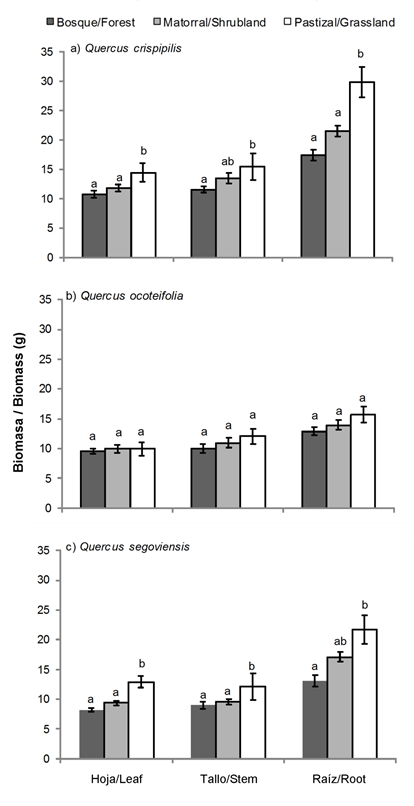
Figura 5 Biomasa acumulada de tres especies de Quercus plantadas en tres condiciones de cobertura (bosque, matorral y pastizal) en San Cristóbal de Las Casas, Chiapas. Letras diferentes indican diferencias significativas entre sitios, acorde con la prueba de Tukey (P < 0.05). ± error estándar de la media (n = 45).
Estos resultados sugieren una asociación directa entre el incremento en el diámetro basal y el incremento en la biomasa radical en Q. crispipilis y Q. segoviensis, además de una relación inversa entre las TRC en altura y diámetro basal de Q. crispipilis y Q. ocoteifolia. Los mayores incrementos en altura -en contraste con el diámetro- se asociaron con niveles bajos e intermedios de radiación solar que variaron de 5 a 50 % (Cardillo & Bernal 2006; Neufeld, 1983) y bajas fluctuaciones de temperatura y humedad bajo el dosel del bosque (Padilla & Pugnaire, 2006; Ramírez-Contreras & Rodríguez-Trejo, 2009). Aunque se considera que la luz es un recurso necesario para el crecimiento, el incremento de radiación no significa necesariamente incremento en el tamaño de las plantas (Espelta et al., 1995). Los resultados indican que, en los encinos, el crecimiento en diámetro basal -asociado con el incremento en la biomasa radical- es más común que el crecimiento en altura, poco después del trasplante, lo que coincide con lo observado por Bonfil y Soberón (1999) y Kabeya y Sakai (2003).
El incremento en la biomasa de cada componente (hojas, tallo y raíz) de Q. crispipilis y Q. segoviensis fue mayor en el pastizal que en el matorral y el bosque (Figura 5). El mayor incremento de biomasa en plantas establecidas en sitios con mayor radiación solar puede atribuirse a que éstas sufren más estrés, el crecimiento en el tallo se vuelve más lento y los carbohidratos se distribuyen hacia las partes inferiores (Dickson, 1990); además, en estas condiciones, las plantas alcanzan mayores tasas fotosintéticas (Feltrin et al., 2016; Kabeya & Sakay, 2003; Ramírez-Contreras & Rodríguez-Trejo, 2009). El componente que alcanzó mayor biomasa fue la raíz; la importancia de este órgano radica en la capacidad de almacenar la mayor parte de los carbohidratos que la planta utilizará durante los periodos de condiciones adversas (Arosa et al., 2015; Kabeya & Sakay, 2003). Las reservas almacenadas en las raíces permiten a los encinos rebrotar en caso de muerte o remoción parcial de la parte aérea (Bonfil & Soberón 1999; Cardillo & Bernal, 2006; Vázquez de Castro, Oliet, Puértolas, & Jacobs, 2014).
