










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.23 no.2 Chapingo may./ago. 2017
https://doi.org/10.5154/r.rchscfa.2016.10.055
Artículos científicos
Ecotono del límite superior del bosque de Pinus hartwegii Lindl.: estructura y límites altitudinales en el Nevado de Toluca, México
1Universidad Autónoma del Estado de México, Instituto de Ciencias Agropecuarias y Rurales (ICAR). Carretera Toluca-Ixtlahuaca km 15.5, El Cerrillo Piedras Blancas. C. P. 50295. Toluca de Lerdo, Estado de México, México.
2Instituto Potosino de Investigación Científica y Tecnológica, División de Ciencias Ambientales. Camino a la Presa de San José núm. 2055 Lomas 4a. C. P. 78216. San Luís Potosí, San Luís Potosí, México.
Introducción:
La complejidad del ecotono del límite superior del bosque de Pinus hartwegii puede funcionar como una barrera para la migración altitudinal.
Objetivos:
La estructura del bosque de P. hartwegii se caracterizó a través del ecotono de su límite superior en el Nevado de Toluca; además, se determinaron los posibles cambios en los límites altitudinales en los últimos 25 años.
Materiales y métodos:
Se determinaron la densidad arbórea, altura y diámetro normal de cada árbol, y la regeneración natural del bosque. Los límites altitudinales se determinaron mediante el método de la ventana móvil en imágenes de satélite de 1989 y 2014.
Resultados y discusión:
La densidad del arbolado se redujo entre 8 y 70 % en el gradiente altitudinal. Los límites altitudinales (3,980-4,130 m) no presentaron diferencias significativas (P = 0.07) entre los años evaluados. El ecotono es de forma difusa, regulado por limitaciones al crecimiento. Esto implica que el bosque modificará su distribución altitudinal en función de los cambios ambientales, de acuerdo con su amplitud ecológica y la rapidez con que puedan establecerse nuevos individuos a mayores altitudes.
Conclusión:
Es importante establecer monitoreos, a largo plazo, para saber si la especie es realmente capaz de migrar altitudinalmente como resultado del incremento de temperatura ambiental.
Palabras clave: Alta montaña; clases diamétricas; dendrometría; gradiente altitudinal; parches de bosque; zona de transición
Introduction:
Treeline ecotone complexity might function as a barrier to altitudinal migration of Pinus hartwegii Lindl.
Objectives:
P. hartwegii forest structure was characterized through its treeline ecotone at Nevado de Toluca; in addition, possible changes in altitudinal limits over the last 25 years were determined.
Materials and methods:
Height and diameter at breast height (DBH) of each tree were determined, as well as natural forest regeneration and tree density. Altitudinal limits were determined on satellite images from 1989 and 2014 using the moving split window method.
Results and discussion:
Tree density fell between 8 and 70 % across altitudinal gradient. Altitudinal limits (3,980-4,130 m) did not show significant differences (P = 0.07) between two evaluated years. The P. hartwegii ecotone displays a diffuse form, regulated by growth limitations. This implies that the forest might modify its altitudinal distribution based on environmental changes, according to its ecological amplitude and how fast new individuals can become established at higher altitudes.
Conclusion:
Long-term monitoring is important to know if P. hartwegii is actually capable of migrating altitudinally as a result of increased environmental temperature.
Keywords: High mountain; diameter classes; dendrometry; altitudinal gradient; forest patches; transition zone
Introducción
Los ecotonos del límite superior del bosque (LSB) delimitan el punto más alto en el cual puede encontrarse un estrato arbóreo (Körner, 1998; Lutz, Larson, Freund, Swanson, & Bible, 2013); son zonas conspicuas de transición entre ecosistemas boscosos y turberas, páramos de altura o pastizales alpinos (Camarero & Gutiérrez, 2004; Körner, 1998). Debido a que las bajas temperaturas extremas son el principal factor regulador de los procesos fisiológicos y ecológicos de los árboles en altitudes elevadas (Tranquillini, 1964), estos ecotonos se consideran altamente sensibles al cambio climático (Holtmeier & Broll, 2005; Körner, 1998). Bajo este contexto, la presencia de árboles de diversas especies (e. g., Larix sibirica Ledeb., Pinus uncinata Raymond ex A. DC., P. hartwegii Lindl.) por arriba de sus límites altitudinales (enclaves) se asocia a la necesidad de buscar condiciones ambientales semejantes a su nicho original (Camarero & Gutiérrez, 2004; Holtmeier & Broll, 2005; Jobbágy & Jackson, 2000). Por tanto, la migración altitudinal, además de la adaptación y extinción, es una de las posibles respuestas de la vegetación alpina al incremento de temperatura ambiental (Aitken, Yeaman, Holliday, Wang, & Curtis-McLane, 2008). La respuesta estará en función de la especie y de la magnitud en que su distribución esté controlada por la temperatura (Jobbágy & Jackson, 2000). No obstante, si el ecotono no es capaz de migrar a la misma velocidad que la especie en forma individual, la tasa de movimiento disminuirá e incluso la migración se inhibirá en su totalidad, por lo que la complejidad del ecotono podría funcionar como una barrera para la migración altitudinal (Lutz et al., 2013).
