










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.23 no.2 Chapingo may./ago. 2017
https://doi.org/10.5154/r.rchscfa.2016.01.004
Artículos científicos
Modelos de volumen específicos y regionales para 12 especies forestales en el estado de Durango, México
1Universidad Juárez del Estado de Durango, Instituto de Silvicultura e Industria de la Madera. Bulevar del Guadiana 501, fracc. Ciudad Universitaria. C. P. 34120. Durango, México.
2Instituto Tecnológico de El Salto. Mesa del Tecnológico s/n. C. P. 34942. El Salto, Pueblo Nuevo, Durango, México.
3Universidad de Santiago de Compostela, Departamento de Enxeñería Agroforestal. Escuela Politécnica Superior, Campus Universitario 27002. Lugo, España.
Introducción:
La estimación del volumen de árboles individuales es un aspecto relevante en la dendrometría y en la realización de inventarios forestales.
Objetivo:
Se evaluaron diferencias significativas en sistemas de ecuaciones aditivas para la estimación de volumen total de árboles individuales en nueve especies de Pinus (P. cooperi, P. durangensis, P. arizonica, P. leiophylla, P. teocote, P. engelmannii, P. lumholtzii, P. strobiformis y P. herrerae), y tres de Quercus (Q. sideroxyla, Q. durifolia y Q. rugosa), y entre unidades de manejo forestal regional (UMAFOR) cuando se trata de una misma especie.
Materiales y métodos:
Para evaluar si el sistema de ecuaciones difiere entre especies de un mismo género y entre las UMAFOR para una misma especie, se utilizaron dos análisis estadísticos complementarios basados en el ajuste de un sistema reducido y un sistema completo de ecuaciones: la prueba F asociada al método de la suma adicional de cuadrados no lineales y el análisis de significancia de los parámetros.
Resultados y discusión:
Para la mayoría de las especies estudiadas, los sistemas de ecuaciones son significativamente diferentes. Se reporta la necesidad de utilizar modelos regionales en 10 de las 12 especies, exceptuando a P. strobiformis y Q. rugosa.
Palabras clave: UMAFOR; Pinus; Quercus; sistema reducido de ecuaciones; sistema completo de ecuaciones
Introduction:
Estimating the volume of individual trees is an important aspect in dendrometry and in the conducting of forest inventories.
Objective:
Significant differences were evaluated in additive equation systems for estimating the total volume of individual trees in nine species of the genus Pinus (P. cooperi, P. durangensis, P. arizonica, P. leiophylla, P. teocote, P. engelmannii, P. lumholtzii, P. strobiformisand P. herrerae), and three of the genus Quercus (Q. sideroxyla, Q. durifoliaand Q. rugosa), and among regional forest management units (UMAFOR) when dealing with the same species.
Materials and methods:
To evaluate whether equation systems are different among tree species of the same genus and among the UMAFORs for the same species, two complementary statistical analyzes were used based on the fitting of a reduced and a full equation system: the F test of nonlinear extra sum of squares method, and the parameter significance analysis.
Results and discussion:
For most of the species studied, equation systems are significantly different. The need to use regional models in 10 of the 12 tree species, except for P. strobiformis and Q. rugosa, is also reported.
Keywords: UMAFOR; Pinus; Quercus; reduced equation system; full equation system
Introducción
El estado de Durango es considerado como la primera reserva forestal de México con una estimación de 422.1 millones de m3, representando 22.4 % del total estimado a nivel nacional (Corral-Rivas, Álvarez-González, Corral-Rivas & López-Sánchez, 2015). De la superficie total del estado, 70.4 % (8.6 millones de ha) está cubierto por recursos forestales, de los cuales 57 % corresponde a bosques templados, donde dominan los rodales multiespecíficos e irregulares (Corral-Rivas, Álvarez-González, Crecente-Campo, & Corral-Rivas, 2014). De acuerdo con González, González, y Márquez (2007), en los bosques del estado de Durango se han registrado 37 especies de Quercus y 21 especies de Pinus, lo que representa 20 % de todas las especies de pino conocidas en el mundo.
En Durango, las especies que resaltan por su abundancia, distribución e importancia económica son las siguientes: Pinus cooperi Blanco, P. durangensis Martínez, P. arizonica Engelm., P. leiophylla Schiede ex Schltdl. & Cham., P. teocote Schltdl. & Cham., P. engelmannii Carr., P. lumholtzii Rob. & Fern., P. strobiformis Engelm., P. herrerae Martínez, P. maximinoi Moore, Quercus sideroxyla Bonpl., Q. durifolia Seemen y Q. rugosa Neé (Corral-Rivas, Álvarez-González, Ruiz-González, & Gadow, 2004). Estas especies se encuentran mezcladas entre sí y con otras especies maderables menos abundantes de los géneros Pinus, Quercus, Arbutus, Juniperus y Alnus, formando rodales mixtos e irregulares en las distintas UMAFOR del estado (Corral-Rivas et al., 2014).
La estimación del volumen de árboles individuales es un aspecto relevante en la dendrometría y en la realización de inventarios forestales. La dificultad en la determinación directa del volumen a través de la cubicación de secciones hace conveniente contar con ecuaciones matemáticas que permitan estimar el volumen de los árboles de manera indirecta a través de mediciones simples como el diámetro normal y la altura total del árbol en pie (Diéguez et al., 2003). En este sentido, en Durango se han desarrollado varios trabajos para la estimación del volumen de especies comerciales (Corral, Návar, & Fernández, 1999; Corral-Rivas, Barrio, Diéguez-Aranda, & Aguirre, 2007a; Corral-Rivas, Diéguez-Aranda, Castedo, & Corral-Rivas, 2007b; Quiñonez-Barraza, De los Santos-Posadas, Álvarez-González, & Velázquez-Martínez, 2014).