Las tres especies de Quercus presentaron valores altos de supervivencia en las tres situaciones experimentales; aunque las variables microclimáticas son distintas entre las condiciones estudiadas, no se puede afirmar que sean los únicos factores que afectan la supervivencia, ya que también pueden influir la herbivoría y la fertilidad de los suelos (Lei et al., 2013). En razón de que Q. crispipilis y Q. segoviensis presentaron mayores incrementos de biomasa en el sitio con mayor iluminación, se recomienda el uso de estas especies en la restauración de pastizales abandonados; mientras que Q. ocoteifolia tendría mayor éxito en la restauración de bosques secundarios degradados, ya que el desempeño mejora a la sombra del dosel forestal.
Conclusiones
La presencia de dosel influye en las variables microclimáticas de los sitios; a menor cobertura en el sitio se recibe mayor radiación y temperatura ambiental y del suelo. A pesar de lo anterior, la supervivencia de Q. crispipilis y Q. segoviensis no fue afectada por la cobertura del dosel. Tal como se esperaba, las tasas relativas de crecimiento de altura y de diámetro cambiaron en función de la cobertura del dosel. La altura de Q. crispipilis y Q. ocoteifolia fue mayor en las condiciones bosque y matorral; el diámetro de las tres especies y la biomasa de cada componente en Q. crispipilis y Q. segoviensis fueron mayores en el pastizal, intermedios en el matorral e inferiores en el bosque. Las tres especies parecen adaptarse a las diferentes condiciones de radiación, temperatura y humedad características de cada sitio. Quercus crispipilis y Q. segoviensis tienen mejores respuestas de supervivencia y crecimiento en el matorral y pastizal, mientras que Q. ocoteifolia resulta favorecida por la presencia de dosel forestal.
Agradecimientos
Al Consejo Nacional de Ciencia y Tecnología por la beca otorgada al primer autor para realizar sus estudios de maestría (No. 574853). Al Sr. Noé Beltrán, por las facilidades y el acceso a los sitios de estudio.
REFERENCIAS
Alba-López, M. P., González-Espinosa, M., Ramírez-Marcial N. & Castillo-Santiago, M. Á. (2003). Determinantes de la distribución de Pinus spp. en la Altiplanicie Central de Chiapas, México. Boletín de la Sociedad Botánica de México, 73(2), 7-15. Retrieved from http://www.redalyc.org/pdf/577/57707301.pdf [ Links ]
Alvarez-Aquino, C., & Williams-Linera, G. (2012). Seedling survival and growth of tree species: site condition and seasonality in tropical dry forest restoration. Botanical Sciences, 90(3), 341-351.doi: 10.17129/botsci.395 [ Links ]
Arosa, M. L., Ceia, R. S., Costa, S. R., & Freitas, H. (2015). Factors affecting cork oak (Quercus suber) regeneration: Acorn sowing success and seedling survival under field conditions. Plant Ecology and Diversity, 1(4), 1−12. doi: 10.1080/17550874.2015.1051154 [ Links ]
Avendaño-Yáñez, M. L., Sánchez-Velásquez, L. R., Meave, J. A. & Pineda-López, M. R. (2014). Is facilitation a promising strategy for cloud forest restoration? Forest Ecology & Management, 329(8), 328-333. doi: 10.1016/j.foreco.2014.01.051 [ Links ]
Bonfil, C., & Soberón, J. (1999). Quercus rugosa seedling dynamics in relation to its re-introduction in a disturbed Mexican landscape. Applied Vegetation Science, 2(2), 189-200. doi: 10.2307/1478982 [ Links ]