En general, el ecotono del LSB presenta condiciones de estrés adicionales a las que se presentan en el núcleo del bosque (Viveros-Viveros et al., 2009). Las bajas temperaturas reducen la actividad de los meristemos apicales y la formación de tejido nuevo (Tranquillini, 1964), generando árboles con deformación estructural o krummholz (Camarero & Gutiérrez, 2004; Körner, 1998) o reduciendo la producción de semillas, la germinación y la supervivencia de plántulas (Coop & Givnish, 2008), lo cual modifica la estructura del ecotono del LSB. Al respecto, con base en el mecanismo regulador dominante en el ecotono, Harsch y Bader (2011) clasifican a los ecotonos del LSB en cuatro tipos: difusos, abruptos, islas y krummholz. El tipo difuso, el más común, se caracteriza por las limitaciones al crecimiento (Harsch & Bader, 2011); los ecotonos abruptos se forman debido al incremento en la mortalidad de plántulas; mientras que la muerte en retroceso produce islas o grupos de árboles aislados entre sí (Bader, Rietkerk, & Bregt, 2007; Harsch, Hulme, McGlone, & Duncan, 2009). La forma de los árboles también puede verse alterada debido a incidencia de vientos intensos o la presencia de nieve sobre los árboles, produciendo krummholz (Körner, 1998). Por lo anterior, la altura y densidad de individuos son atributos clave para entender la estructura y procesos que regulan el establecimiento en el LSB (Bader et al., 2007), y los factores locales que podrían limitar la migración altitudinal de los bosques, bajo el contexto del cambio climático (Harsch & Bader, 2011).
Pinus hartwegii es la especie de pino que se distribuye a mayor altitud (2,800 a 4,200 m) en el mundo (Perry, 1991). Esta especie forma bosques monoespecíficos que forman ecotonos con el pastizal de alta montaña (Challenger & Soberón, 2008); presenta gran adaptación a temperaturas bajas extremas hasta de -30 °C (Perry, 1991), soportando la caída de nieve durante los meses de diciembre a febrero (Challenger & Soberón, 2008). No obstante, la adaptación de la especie a las bajas temperaturas la expone como una de las más vulnerables al cambio climático (Ricker, Gutierrez-García, & Daly 2007).
En México, se han reportado cambios en la distribución altitudinal de P. hartwegii en el Parque Nacional Izta-Popo (Beaman, 1962), Nevado de Toluca (Lauer, 1973) y Cofre de Perote (Iglesias & Tivo, 2006), asociados a cambios en la temperatura local. Sin embargo, tales reportes están enfocados en P. hartwegii como especie, sin considerar la estructura, composición y dinámica del ecotono en conjunto (Harsch & Bader, 2011). Con base en lo anterior, es importante conocer la estructura de P. hartwegii a través de la zona de transición hacia el pastizal alpino desarrollado a mayor altitud y bajo condiciones ambientales más adversas. Por lo tanto, los objetivos del presente trabajo fueron caracterizar la estructura del ecotono del LSB de P. hartwegii en el Nevado de Toluca y determinar si ocurrió algún cambio en los límites altitudinales durante los últimos 25 años. Esto, bajo la expectativa de que el estudio pueda servir como línea base para monitorear el efecto potencial del cambio climático sobre la distribución altitudinal de P. hartwegii.
Materiales y métodos
Área de estudio
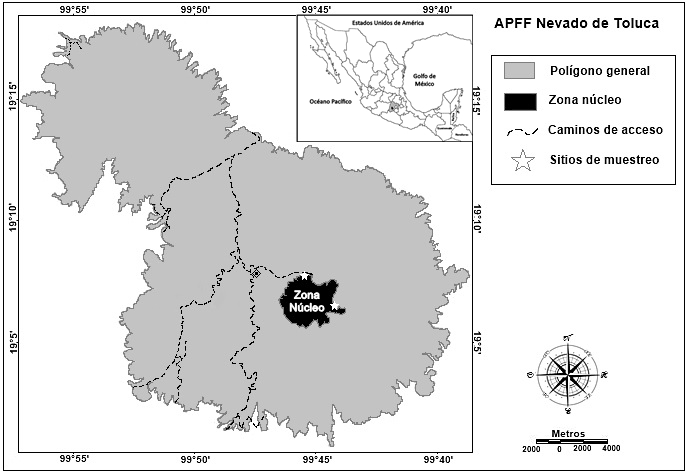
El trabajo se realizó en el Área de Protección de Flora y Fauna (APFF) Nevado de Toluca en el Estado de México, entre los valles de Toluca y Tenango (Figura 1). El área de estudio se localiza entre los 18° 51' - 19° 19' LN y 99° 38' - 100° 09' LO, con un intervalo altitudinal entre 3,000 y 4,680 m (Körner & Paulsen, 2004). Predominan los climas semifrío-subhúmedo C(E)wig y frío E(T)Hwig (García, 1990); la temperatura media anual oscila entre los -2 y 7 °C; y la precipitación media anual, entre 200 y 1,800 mm con caída de nieve en los meses de diciembre a febrero (Challenger & Soberón, 2008). El grupo de suelo dominante es Andosol, presentándose en menor proporción los grupos Feozem, Regosol, Cambisol y Litosol (Körner & Paulsen, 2004). Los principales ecosistemas son el bosque templado de especies de los géneros Abies, Pinus y Quercus, situados a una altitud de 3,000 a 4,100 m; y el pastizal de alta montaña compuesto por los géneros Festuca y Calamagrostis, en altitudes de 4,100 a 4,500 m (Calderón de Rzedowski & Rzedowski, 2005).