En México, la Secretaría del Medio Ambiente y Recursos Naturales (SEMARNAT) y la Comisión Nacional Forestal (CONAFOR) solicitan a los profesionistas forestales, como parte de una nueva política forestal, planear y ejecutar las actividades de manejo a una escala territorial conocida como unidad de manejo forestal regional (UMAFOR) (Cámara de Diputados del H. Congreso de la Unión, 2003). Una UMAFOR integra al menos 30 % de los propietarios forestales de una determinada región forestal y es delimitada considerando una similitud regional, por tanto, considera interdependencia comunal y sentido biológico.
El estado de Durango está dividido en 13 UMAFOR y para 11 de éstas se han desarrollado ecuaciones específicas de volumen a través de la ejecución de proyectos financiados por la CONAFOR. No obstante, la aplicación práctica de ecuaciones específicas en la estimación del volumen total de árboles individuales, ya sea con fines de cálculo de existencias o de cosecha, supone para los responsables del manejo forestal las interrogantes siguientes: (i) ¿es necesario usar una ecuación específica por especie para cubicar árboles de un mismo género? y (ii) ¿se requiere cambiar la ecuación específica por especie entre las distintas UMAFOR? Considerando lo anterior, este trabajo tuvo como objetivo evaluar diferencias en sistemas de ecuaciones utilizadas para la estimación del volumen en 12 especies comerciales y entre regiones forestales para una misma especie.
Materiales y métodos
El estudio se realizó con datos colectados en los años 2010 a 2013 en 11 de las 13 UMAFOR (1001 a 1011) del estado de Durango (Figura 1). Cada UMAFOR está integrada por ejidos y comunidades caracterizados por la presencia de terrenos accidentados con elevaciones y depresiones irregulares que varían de 1,400 a 3,000 m de altitud. El clima predominante es templado con lluvias en verano; la precipitación media anual oscila de 700 a 1,440 mm con temperatura promedio de 8.3 en las partes más altas y de 16 °C en las partes más bajas (Martínez-Antúnez et al., 2013).
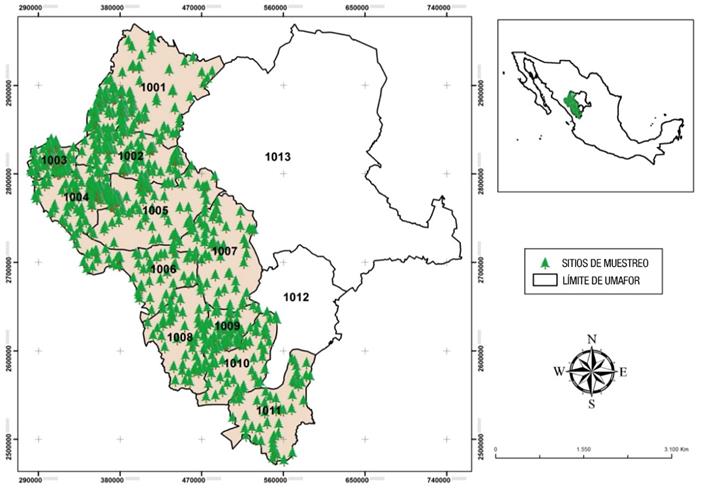
Figura 1 Localización del área de estudio. Los puntos marcados sobre el mapa representan sitios de muestreo de especies de Pinus y Quercus estudiadas en 11 unidades de manejo forestal regional (UMAFOR) del estado de Durango.
La muestra utilizada en el estudio comprende 10,828 árboles correspondientes a 12 especies forestales maderables. Los árboles se analizaron a través de muestreo destructivo (derribo y troceo) en rodales con potencial de manejo forestal que corresponden a una superficie de aproximadamente 3.4 millones de hectáreas (López-Serrano, López Sánchez, Solís-Moreno, & Corral-Rivas, 2016). La muestra tuvo condiciones variables de calidad de estación y densidad, y una distribución cercana a una normal estándar de los árboles en términos de clases de diámetro por especie. Las 12 especies estudiadas se codificaron de la manera siguiente, para ser consistentes con el catálogo de especies que se utiliza mayormente en Durango: Pinus cooperi (1), P. durangensis (2), P.arizonica (3), P. leiophylla (4), P. teocote (5), P. engelmannii (6), P.lumholtzii (7), P. strobiformis (8), P. herrerae (15), Quercus sideroxyla (41), Q. durifolia (42) y Q. rugosa (66) (Cuadro 1).