Bonfil, C., Rodríguez de la Vega, H., & Peña, R. V. (2000). Evaluación del efecto de las plantas nodriza sobre el establecimiento de una plantación de Quercus L. Revista Ciencia Forestal en México, 25(1), 59-73. Retrieved from http://cienciasforestales.inifap.gob.mx/editorial/index.php/Forestales/article/view/18 [ Links ]
Bustamante, R. O., Badano, E. I., & Pickett, S. T. A. (2012). Impacts of land use change on seed removal patterns of native and exotic species in a forest landscape. Community Ecology, 13(2), 171-177. doi: 10.1556/ComEc.13.2012.2.6 [ Links ]
Camacho-Cruz, A., González-Espinosa, M., Wolf, J. H. D., & de Jong, B. H. J. (2000). Germination and survival of tree species in disturbed forests of the highlands of Chiapas, Mexico. Canadian Journal of Botany, 78(10), 1309-1318. doi: 10.1139/b00-103 [ Links ]
Cardillo, E., & Bernal, C. J. (2006). Morphological response and growth of cork oak (Quercus suber L.) seedlings at different shade levels. Forest Ecology and Management, 222(1-3), 296-301. doi: 10.1016/j.foreco.2005.10.026 [ Links ]
Castro, J., Zamora, R., & Hódar, J. A. (2006). Restoring Quercus pyrenaica forests using pioneer shrubs as nurse plants. Applied Vegetation Science, 9(1), 137-142. doi: 10.1658/1402-2001(2006)9[137:RQPFUP]2.0.CO;2 [ Links ]
Cayuela, L., Rey-Benayas, J. M., & Echeverría, C. (2006). Clearance and fragmentation of tropical montane forests in the highlands of Chiapas, Mexico (1975-2000). Forest Ecology and Management, 226(1-3), 208-218. doi: 10.1016/j.foreco.2006.01.047 [ Links ]
Comisión Nacional del Agua (CONAGUA). (2017). Normales climatológicas: Chiapas. Retrieved April 10, 2017 from http://smn1.conagua.gob.mx/index.php?option=com_content&view=article&id=174&tmpl=component [ Links ]
Crawley, M. J. (2013). The R Book (Second edition). West Sussex, United Kingdom: John Wiley & Sons. [ Links ]
De la Mora-Estrada, L. F., Ruiz-Montoya, L., Ramírez-Marcial, N., Morón-Ríos, A., & Mayorga-Martínez, M. C. (2017). Diversidad de chinches (Hemiptera: Heteroptera) en bosques secundarios de pino-encino de San Cristóbal de Las Casas, Chiapas, México. Revista Mexicana de Biodiversidad, 88(1), 86−105. doi: 10.1016/j.rmb.2017.01.016 [ Links ]
Dickson, R. E. (1990). Assimilate distribution and storage. En A. S. Raghavendra (Ed.), Physiology of trees (pp. 51-85). New York, USA: Wiley and Sons. [ Links ]
Espelta, J. M., Riba, M., & Retana, J. (1995). Patterns of seedling recruitment in West-Mediterranean Quercus ilex forests influenced by canopy development. Journal of Vegetation Science, 6(4), 465-472. doi: 10.2307/3236344 [ Links ]
Feltrin, R. P., Will, R. E., Meek, C. R., Masters, R. E., Waymire, J., & Wilson, D. S. (2016). Relationship between photosynthetically active radiation and understory productivity across a forest-savanna continuum. Forest Ecology and Management, 374(1), 51-60. doi: 10.1016/j.foreco.2016.04.049 [ Links ]
Flores-Cano, J., Badano, E. I., & Flores, J. (2012). Effects of burial depth on seed germination and seedling emergence of Mexican oaks: A glasshouse experiment. Archives of Biological Sciences, 64(2), 1543-1554. doi: 10.2298/ABS1204543C [ Links ]
Galindo-Jaimes, L., González-Espinosa, M., Quintana-Ascencio, P., & García-Barrios, L. (2002). Tree composition and structure in disturbed stands with varying dominance by Pinus spp. in the highlands of Chiapas, México. Plant Ecology, 162(2), 259-272. doi: 10.1023/A:1020309004233 [ Links ]