Especie en estudio
Pinus hartwegii es un pino adaptado a temperaturas bajas extremas que se distribuye en las cimas de las montañas más altas, desde Nuevo León en el noreste de México hasta El Salvador (Calderón de Rzedowski & Rzedowski, 2005; Perry, 1991), en altitudes de 3,000 a 4,200 m (Beaman, 1962; Lauer, 1973; Perry, 1991). La distribución natural es discontinua y está restringida a sitios de gran altitud como el Nevado de Colima, el Pico de Tancítaro, Nevado de Toluca, Ajusco, Popocatépetl, Iztaccíhuatl, Malinche y Pico de Orizaba, en la Faja Volcánica Transmexicana, donde constituye el límite altitudinal de la vegetación arbórea (Calderón de Rzedowski & Rzedowski, 2005; Vera-Vilchis & Rodríguez-Trejo, 2007). La especie es de gran importancia ecológica debido a su distribución geográfica limitada y su tolerancia a bajas temperaturas, las cuales pueden alcanzar hasta los -30 °C (Beaman, 1962; Perry, 1991).
Diseño de muestreo
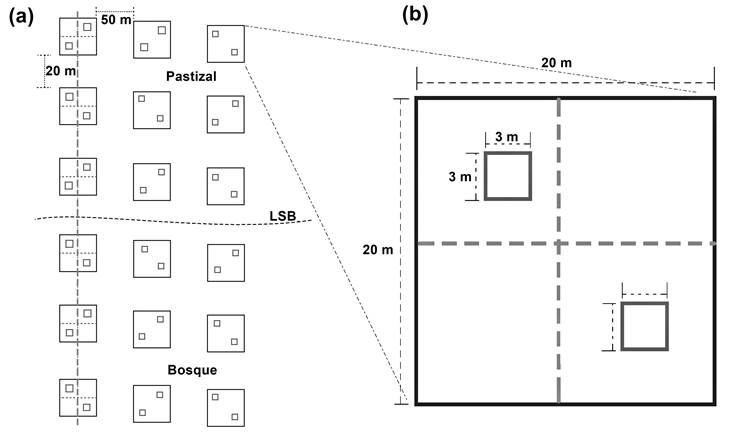
El muestreo fue de tipo estratificado-sistemático (Matteucci & Colma, 1982). La línea del LSB se tomó como punto de partida y a partir de ella se establecieron cinco transectos perpendiculares de 220 m de longitud. Dos transectos se ubicaron en dirección noroeste y tres en dirección este-sureste con respecto al cráter volcánico. En el establecimiento de los transectos se consideró la accesibilidad, estado de conservación de la línea del arbolado y similitud de la pendiente. En cada transecto se formaron seis parcelas de 20 m x 20 m con base en el modelo de transectos y parcelas de Camarero y Fortin (2006); tres parcelas se ubicaron por debajo de la línea forestal (área de bosque) y tres por arriba de la línea (área de pastizal), con una separación de 40 m entre el centro de dos parcelas adyacentes, como se ilustra en la Figura 2. Las parcelas ubicadas en un mismo nivel altitudinal de diferente transecto se consideraron como repeticiones en los cálculos de densidad del arbolado; mientras que para los cálculos de altura y diámetro normal (DN a 1.3 m de altura), los árboles se consideraron de manera individual.
Estructura del ecotono del límite superior del bosque de P. hartwegii
La estructura del ecotono del LSB de P. hartwegii se caracterizó en los meses de agosto a octubre de 2014, a través del registro dendrométrico de los árboles (altura y DN de cada individuo) en las parcelas (Cárdenas, Martínez, Bravo, Núñez, & Caballero, 2008) y de la densidad del arbolado en el gradiente altitudinal (3,980 a 4,120 m). La dimensión de los árboles se comparó estableciendo clases diamétricas por nivel altitudinal e identificando los estratos que componen el ecotono. El porcentaje de reducción proporcional de la densidad del arbolado entre los niveles altitudinales se calculó con la finalidad de identificar la forma que presenta el ecotono del LSB del Nevado de Toluca, de acuerdo con la clasificación de Harsch y Bader (2011). La regeneración del bosque se evaluó mediante el conteo y cálculo de la densidad de plántulas e individuos menores de 1.3 m de altura y 10 cm de DN.