Cuadro 1 Distribución del número de árboles por especie y por unidad de manejo forestal regional (UMAFOR), considerados en el estudio en el estado de Durango.
| UMAFOR | Superficie (ha) | Especie | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 15 | 41 | 42 | 66 | Total | ||
| 1001 | 423,990 | 194 | 152 | 153 | 154 | 70 | 166 | 14 | 20 | - | 140 | - | - | 1,063 |
| 1002 | 351,498 | - | 154 | 154 | 152 | 159 | 101 | 20 | 25 | - | 102 | - | - | 867 |
| 1003 | 126,054 | 27 | 175 | 67 | 105 | - | 52 | 12 | 27 | 164 | 151 | - | - | 780 |
| 1004 | 318,104 | - | 148 | 150 | 51 | 148 | 50 | 32 | 33 | 151 | 150 | - | - | 913 |
| 1005 | 424,753 | - | 168 | 203 | 46 | 136 | - | - | 27 | - | 129 | - | - | 709 |
| 1006 | 429,806 | 130 | 256 | - | 190 | 219 | 157 | 66 | 115 | 89 | 136 | - | 64 | 1,422 |
| 1007 | 253,619 | 190 | 163 | - | 162 | 204 | 114 | 28 | 19 | - | 134 | - | - | 1,014 |
| 1008 | 373,308 | 167 | 182 | - | 173 | 176 | 167 | 108 | 107 | 148 | 163 | 138 | 16 | 1,545 |
| 1009 | 162,075 | 103 | 189 | - | 156 | 126 | 122 | - | - | 3 | 119 | 55 | - | 873 |
| 1010 | 358,944 | 132 | 119 | - | 133 | 130 | 160 | 159 | 12 | 55 | - | - | - | 900 |
| 1011 | 262,488 | 156 | 152 | - | 39 | 35 | 155 | 37 | 1 | 46 | 60 | - | 61 | 742 |
| Total | 3,484,639 | 1,099 | 1,858 | 727 | 1,361 | 1,403 | 1,244 | 476 | 386 | 656 | 1,284 | 193 | 141 | 10,828 |
1 = Pinus cooperi, 2 = P. durangensis, 3 = P. arizonica, 4 = P. leiophylla, 5 = P. teocote, 6 = P. engelmannii, 7 = P. lumholtzii, 8 = P. strobiformis, 15 = P. herrerae, 41 = Quercus sideroxyla, 42 = Q. durifolia y 66 = Q. rugosa.
En cada árbol se midieron las variables dendrométricas siguientes: i) D = diámetro normal con corteza (1.30 m sobre el nivel del suelo, cm), ii) H = altura total (m), y iii) d i = diámetro con y sin corteza para diferentes secciones a lo largo del fuste con respecto al suelo (cm). El protocolo de muestreo consideró la medición de dos secciones de 0.30 m por encima del tocón, la sección siguiente correspondió con el diámetro normal (1.30 m), continuando con secciones de 2.54 m de longitud hasta llegar a la punta del árbol, como se ilustra en la Figura 2. Adicionalmente, todas las ramas con diámetro en la base (inicio de la rama) mayor o igual a 5 cm fueron seccionadas a longitudes variables, registrando los diámetros con y sin corteza para las diferentes secciones (dr i , cm). Los árboles se cubicaron por secciones; el volumen de cada sección de fuste y de rama se estimó mediante la fórmula de Smalian y las puntas se cubicaron mediante la ecuación del cono. El volumen total del árbol se estimó sumando el volumen total con corteza del fuste y el de todas las ramas que superaron los 5 cm al inicio de las mismas. El Cuadro 2 presenta el resumen con las estadísticas descriptivas de los árboles muestra de las especies estudiadas.
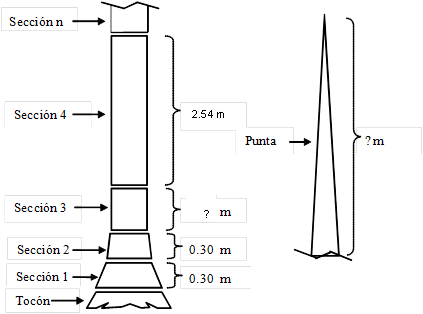
Figura 2 Ilustración de las secciones de los árboles muestra para la medición del volumen total con corteza. La altura donde termina la sección 3 corresponde a la altura del diámetro normal (1.3 m).
Cuadro 2 Resumen de las variables diámetro normal (D), altura total del árbol (H) y volumen total del árbol con corteza (vta cc ) de las especies estudiadas en 11 unidades de manejo forestal regional (UMAFOR) del estado de Durango.
| Variable | Estadísticos descriptivos | Especie | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 15 | 41 | 42 | 66 | ||
| D (cm) | Mínimo | 7.5 | 7.70 | 7.80 | 7.50 | 7.50 | 7.7 | 7.70 | 7.50 | 7.00 | 7.50 | 8.30 | 7.70 |
| Media | 30.57 | 31.10 | 33.44 | 29.75 | 29.41 | 29.18 | 24.62 | 26.23 | 31.20 | 28.39 | 26.38 | 23.30 | |
| Máximo | 67.50 | 91.24 | 88.14 | 71.60 | 82.81 | 68.00 | 58.00 | 68.00 | 73.25 | 68.00 | 70.10 | 45.50 | |
| H (m) | Mínimo | 4.19 | 3.84 | 3.97 | 3.64 | 3.97 | 3.57 | 4.18 | 5.34 | 6.38 | 3.10 | 3.77 | 4.67 |
| Media | 16.64 | 17.76 | 19.61 | 15.96 | 15.98 | 15.60 | 13.25 | 15.68 | 19.09 | 13.73 | 11.19 | 12.15 | |
| Máximo | 33.55 | 35.65 | 42.60 | 33.93 | 42.20 | 39.07 | 28.50 | 36.92 | 38.36 | 30.74 | 20.32 | 20.58 | |
| vtacc (m3) | Mínimo | 0.02 | 0.01 | 0.02 | 0.01 | 0.01 | 0.01 | 0.01 | 0.02 | 0.02 | 0.01 | 0.02 | 0.01 |
| Media | 1.01 | 1.07 | 1.39 | 0.89 | 0.86 | 0.87 | 0.56 | 0.65 | 1.12 | 0.67 | 0.51 | 0.37 | |
| Máximo | 6.24 | 12.22 | 13.42 | 6.83 | 11.16 | 6.28 | 2.97 | 5.31 | 7.24 | 4.72 | 5.70 | 1.36 | |
1= Pinus cooperi, 2 = P. durangensis, 3 = P. arizonica, 4 = P. leiophylla, 5 = P. teocote, 6 = P. engelmannii, 7 = P. lumholtzii, 8 = P. strobiformis, 15 = P. herrerae, 41 = Quercus sideroxyla, 42 = Q. durifolia y 66 = Q. rugosa.