González-Espinosa, M., Meave, J. A., Lorea-Hernández, F. G., Ibarra-Manríquez, G., & Newton, A. C. (2011). The red list of Mexican cloud forest trees. Cambridge: Fauna & Flora International. [ Links ]
González-Espinosa, M., Meave, J. A., Ramírez-Marcial, N., Toledo-Aceves, T., Lorea-Hernández, F. G., & Ibarra-Manríquez, G. (2012). Los bosques de niebla de México: conservación y restauración de su componente arbóreo. Ecosistemas, 21(1), 36-52. Retrieved from http://www.revistaecosistemas.net/index.php/ecosistemas/article/viewFile/26/20 [ Links ]
Guo, H., Wang, X. A., Zhu, Z. H., Wang, S. X., & Guo, J. C. (2011). Seed and microsite limitation for seedling recruitment of Quercus wutaishanica on Mt. Ziwuling, Loess Plateau, China. New Forests, 41(1), 127-137. [ Links ]
Gutiérrez, E., & Trejo, I. (2014). Efecto del cambio climático en la distribución potencial de cinco especies arbóreas de bosque templado en México. Revista Mexicana de Biodiversidad, 85(1), 179-188. doi: 10.7550/rmb.37737 [ Links ]
Hunt, R., Causton, D. R., Shipley, B., & Askew, A. P. (2002). A modern tool for classical plant analysis. Annals of Botany, 90(4), 485-488. doi: 10.1093/aob/mcf214 [ Links ]
Kabeya, D., & Sakai, S. (2003). The role of roots and cotyledons as storage organs in early stages of establishment in Quercus crispula: A quantitative analysis of the nonstructural carbohydrate in cotyledons and roots. Annals of Botany, 92(4), 537-45. doi: 10.1093/aob/mcg165 [ Links ]
Lamb, D., Stanturf, J., & Madsen, P. (2012). What is forest landscape restoration? En J. Stanturf, D. Lamb, & M. Palle (Eds.), Forest landscape restoration (pp 3-24). Athens, Georgia, USA: Springer. [ Links ]
Lei, J. P., Xiao, W., Liu, J. F., Xiong, D., Wang, P., Pan L., Jiang Y., & Li, M. H. (2013). Responses of nutrients and mobile carbohydrates in Quercus variabilis seedlings to environmental variations using in situ and ex situ experiments. PLOS ONE, 8(4), e61192. doi: 10.1371/journal.pone.0061192 [ Links ]
Neufeld, H. S. (1983). Effects of light on growth, morphology, and photosynthesis in Bald cypress (Taxodium distichum (L.) Rich.) and Pond cypress (T. ascendens Brongn.) seedlings. Bulletin of the Torrey Botanical Club, 110(1), 43-54. Retrieved from http://www.appstate.edu/~neufeldhs/publications/neuf1983.pdf [ Links ]
Padilla, F. M., & Pugnaire, F. I. (2006). The role of nurse plants in the restoration of degraded environments. Frontiers in Ecology and the Environment, 4(4), 196-202. doi: 10.1890/1540-9295(2006)004[0196:TRONPI]2.0.CO;2 [ Links ]
Pulsford, S. A., Lindenmayer, D. B., & Driscoll, D. A. (2016). A succession of theories: Purging redundancy from disturbance theory. Biological Reviews, 91(1), 148-167. doi: 10.1111/brv.12163 [ Links ]
R Development Core Team. (2015). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Retrieved from http://www.R-project.org [ Links ]
Ramírez-Contreras, A., & Rodríguez-Trejo, D. A. (2004). Effect of seedling quality, aspect and microsite on a Quercus rugosa plantation. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 10(1), 5-11. Retrieved from https://chapingo.mx/revistas/forestales/contenido.php?id_articulo=403&doi=&id_revista=3 [ Links ]