Delimitación altitudinal del ecotono del límite superior del bosque de P. hartwegii
Los límites altitudinales, inferior y superior, entre los que se distribuye la zona de transición del bosque de P. hartwegii a pastizal alpino en el Nevado de Toluca se determinaron mediante la adaptación del método de la ventana móvil de Camarero y Fortin (2006). Esto se llevó a cabo tomando como referencia los pixeles de la imagen correspondiente al año 2014, cuya resolución espacial fue ajustada a 15 m mediante fusión con una imagen pancromática del mismo año. Los promedios de densidad del arbolado por pixel y los bordes del ecotono en los pixeles que presentaron diferencias estadísticas significativas se obtuvieron con el programa IDRISI versión Selva (Clarck Labs, 2012). Estos bordes se corroboraron mediante la clasificación supervisada de 30 puntos georreferenciados dentro del ecotono y 30 puntos en cada cobertura que conforma la zona de transición (bosque y pastizal de alta montaña), los cuales se obtuvieron en campo con ayuda de un GPS con altímetro (marca Garmín, modelo V, EUA). Los límites altitudinales se extrapolaron a todo el ecotono en el Nevado de Toluca; con base en la delimitación realizada, se hizo un análisis de cambios altitudinales para determinar las modificaciones en la distribución altitudinal del ecotono del LSB de P. hartwegii en los últimos 25 años. Para ello, se utilizaron dos imágenes Landsat, una de 1989 y otra de 2014, ajustadas a 15 m de resolución, que se compararon a través del módulo Land Change Modeler de IDRISI versión Selva (Clarck Labs, 2012).
Análisis estadístico
La altura y DN de los árboles, y la densidad de individuos se utilizaron como factores principales en un análisis de varianza de una vía (ANOVA), seguido de una prueba de comparación múltiple de Tukey. Por otra parte, con la finalidad de establecer la relación entre la altura y el DN de los árboles a lo largo del gradiente altitudinal, se aplicó una regresión lineal para la siguiente asociación de variables: altura vs. DN, altura vs altitud, DN vs. altitud, altura-DN vs. altitud. Los resultados del ANOVA aplicado a la densidad del arbolado se utilizaron para obtener los puntos en altitud entre los cuales, las diferencias en densidad fueron estadísticamente significativas; dichos puntos se utilizaron como referencia en la extrapolación de los límites altitudinales. Todos los análisis se realizaron en JMP8 (Statistical Analysis System [SAS], 2008) con un nivel de confianza de 95 %.
Resultados y discusión
Estructura del ecotono del límite superior del bosque de P. hartwegii
El ecotono del LSB del bosque de Pinus hartwegii en el Nevado de Toluca se ajusta a la forma difusa de la clasificación propuesta por Harsch y Bader (2011). Dicha forma describe la reducción gradual de la altura y densidad de individuos conforme se alejan del centro del bosque y la altitud incrementa.
Los árboles del ecotono del LSB de P. hartwegii se encontraron en un rango de 0.3 a 22.0 m de altura con una media de 3.88 m (DE = 4.5). Por debajo de la línea forestal (Bos1, Bos2 y Bos3), los árboles tuvieron una altura media de 4.09 m (DE = 4.7); mientras que por arriba de la línea (Pas4, Pas5 y Pas6), la media fue de 2.2 m (DE = 1.4). Los árboles presentes a 4,043 m (Bos1) registraron la mayor y menor altura.
Respecto al DN, los árboles midieron entre 1.0 y 72.0 cm, con una media de 11.82 cm (DE = 11.85). La Figura 3 muestra que los diámetros más grandes se registraron en los árboles de los niveles Bos1 (4,043 m) y Bos2 (4,050 m). Por debajo de la línea forestal (Bos1, Bos2 y Bos3), los árboles midieron entre 1.0 y 72.0 cm de DN, con una media de 12.05 cm (DE = 12.3); mientras que por arriba de la línea (Pas4, Pas5 y Pas6), midieron entre 1.5 y 26.5 cm, con una media de 9.96 cm (DE = 5.63).
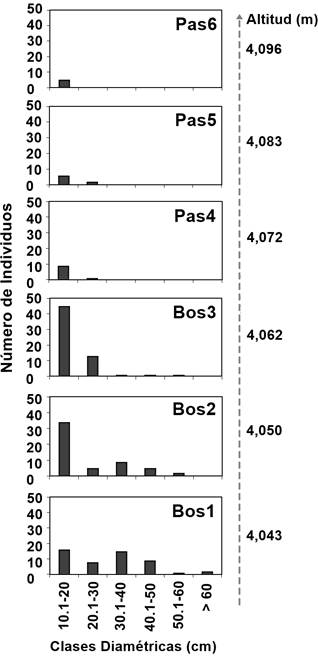
Figura 3 Número de individuos de Pinus hartweggi por clases diamétricas en los niveles altitudinales por arriba (Pas4, Pas5 y Pas6) y por abajo (Bos1, Bos2 y Bos3) de la línea forestal.
La separación por clases diamétricas permitió la identificación del intervalo de altura de los árboles de regeneración (DN ≤ 10 cm), el cual se encontró entre 0.3 y 4.0 m; para individuos de porte juvenil y adulto (DN > 10 cm), la altura varió entre 1.5 y 22 m. Tanto la altura (r2 = 0.144, EE = 4.23) como el DN (r2 = 0.06, EE = 11.39) presentaron relaciones negativas significativas (P = 0.0001) con el incremento en altitud.