Análisis estadístico
Un primer análisis comparativo de los datos utilizando diferentes tarifas de volumen indicó que, para más del 98 % de las especies estudiadas, la ecuación de Schumacher y Hall (1933) se ajustó mejor a la relación entre el volumen rollo total del árbol con corteza (vrta) como variable dependiente y el diámetro normal y la altura total como variables independientes. Por ello, en este trabajo se utilizó dicha ecuación como la tarifa base para comparar la estimación del volumen entre especies y en una misma especie entre las UMAFOR. El sistema de ecuaciones aditivas para estimar el volumen del fuste, de ramas y volumen total del árbol quedó integrado por las ecuaciones siguientes:

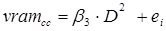
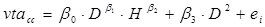
donde:
vrta cc = |
Volumen rollo total del árbol con corteza; es decir, el volumen del fuste principal (m3) |
vram cc = |
Volumen de las ramas con corteza (m3) |
vta cc = |
Volumen total del árbol con corteza (vrta cc + vram cc , m3) |
D= |
Diámetro normal del árbol (cm) |
H= |
Altura total del árbol (m) |
β i = |
Coeficientes estimados mediante la regresión |
e i = |
Error residual (m3) |
Para evaluar si el sistema de ecuaciones aditivas difiere entre especies de un mismo género y en una misma especie entre las UMAFOR, se utilizaron dos análisis complementarios basados en el ajuste de un sistema de ecuaciones reducido y de un sistema completo de ecuaciones (Bates & Watts, 1988). El sistema reducido correspondió al mismo conjunto de parámetros globales (β 0 , β 1 , β 2 y β 3 ) para todas las especies que pertenecen al mismo género sin distinguir entre ellas ni entre las UMAFOR donde una misma especie coexiste. El sistema completo correspondió a diferentes conjuntos de parámetros para cada especie o para cada UMAFOR, y se obtuvo mediante la expansión de los parámetros y la inclusión de un parámetro asociado y una variable dicotómica que permite diferenciar entre especies y entre las UMAFOR, según sea el caso. Para realizar tal prueba, el tamaño mínimo de muestra requerido por especie dentro de una UMAFOR fue 50 individuos; por tal razón, en el caso de P. strobiformis, la prueba solo se hizo en las UMAFOR 1006 y 1008.
En el caso de las nueve especies del género Pinus, los parámetros del sistema de ecuaciones aditivas se expandieron, resultando un nuevo sistema de tres ecuaciones con 36 parámetros acompañados de una variable dicotómica para diferenciar entre especies. Los parámetros del modelo completo quedaron expresados como:
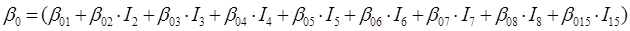
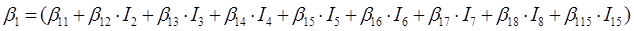
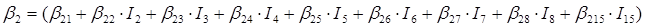
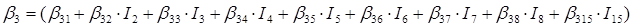
β 0i, β 1i, β 2i y β 3i son los coeficientes a estimar; I i es la variable dicotómica definida de la forma siguiente (considerando a P. cooperi como la especie base o de referencia): I 2 = 1 para la especie P. durangensis y 0 para el resto, y así sucesivamente hasta llegar a la especie codificada mediante el número 15 (P. herrerae). Cuando la comparación se hizo entre especies del género Quercus, I 2 = 1 para Q. durifolia y 0 para el resto, y así sucesivamente hasta llegar a la especie codificada con el número 66 (Q. rugosa). En este caso Q. sideroxyla fue la especie de referencia para expandir el sistema. En el caso de las UMAFOR, I 2 = 1 para la UMAFOR 1002 y 0 para el resto, y así sucesivamente hasta llegar a la UMAFOR 1011 (UMAFOR 1001 de referencia). Los dos análisis realizados se describen a continuación:
Método de la suma adicional de cuadrados no lineales
El primer análisis estadístico realizado se conoce como el método de la suma adicional de cuadrados no lineales (Bates & Watts, 1988) y la prueba estadística apropiada que sigue una distribución F se define como:
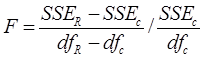
donde:
SSE R = |
Suma de cuadrados del error del modelo reducido |
df R = |
Grados de libertad asociados al modelo reducido |
SSE c = |
Suma de cuadrados del error del modelo completo |
df c = |
Grados de libertad asociados con el modelo completo |
La regla de decisión estadística en este análisis para un determinado nivel de significación (α) es la siguiente: si F > F crítico (1 − α; df R − df c , df c ) o si la probabilidad asociada (P > F crítico) < α, se rechaza la hipótesis nula (H 0: no diferencia entre especies o entre las UMAFOR). Lo anterior indica que se requieren diferentes sistemas de ecuaciones para la predicción del volumen de las distintas especies de Pinus y Quercus o para una misma especie que crece en diferentes UMAFOR. Si por el contrario F < F crítico (1 − α; df R − df c , df c ) o si la probabilidad asociada (P > Fcrítico) > α, se acepta la hipótesis nula H 0 y se propone el mismo sistema de ecuaciones para dos o más especies de un mismo género o para una misma especie en dos o más UMAFOR.