Ramírez-Contreras, A., & Rodríguez-Trejo, D. A. (2009). Nurse plants in the reforestation with Pinus hartwegii Lindl. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 15(1), 43-48. Retrieved from https://chapingo.mx/revistas/forestales/contenido.php?id_articulo=511&doi=&id_revista=3 [ Links ]
Ramírez-Marcial, N., Camacho-Cruz, A., & González-Espinosa, M. (2008). Clasificación de grupos funcionales vegetales para la restauración del bosque mesófilo de montaña. En L. R. Sánchez-Velásquez, J. Galindo-González, & F. Díaz-Fleischer (Eds.), Ecología, manejo y conservación de los ecosistemas de montaña en México (pp. 51-72). México: Mundi Prensa. [ Links ]
Ramírez-Marcial, N., Camacho-Cruz, A., González-Espinosa, M., & López-Barrera, F. (2006). Establishment, survival and growth of tree seedlings under successional montane oak forests in Chiapas, Mexico. In M. Kappelle (Ed.), Ecology and conservation of neotropical montane oak forests (pp. 177-189). Berlin: Springer. doi: 10.1007/3-540-28909-7_14 [ Links ]
Ramírez-Marcial, N., Camacho-Cruz, A., Martínez-Icó, M., Luna-Gómez, A., Golicher, D., & González-Espinosa, M. (2010). Árboles y arbustos de los bosques de montaña en Chiapas. San Cristóbal de Las Casas, Chiapas, México: El Colegio de la Frontera Sur. [ Links ]
Ramírez-Marcial, N., González-Espinosa, M., García-Moya, E. (1996). Establecimiento de Pinus spp. y Quercus spp. en matorrales y pastizales de Los Altos de Chiapas, México. Agrociencia, 30(2), 249-257. [ Links ]
Ramos-Palacios, C. R., & Badano, E. I. (2014). The relevance of burial to evade acorn predation in an oak forest affected by habitat loss and landscape use changes. Botanical Sciences, 92(2), 299-308. doi: 10.17129/botsci.101 [ Links ]
Rich, P. M., Wood, J., Vieglais, D. A., Burek, K., & Webb, N. (1999). HemiView user manual. Retrieved from ftp://ftp.dynamax.com/manuals/HemiView_Manual.pdf [ Links ]
Sánchez-Velásquez, L. R., Domínguez-Hernández, D., Pineda-López, M. R., & Lara-González, R. (2011). Does Baccharis conferta shrub act as a nurse plant to the Abies religiosa seedling? The Open Forest Science Journal, 4(1), 67-70. doi: 10.2174/1874398601104010067 [ Links ]
Sánchez-Velásquez, L. R., Ramírez-Bamonde, E. S., Andrade-Torres, A., & Rodríguez-Torres, P. (2008). Ecología florística y restauración del bosque mesófilo de montaña. En L. R. Sánchez-Velásquez, J. Galindo-González, & F. Díaz-Fleischer (Eds.), Ecología, manejo y conservación de los ecosistemas de montaña en México (pp. 9-50). México: Mundi Prensa . [ Links ]
Stanturf, J. A., Palik, B. J., Williams, M. I., Dumroese, R. K., & Madsen, P. (2014). Forest restoration paradigms. Journal of Sustainable Forestry, 33, S161−S194. doi: 10.1080/10549811.2014.884004 [ Links ]
Torres-Miranda, A., Luna-Vega, I., & Oyama, K. (2011). Conservation biogeography of red oaks (Quercus, Section Lobatae) in Mexico and Central America. American Journal of Botany, 98(2), 290-305. doi: 10.3732/ajb.1000218 [ Links ]
Vázquez de Castro, A., Oliet, J. A., Puértolas, J., & Jacobs, D. F. (2014). Light transmissivity of tube shelters affects root growth and biomass allocation of Quercus ilex L. and Pinus halepensis Mill. Annals of Forest Science, 71(1), 91-99. doi: 10.1007/s13595-013-0335-3 [ Links ]
Recibido: 02 de Enero de 2017; Aprobado: 12 de Abril de 2017
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