De acuerdo con Wong y Lertzman (2001), la reducción de la altura y DN de los árboles a lo largo del gradiente altitudinal, no implica necesariamente una menor edad en los individuos. En altitudes elevadas, la tasa de crecimiento de P. hartwegii se reduce debido a las limitaciones impuestas por las bajas temperaturas (Tranquillini, 1964). En estos sitios pueden encontrarse individuos maduros (62 años) por arriba de los 4,000 m con tallas significativamente menores en comparación con los árboles del núcleo del bosque (Beaman, 1962); la gran adaptación de la especie a temperaturas congelantes, la hace al mismo tiempo vulnerable a los cambios en los rangos de temperatura ambiental, repercutiendo en el crecimiento. Al respecto, Ricker et al. (2007) predicen una reducción de 10.6 % en la tasa de crecimiento de P. hartwegii asociada al incremento de 0.6 °C en la temperatura media ambiental en los próximos 100 años. Aunado a lo anterior, P. hartwegii puede presentar un estado cespitoso durante los primeros años; es decir, su crecimiento es reducido o prácticamente nulo, pudiéndose prolongar hasta siete años (González-Rosales & Rodríguez-Trejo, 2004). Lo anterior dificulta aún más la determinación del momento de establecimiento y la relación que pudiese tener con las variables bióticas y abióticas. Por lo tanto, es imprescindible el desarrollo y la aplicación de técnicas que permitan determinar la edad precisa de individuos en el ecotono del LSB y de enclaves, para dilucidar el efecto del incremento de la temperatura ambiental sobre la distribución altitudinal de la especie en cuestión.
En cuanto a la densidad del arbolado, algunos autores (Holtmeier & Broll, 2005; Jobbágy & Jackson, 2000) señalan que esta variable tiene efectos importantes en la producción y germinación de semillas, así como en el establecimiento y desarrollo de nuevos individuos. De acuerdo con la Figura 4, a lo largo del ecotono del LSB, la densidad del arbolado mostró una reducción gradual de entre 8 y 70 % en los niveles altitudinales. Por otra parte, en la Figura 5 se observa que la mayor densidad se registró en el área de bosque; el nivel altitudinal Bos1 (4,043 m) tuvo el mayor promedio con 37 árboles por parcela (925 árboles·ha-1; DE = 37.3), mientras que el nivel Pas6 (4,096 m) tuvo la menor densidad, siendo de un árbol por parcela (25 árboles·ha-1). Con base en dicha reducción a lo largo del gradiente altitudinal, se puede afirmar que el ecotono del LSB de P. hartwegii corresponde a la forma difusa de la clasificación de Harsch y Bader (2011), siendo relevante que los ecotonos difusos han registrado avances altitudinales con mayor frecuencia a nivel global (Harsch & Bader, 2011; Holtmeier & Broll, 2005; Körner, 1998). Lo anterior confirma que las condiciones ambientales limitan el crecimiento de P. hartwegii sobre los 3,980 m, formando una línea de árboles bien definida (Harsch & Bader, 2011).
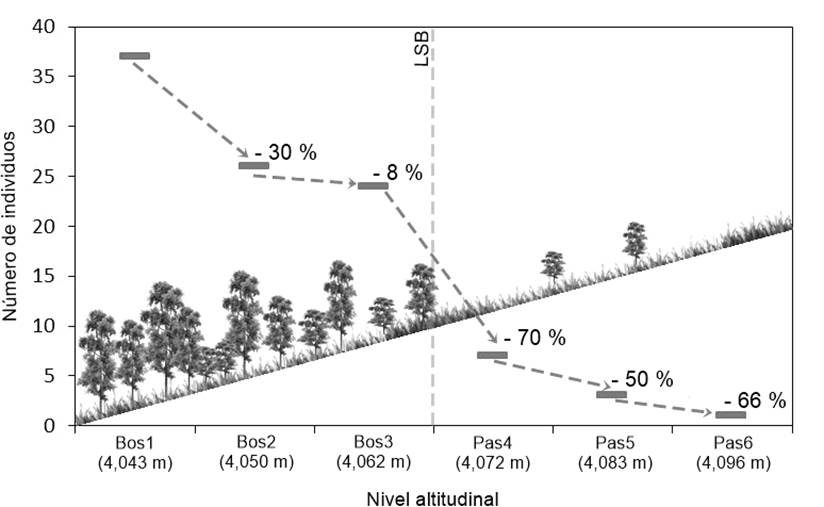
Figura 4 Reducción proporcional de la densidad del arbolado de Pinus hartwegii con respecto al nivel altitudinal inmediato anterior. LSB: límite superior del bosque.