Prueba de significancia de los parámetros asociados a la variable dicotómica
El segundo análisis estadístico consistió en comprobar la significancia de los parámetros asociados a la variable dicotómica, para diferenciar entre especies y entre las UMAFOR en el sistema completo. En ambas comparaciones (entre especies y entre las UMAFOR) se estableció la hipótesis nula de que los parámetros son iguales para todas las especies o las UMAFOR (i. e. H 0 : β 0i = β 3i = 0) contra la hipótesis alternativa (H 1 ) que establece que al menos uno de los parámetros del sistema es diferente. En este análisis se rechazó H 0 cuando uno de los parámetros asociados a la variable dicotómica, para diferenciar entre especies y entre las UMAFOR, resultó significativo (P < 0.05). En este caso se requerirán diferentes sistemas de ecuaciones para predecir el volumen de dos o más especies de los géneros Pinus y Quercus o para una misma especie en diferentes UMAFOR.
Cuando en alguno de los análisis previos se detectó la existencia de diferencias significativas entre los sistemas de ecuaciones, se realizaron pruebas adicionales para evaluar si las diferencias eran causadas únicamente por dos especies o en el caso de una misma especie, solo para dos UMAFOR, tal como lo sugieren Barrio-Anta, Balboa, Castedo, Diéguez-Aranda, y Álvarez-González (2006); Calama, Cañadas, y Montero (2003); Corral-Rivas et al. (2007a); Huang, Price, y Titus (2000). Por lo tanto, se comparó un modelo completo con su correspondiente modelo reducido para todos los posibles pares de especies o de UMAFOR, a partir de los análisis descritos con anterioridad, y solamente cuando se obtuvo un valor F no significativo (en este caso, un valor de P > 0.1 para protección del error tipo II); cuando ningún parámetro asociado resultó significativo (P > 0.05) se recomendó el mismo sistema de ecuaciones para la estimación del volumen del par de especies o de las UMAFOR en estudio.
Los parámetros del modelo reducido y del modelo completo se obtuvieron de forma simultánea utilizando la metodología NSUR (Nonlinear Seemingly Unrelated Regression), mediante el procedimiento MODEL del programa SAS/ETS® (Statistical Analysis System [SAS Institute Inc.], 2008), garantizando así la aditividad de las ecuaciones de volumen.
Corrección de heterocedasticidad
Para corregir el problema de heterocedasticidad en las ecuaciones de volumen y garantizar
que las estimaciones de los parámetros se obtuvieran bajo condiciones de
homocedasticidad, el ajuste se realizó utilizando regresión ponderada,
aplicando los mismos pesos a la inversa de la varianza de cada
observación (Cunia, 1964; Parresol, 1999; Schilaegel, 1982). Dicha varianza,
a priori desconocida, se estimó utilizando una
función potencial
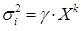 . Los valores de
. Los valores de  y del exponente
k se optimizaron empleando la metodología propuesta
por Harvey (1976) que consiste en
emplear los errores del modelo ajustado sin pesos como variable
dependiente en el modelo potencial de varianza del error. El peso
considerado fue 1/D
2
para la ecuación de volumen de las ramas con corteza
(vram
cc
) y 1/D
2
·H para la ecuación de volumen rollo total del árbol
con corteza (vrta
cc
).
y del exponente
k se optimizaron empleando la metodología propuesta
por Harvey (1976) que consiste en
emplear los errores del modelo ajustado sin pesos como variable
dependiente en el modelo potencial de varianza del error. El peso
considerado fue 1/D
2
para la ecuación de volumen de las ramas con corteza
(vram
cc
) y 1/D
2
·H para la ecuación de volumen rollo total del árbol
con corteza (vrta
cc
).
Resultados y discusión
Comparación de sistemas de ecuaciones aditivas para la estimación de volumen entre especies
Los resultados del método de la suma adicional de cuadrados no lineales obtenidos en el ajuste de los sistemas reducido y completo se muestran en el Cuadro 3. El valor del estadístico F estimado en la comparación para evaluar si el sistema de ecuaciones aditivas difiere entre las especies del género Pinus fue de 6.45, y la probabilidad de encontrar un valor crítico mayor que 6.45 fue menor que 0.0001; por lo tanto, se rechazó la hipótesis nula de que un modelo único es adecuado para todas las especies de pino juntas.