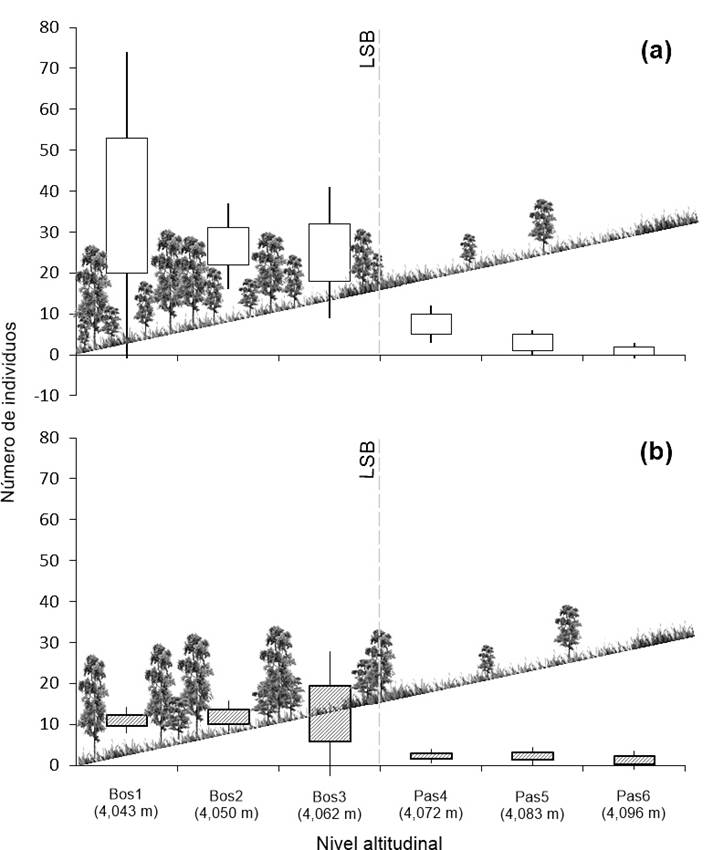
Figura 5 a) Densidad total de árboles en el ecotono del límite superior del bosque (LSB) y b) densidad de árboles mayores de 10 cm de diámetro normal y 1.3 m de altura.
Una baja densidad de individuos de porte adulto (300 a 700 árboles·ha-1) favorece la regeneración arbórea (Camarero & Gutiérrez, 2004), debido a que la presencia de otros árboles mitiga los efectos de las bajas temperaturas extremas e incrementa la disponibilidad de micrositios (Coop & Givnish, 2008). La baja densidad también proporciona espacio suficiente para el establecimiento y desarrollo de nuevos individuos (Viveros-Viveros et al., 2009); en contraste, con una densidad mayor de 1,000 árboles·ha-1, el desarrollo disminuye considerablemente a causa de la reducida capacidad de supervivencia de los individuos de regeneración (Vera-Vilchis & Rodríguez-Trejo, 2007). Esto se refleja en la presencia de enclaves, la cual se considera indicador del avance altitudinal de P. hartwegii; sin embargo, no implica un avance del bosque en sí, más bien representa el éxito de los individuos genéticamente más resistentes, capaces de sobreponerse a las condiciones adversas y que encontraron un micrositio favorable para el establecimiento de plántulas.
Bajo las condiciones de la alta montaña, una mayor densidad puede incluso disminuir la temperatura del suelo bajo los árboles por el sombreo, reduciendo la capacidad de supervivencia y desarrollo del sistema radical y de las plántulas en general. Al respecto, Körner y Paulsen (2004) reportaron que temperaturas inferiores a 5 °C y superiores a 12 °C reducen el éxito de germinación y crecimiento inicial de P. hartwegii en el Nevado de Toluca y Pico de Orizaba. El rango tan estrecho de temperatura óptima sugiere que un cambio mínimo de temperatura ambiental podría impactar significativamente la dinámica poblacional de la especie. Esto pone de manifiesto la necesidad de conocer, de manera más precisa, el efecto de las condiciones ambientales (temperatura y características del suelo) y del gradiente altitudinal sobre los mecanismos que restringen el crecimiento de P. hartwegii en el límite de su distribución altitudinal. Lo anterior con la finalidad de obtener predicciones más puntuales sobre la distribución potencial de P. hartwegii a largo plazo y saber si la especie es realmente capaz de migrar altitudinalmente como resultado del incremento de temperatura ambiental.
Delimitación altitudinal del ecotono del límite superior del bosque de P. hartwegii
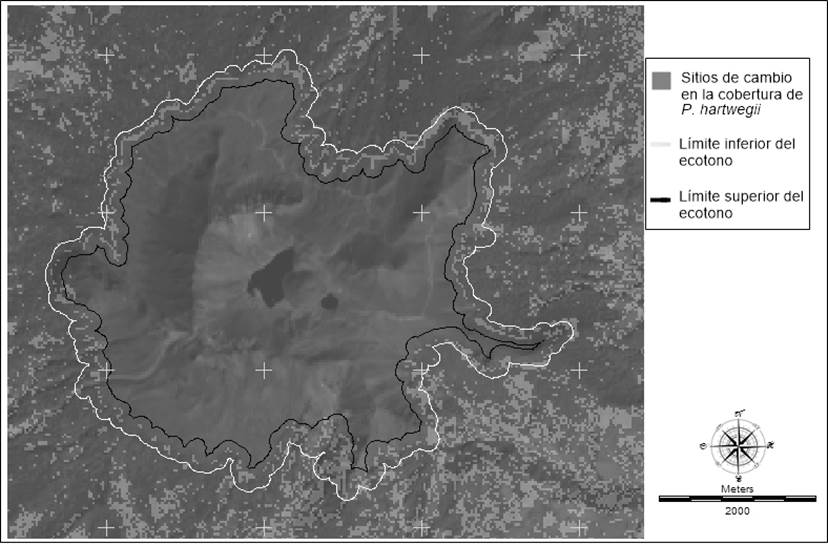
Los límites altitudinales del ecotono del LSB de P. hartwegii identificados en la imagen de 1989 se encontraron en un rango de 3,990 a 4,040 m como límite inferior y entre 4,055 y 4,110 m como límite superior. En las imágenes de 2014, el límite inferior se identificó entre 3,980 y 4,090 m, y el superior entre 4,030 y 4,130 m (Figura 6). Estos rangos son congruentes con los reportados hace 55 años por Beaman (1962) en el Iztaccíhuatl (3,920 a 4,111 m) y Popocatépetl (3,753 a 4,076 m). Dichas similitudes evidencian no solo un patrón de comportamiento de la especie, sino una convergencia de las condiciones en las que se distribuye y el efecto de los cambios históricos que las condiciones ambientales han tenido sobre ella (Jobbágy & Jackson, 2000; Körner & Paulsen, 2004). El límite superior identificado para 2014 fue superior hasta en 20 m en comparación con el de 1989; sin embargo, no se encontraron diferencias significativas (P = 0.07) entre los dos años muestreados. No se encontraron diferencias significativas (P = 0.11) en la altitud del LSB entre las exposiciones noroeste y este-sureste muestreadas en campo; la cara norte del cono volcánico registró la mayor variación altitudinal del ecotono del LSB (Figura 6). El rango de avance de la cara norte fue de entre 25 y 60 m lineales equivalentes a un incremento promedio de 6.5 m en altitud (DE = 4.5) en el borde superior del ecotono del LSB. Por su parte, el lado sur no presentó cambios visibles en la distribución altitudinal del ecotono