Cuadro 3 Prueba F para evaluar diferencias en el volumen de las 12 especies estudiadas en 11 unidades de manejo forestal regional (UMAFOR) del estado de Durango.
| Combinación | N | SSE C | df C | SSE R | df R | F | P > F |
|---|---|---|---|---|---|---|---|
| Todas las especies Pinus | 9,210 | 225 | 9,174 | 229.60 | 9206 | 6.45 | <0.0001 |
| 1-2 | 2,957 | 72.72 | 2,948 | 77.49 | 2,952 | 48.34 | <0.0001 |
| 1-3 | 1,826 | 48.16 | 1,818 | 63.13 | 1,822 | 141.24 | <0.0001 |
| 1-4 | 2,460 | 51.86 | 2,452 | 55.85 | 2,456 | 47.22 | <0.0001 |
| 1-5 | 2,502 | 55.99 | 2,494 | 63.21 | 2,498 | 80.45 | <0.0001 |
| 1-6 | 2,343 | 49.81 | 2,335 | 51.77 | 2,339 | 22.98 | <0.0001 |
| 1-7 | 1,575 | 36.93 | 1,567 | 39.49 | 1,571 | 27.24 | <0.0001 |
| 1-8 | 1,485 | 33.80 | 1,477 | 48.96 | 1,481 | 165.62 | <0.0001 |
| 1-15 | 1,755 | 51.49 | 1,747 | 57.66 | 1,751 | 52.29 | <0.0001 |
| 2-3 | 2,585 | 74.87 | 2,576 | 75.40 | 2,580 | 4.58 | <0.0001 |
| 2-4 | 2,219 | 56.42 | 2,905 | 56.71 | 2,909 | 3.08 | <0.0001 |
| 2-5 | 3,261 | 70.83 | 3,252 | 71.26 | 3,256 | 58.34 | <0.0001 |
| 2-6 | 3,102 | 64.62 | 3,093 | 64.99 | 3,097 | 4.42 | <0.0001 |
| 2-7 | 2,334 | 52.11 | 2,325 | 52.67 | 2,329 | 6.20 | <0.0001 |
| 2-8 | 2,244 | 48.54 | 2,235 | 56.25 | 2,239 | 88.78 | <0.0001 |
| 2-15 | 2,514 | 66.02 | 2,505 | 66.28 | 2,509 | 2.50 | 0.04 |
| 3 -4 | 2,088 | 54.27 | 2,080 | 54.87 | 2,084 | 5.68 | <0.0001 |
| 3-5 | 2,130 | 58.63 | 2,122 | 60.43 | 2,126 | 16.26 | <0.0001 |
| 3-6 | 1,971 | 48.89 | 1,808 | 49.15 | 1,812 | 2.40 | 0.04 |
| 3-7 | 1,203 | 39.71 | 1,195 | 40.52 | 1,199 | 6.11 | <0.0001 |
| 3-8 | 1,113 | 36.29 | 1,105 | 45.56 | 1,109 | 70.58 | <0.0001 |
| 3-15 | 1,383 | 53.73 | 1,375 | 55.18 | 1,379 | 9.28 | <0.0001 |
| 4-5 | 2,764 | 50.03 | 2,756 | 50.66 | 2,760 | 8.64 | <0.0001 |
| 4-6 | 2,605 | 43.77 | 2,597 | 44.10 | 2,601 | 5.00 | <0.0001 |
| 4-7 | 1,837 | 31.15 | 1,829 | 31.76 | 1,833 | 9.03 | <0.0001 |
| 4-8 | 1,747 | 27.67 | 1,739 | 34.08 | 1,743 | 100.83 | <0.0001 |
| 4-15 | 2,017 | 43.54 | 1,924 | 43.91 | 1,928 | 4.20 | <0.0001 |
| 5-6 | 2,457 | 47.95 | 2,639 | 49.44 | 2,643 | 20.49 | <0.0001 |
| 5-7 | 1,844 | 34.06 | 1,799 | 34.29 | 1,803 | 2.90 | 0.02 |
| 5-8 | 1,754 | 32.01 | 1,781 | 37.33 | 1,785 | 73.93 | <0.0001 |
| 5-15 | 2,024 | 32.13 | 2,051 | 32.38 | 2,055 | 4.09 | <0.0001 |
| 6-7 | 1,565 | 29.27 | 1,712 | 30.25 | 1,716 | 14.30 | <0.0001 |
| 6-8 | 1,475 | 25.93 | 1,622 | 34.09 | 1,626 | 127.59 | <0.0001 |
| 6-15 | 1,745 | 43.32 | 1,892 | 44.36 | 1,896 | 11.38 | <0.0001 |
| 7-8 | 862 | 13.24 | 854 | 14.62 | 858 | 22.27 | <0.0001 |
| 7-15 | 1,132 | 15.80 | 1,124 | 16.28 | 1,128 | 8.54 | <0.0001 |
| 8-15 | 1,042 | 27.33 | 1,034 | 32.56 | 1,038 | 49.55 | <0.0001 |
| Todas las especies de Quercus | 1,618 | 18.70 | 1,610 | 19.20 | 1,614 | 11.09 | <0.0001 |
| 41-42 | 1,477 | 38.08 | 1,469 | 40.15 | 1,473 | 20.05 | <0.0001 |
| 41-66 | 1,425 | 32.53 | 1,417 | 32.57 | 1,421 | 0.43 | 0.78 |
| 42-66 | 334 | 6.98 | 326 | 7.12 | 330 | 1.65 | 0.16 |
Un valor P < 0.1 indica que los modelos son diferentes. N = Número de individuos, SSEC = Suma de cuadrados del error con el modelo completo, dfC = grados de libertad asociados con el modelo completo, SSER = Suma de cuadrados del error con el modelo reducido, df R = Grados de libertad asociados con el modelo reducido. 1 = Pinus cooperi, 2 = P. durangensis, 3 = P. arizonica, 4 = P. leiophylla, 5 = P. teocote, 6 = P. engelmannii, 7 = P. lumholtzii, 8 = P. strobiformis, 15 = P. herrerae, 41 = Quercus sideroxyla, 42 = Q. durifolia y 66 = Q. rugosa.