con las imágenes utilizadas. Es importante considerar que las tasas de crecimiento de formas de vida arbórea, y particularmente, cuando se desarrollan en los límites altitudinales de la alta montaña, son sumamente lentas. Por lo anterior, el monitoreo de las especies y su distribución altitudinal, a largo plazo, tanto del ecotono como de los enclaves situados por arriba de la zona de transición, es fundamental para determinar si existen procesos de regeneración asociados al mantenimiento del ecotono; es decir, si se trata de un ecotono estable o dinámico con avances y retrocesos constantes (Harsch et al., 2009; Harsch & Bader, 2011).
Conclusiones
A nivel global, la forma difusa en ecotonos se considera la más frecuente, siendo dominantes las limitaciones al crecimiento de los árboles. La forma difusa del ecotono del LSB de P. hartwegii refleja las condiciones adversas a las que está sometida. Considerando que este tipo de ecotono puede presentar una dinámica de avance y retroceso altitudinal constante, el presente estudio brinda una línea base para comparaciones posteriores. Dado que no se encontraron diferencias en la distribución altitudinal del ecotono de P. hartwegii entre los periodos evaluados, es importante establecer monitoreos a largo plazo para evidenciar la dinámica de la especie, y saber si es realmente capaz de migrar altitudinalmente como resultado del incremento de temperatura ambiental. Esto permitiría, en su momento, la aplicación de técnicas eficientes de manejo adaptativo.
Agradecimientos
Los autores agradecen al CONACYT por la beca otorgada a Farid U. Alfaro Ramírez para la realización de la Maestría en Ciencias Agropecuarias y Recursos Naturales, y a la CEPANAF por las facilidades otorgadas al interior del APFF Nevado de Toluca para la realización del estudio. El presente trabajo de investigación fue financiado parcialmente por los proyectos UAEMEX3770/2014/CID y CONACYT Ciencia Básica núm. 219696.
REFERENCIAS
Aitken, N. S., Yeaman, S., Holliday, J. A., Wang, T., & Curtis-McLane, S. (2008). Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evolutionary Applications, 1, 95−111. doi: 10.1111/j.1752-4571.2007.00013.x [ Links ]
Bader, M. Y., Rietkerk, M., & Bregt, A. K. (2007). Vegetation structure and temperature regimes of tropical alpine treelines. Arctic, Antarctic, and Alpine Research, 39, 353−364. doi: 10.1657/1523-0430(06-055)[BADER]2.0.CO;2 [ Links ]
Beaman, J. H. (1962). The timberlines of Iztaccíhuatl and Popocatepetl, Mexico. Ecology, 43, 377−385. doi: 10.2307/1933367 [ Links ]
Calderón de Rzedowski, G. & Rzedowski, J. (2005). Flora fanerogámica del Valle de México. Michoacán, México: INECOL-CONABIO. [ Links ]
Camarero, J. J., & Fortin, M. J. (2006). Detección cuantitativa de fronteras ecológicas y ecotonos. Ecosistemas, 15, 77−88. Retrieved from http://www.revistaecosistemas.net/index.php/ecosistemas/article/view/164 [ Links ]
Camarero, J. J., & Gutiérrez, E. (2004). Pace and pattern of recent treeline dynamics: Response of ecotones to climatic variability in the Spanish Pyrenees. Climatic Change, 63, 181−200. doi: 10.1023/B:CLIM.0000018507.71343.46 [ Links ]
Cárdenas, D. I., Martínez, V. M. M., Bravo, I. J. A., Núñez, B. A., & Caballero, G. R. (2008). Manejemos el bosque. La Habana, Cuba: ACTAF. [ Links ]
Challenger, A., & Soberón, J. (2008). Los ecosistemas terrestres. En Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO) (Eds.), Capital natural de México, vol. I: Conocimiento actual de la biodiversidad (pp.87−108). México: CONABIO. [ Links ]
Clarck Labs. (2012). IDRISI Selva. Worcester, MA, USA: Clark University. [ Links ]
Coop, J. D., & Givnish, T. J. (2008). Constraints on tree seedling establishment in montane grasslands of the Valles Caldera, New Mexico. Ecology, 89, 1101−1111. doi: 10.1890/06-1333.1 [ Links ]
García, E. (1990). Carta de climas, Atlas Nacional de México. México: Universidad Nacional Autónoma de México (UNAM). [ Links ]
González-Rosales, A., & Rodríguez-Trejo, D. A. (2004). Efecto del chamuscado de copa en el crecimiento en diámetro de Pinus hartwegii Lindl. en el Distrito Federal, México. Agrociencia, 38, 537−544. Retrieved from http://www.fire.uni-freiburg.de/GlobalNetworks/MesoAmerica/Research/Chamuscado-Dante.pdf [ Links ]
Harsch, M. A., & Bader, M. Y. (2011). Treeline form - a potential key to understanding treeline dynamics. Global Ecology and Biogeography, 20, 582−596. doi: 10.1111/j.1466-8238.2010.00622.x [ Links ]
Harsch, M. A., Hulme, P. E., McGlone, M. S., & Duncan, R. P. (2009). Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecology Letters, 12, 1040−1049. doi: 10.1111/j.1461-0248.2009.01355.x [ Links ]
Holtmeier, F. K., & Broll, G. (2005). Sensitivity and response of northern hemisphere altitudinal and polar treelines to environmental change at landscape and local scales. Global Ecology and Biogeography, 14, 395−410. doi: 10.1111/j.1466-822x.2005.00168.x [ Links ]
Iglesias, A. L. G., & Tivo, F. Y. (2006). Caracterización morfométrica de la población de Pinus hartwegii Lindl. del Cofre de Perote, Veracruz, México. Ra Ximhai, 2, 449−468. Retrieved from http://www.revistas.unam.mx/index.php/rxm/article/view/6883 [ Links ]
Jobbágy, E. G., & Jackson, R. B. (2000). Global controls of forest line elevation in the northern and southern hemispheres. Global Ecology and Biogeography, 9, 253-268. doi: 10.1046/j.1365-2699.2000.00162.x [ Links ]
Körner, C. (1998). A re-assessment of high elevation treeline positions and their explanation. Oecologia, 115, 445−459. doi: 10.1007/s004420050540 [ Links ]
Körner, C., & Paulsen, J. (2004). A world-wide study of high altitude treeline temperatures. Journal of Biogeography, 31, 713−732. doi: 10.1111/j.1365-2699.2003.01043.x [ Links ]
Lauer, W. (1973). The altitudinal belts of the vegetation in the central Mexican highlands and their climatic conditions. Arctic and Alpine Research, 5, A99−A113. Retrieved from http://www.jstor.org/stable/1550159 [ Links ]
Lutz, J. A., Larson, A. J., Freund, J. A., Swanson, M. E. & Bible, K. J. (2013). The importance of large-diameter trees to forest structural heterogeneity. PLoS ONE, 812, e82784. doi: 10.1371/journal.pone.0082784 [ Links ]
Matteucci, S. D., & Colma, A. (1982). Metodología para el estudio de la vegetación. Washington, DC, USA: Secretaría General de la Organización de los Estados Americanos-Programa Regional de Desarrollo Científico y Tecnológico. Retrieved from https://aprobioma.files.wordpress.com/2011/03/metod_para_el_estudio_de_la_vegetacion_archivo1.pdf [ Links ]
Perry, J. P. Jr. (1991). The pines of Mexico and Central America. Portland, USA: Timber Press. [ Links ]
Ricker, M., Gutiérrez-García, G., & Daly, D. C. (2007). Modeling long-term tree growth curves in response to warming climate: Test cases from a subtropical mountain forest and a tropical rainforest in México. Canadian Journal of Forest Research, 37, 977−989. doi: 10.1139/X06-304 [ Links ]
Statistical Analysis System (SAS Institute Inc.). (2008). JMP 8.0, Statistical discovery from SAS. Cary, NC, USA: Author. [ Links ]
Tranquillini, W. (1964). The physiology of plants at high altitudes. Plant Physiology, 15, 345−362. doi: 10.1146/annurev.pp.15.060164.002021 [ Links ]
Vera-Vilchis, V., & Rodríguez-Trejo, D. A. (2007). Survival and height increment of Pinus hartwegii two years after prescribed burns and experimental forest fires. Agrociencia, 41, 219−230. Retrieved from http://www.colpos.mx/agrocien/Bimestral/2007/feb-mar/art-9.pdf [ Links ]
Viveros-Viveros, H., Sáenz-Romero, C., Vargas-Hernández, J. J., López-Upton, J., Ramírez-Valverde, G., & Santacruz-Varela, A. (2009). Altitudinal genetic variation in Pinus hartwegii Lindl.: Height growth, shoot phenology and cold damage in seedlings. Forest Ecology and Management, 257, 836−842. doi: 10.1016/j.foreco.2008.10.021 [ Links ]
Wong, C. M., & Lertzman, K. P. (2001). Errors in estimating tree age: Implications for studies of stand dynamics. Canadian Journal of Forest Research, 31, 1262−1271. doi: 10.1139/x01-060 [ Links ]
Recibido: 03 de Octubre de 2016; Aprobado: 30 de Marzo de 2017
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