En la comparación realizada para evaluar si el sistema de ecuaciones aditivas difiere entre las tres especies del género Quercus, el valor F fue 11.09, y la probabilidad de encontrar un valor critico mayor que 11.09 fue menor que 0.0001; por lo tanto, también se rechazó la hipótesis nula de un modelo único para las tres especies de encino.
Los resultados de las pruebas adicionales para evaluar si las diferencias son causadas solo por dos, o bien por las nueve especies de pino, se muestran también en el Cuadro 3. Se encontró que en ninguna de las 36 comparaciones pareadas se debe rechazar la hipótesis nula de un modelo único para el par analizado (P < 0.1). Por otro lado, los resultados de la evaluación pareada de las tres especies de encino indican que las especies Q. sideroxyla y Q. durifolia requieren diferentes sistemas de ecuaciones de volumen; mientras que las comparaciones Q. sideroxyla y Q. rugosa, y Q. durifolia y Q. rugosa no difirieron significativamente (P > 0.1).
Los resultados del segundo análisis mostraron que, en el ajuste del modelo completo, la mayoría de los parámetros asociados a la variable dicotómica utilizada para diferenciar entre las especies fueron significativos (P < 0.05) (Cuadro 4). Las comparaciones entre especies del género Pinus indican que al menos uno de los parámetros asociados a la variable dicotómica, para diferenciar entre especies y entre UMAFOR en el modelo completo (β 01 , β 11 , β 21 ) fue significativo (P < 0.05). En el caso de Quercus, los resultados sugieren diferencias significativas entre los modelos desarrollados para las especies Q. sideroxyla y Q. rugosa y entre Q. sideroxyla y Q. durifolia, y consistentemente con el método de la suma adicional de cuadrados no lineales no existen diferencias significativas entre los sistemas de ecuaciones desarrolladas para Q. sideroxyla y Q. rugosa.
Cuadro 4 Parámetros asociados a la variable dicotómica utilizada para diferenciar entre especies (β 01 , β 11 y β 21 ).
| Comparación | β 0 | β 01 | β 1 | β 11 | β 2 | β 21 | β 3 | Β 31 |
|---|---|---|---|---|---|---|---|---|
| 1-2 | 4.91E-05 | 1.08E-05 | 1.9855 | 0.0085 | 0.9771 | -0.0870 | 6.21E-05 | -1.75E-05 |
| 1-4 | 4.84E-05 | 9.25E-06 | 1.9875 | 0.0340 | 0.9791 | -0.1112 | 6.09E-05 | -1.64E-05 |
| 1-5 | 4.92E-05 | 2.02E-05 | 1.9809 | -0.0410 | 0.9820 | -0.0803 | 6.14E-05 | -1.7E-05 |
| 1-6 | 4.87E-05 | 6.15E-06 | 1.9862 | 0.0142 | 0.9790 | -0.0686 | 6.1E-05 | -6.83E-07 |
| 1-7 | 4.89E-05 | 2.78E-05 | 1.9844 | 0.0351 | 0.9798 | -0.2221 | 6.13E-05 | 2.58E-05 |
| 1-8 | 4.83E-05 | 1.78E-05 | 1.9903 | -0.0226 | 0.9765 | -0.1310 | 6.1E-05 | -1.94E-05 |
| 1-15 | 4.88E-05 | 1.13E-05 | 1.9812 | -0.0035 | 0.9835 | -0.0852 | 6E-05 | -1.39E-05 |
| 2-3 | 6.08E-05 | 2.3E-06 | 1.9878 | -0.0032 | 0.8938 | -0.0003 | 4.53E-05 | -1.45E-05 |
| 2-4 | 6.04E-05 | -2.4E-06 | 1.9911 | 0.0206 | 0.8916 | -0.0128 | 4.49E-05 | 4.57E-07 |
| 2-5 | 6.07E-05 | 9.17E-06 | 1.9884 | -0.0517 | 0.8932 | 0.0099 | 4.5E-05 | 1.44E-06 |
| 2-6 | 6.05E-05 | -4.64E-06 | 1.9900 | 0.0006 | 0.8921 | 0.0240 | 4.48E-05 | 1.68E-05 |
| 2-7 | 6.09E-05 | 1.7E-05 | 1.9866 | 0.0059 | 0.8942 | -0.1080 | 4.51E-05 | 4.31E-05 |
| 2-8 | 6.07E-05 | 6.05E-06 | 1.9893 | -0.0481 | 0.8920 | -0.0181 | 4.52E-05 | -2.52E-06 |
| 2-15 | 6.02E-05 | 1.12E-06 | 1.9898 | -0.0130 | 0.8939 | -0.0002 | 4.47E-05 | 3.9E-06 |
| 3-4 | 6.32E-05 | -5.39E-06 | 1.9854 | 0.0140 | 0.8921 | 0.0029 | 3.04E-05 | 1.48E-05 |
| 3-5 | 6.32E-05 | 5.86E-06 | 1.9846 | -0.0426 | 0.8931 | 0.0081 | 3.06E-05 | 1.65E-05 |
| 3-6 | 6.32E-05 | -6.98E-06 | 1.9851 | 0.0035 | 0.8923 | 0.0241 | 3.02E-05 | 3.15E-05 |
| 3-7 | 6.3E-05 | 1.21E-05 | 1.9829 | 0.0095 | 0.8961 | -0.0936 | 3.06E-05 | 5.93E-05 |
| 3-8 | 6.27E-05 | 2.8E-06 | 1.9846 | -0.0599 | 0.8952 | 0.0055 | 3.09E-05 | 1.22E-05 |
| 3-15 | 6.26E-05 | -1.82E-06 | 1.9850 | 0.0020 | 0.8955 | -0.0105 | 2.99E-05 | 1.94E-05 |
| 4-5 | 5.85E-05 | 1.17E-05 | 2.0132 | -0.0778 | 0.8740 | 0.0297 | 4.5E-05 | 4.83E-07 |
| 4-6 | 5.84E-05 | -3.11E-06 | 2.0206 | -0.0270 | 0.8652 | 0.0512 | 4.47E-05 | 1.61E-05 |
| 4-7 | 5.87E-05 | 2.15E-05 | 2.0203 | -0.0278 | 0.8636 | -0.0892 | 4.49E-05 | 4.19E-05 |
| 4-8 | 5.82E-05 | 8.59E-06 | 2.0219 | -0.0639 | 0.8646 | -0.0107 | 4.5E-05 | -2.92E-06 |
| 4-15 | 5.83E-05 | 2.7E-06 | 2.0079 | -0.0353 | 0.8811 | 0.0194 | 4.42E-05 | 2.86E-06 |
| 5-6 | 7.06E-05 | -1.47E-05 | 1.9311 | 0.0552 | 0.9070 | 0.0144 | 4.45E-05 | 1.66E-05 |
| 5-7 | 7.14E-05 | 9.47E-06 | 1.9222 | 0.0540 | 0.9144 | -0.1217 | 4.5E-05 | 4.16E-05 |
| 5-8 | 7.06E-05 | -3.71E-06 | 1.9329 | 0.0078 | 0.9047 | -0.0305 | 4.6E-05 | -3.92E-06 |
| 5-15 | 6.94E-05 | -7.74E-06 | 1.9340 | 0.0369 | 0.9091 | -0.0099 | 4.44E-05 | 3.12E-06 |
| 6-7 | 5.6E-05 | 2.63E-05 | 1.9822 | -0.0039 | 0.9261 | -0.1431 | 6.07E-05 | 2.29E-05 |
| 6-8 | 5.56E-05 | 1.17E-05 | 1.9903 | -0.0373 | 0.9185 | -0.0607 | 6.09E-05 | -1.98E-05 |
| 6-15 | 5.55E-05 | 6.16E-06 | 1.9894 | -0.0191 | 0.9198 | -0.0198 | 6.02E-05 | -1.42E-05 |
| 7-8 | 8.52E-05 | -1.66E-05 | 1.9549 | -0.0113 | 0.7988 | 0.0641 | 8.38E-05 | -4.23E-05 |
| 7-15 | 7.73E-05 | -1.53E-05 | 1.9952 | -0.0318 | 0.7864 | 0.1203 | 8.71E-05 | -4.11E-05 |
| 8-15 | 6.53E-05 | -4.29E-06 | 1.9424 | 0.0306 | 0.8806 | 0.0195 | 4.14E-05 | 5.5E-06 |
| 41-42 | 5.02E-05 | 2.62E-05 | 1.9503 | -0.1631 | 0.9562 | 0.0370 | 7.57E-05 | 0.00015 |
| 41-66 | 5.02E-05 | 2.62E-05 | 1.9503 | -0.1631 | 0.9562 | 0.0370 | 7.57E-05 | 0.00015 |
| 42-66 | 8.11E-05 | -2.14E-05 | 1.7838 | 0.1550 | 0.9698 | -0.0705 | 0.000219 | -0.000133 |
Los parámetros marcados en negritas fueron no significativos (P > 0.05). Nótese que en la comparación entre los sistemas de ecuaciones de Q. sideroxyla y Q. rugosa, ninguno de los parámetros aditivos resultó significativo. 1 = Pinus cooperi, 2 = P. durangensis, 3 = P. arizonica, 4 = P. leiophylla, 5 = P. teocote, 6 = P. engelmannii, 7 = P. lumholtzii, 8 = P. strobiformis, 15 = P. herrerae, 41 = Quercus sideroxyla, 42 = Q. durifolia y 66 = Q. rugosa.
Los resultados de los dos análisis estadísticos para evaluar diferencias en volumen de las 12 especies estudiadas fueron consistentes en 97.4 % de los casos estudiados, indicando el rechazo de la hipótesis nula de un modelo único. Únicamente se observó una discrepancia entre las dos pruebas estadísticas realizadas, la cual consistió en que la suma adicional de cuadrados no lineales recomienda el uso de un modelo reducido para estimar el volumen de Q. durifolia y Q. rugosa, mientras que el análisis de la significancia de los parámetros recomienda emplear modelos diferentes.
En este trabajo se decidió rechazar la hipótesis nula de un modelo único, si en alguna de las dos pruebas estadísticas realizadas se sugiere el rechazo. Lo anterior se fundamentó en el hecho de que se cuenta con información a nivel de especie para el desarrollo de los modelos, y de acuerdo con los resultados de este trabajo solamente el volumen de las especies Q. sideroxyla y Q. rugosa podría estimarse con un mismo sistema de ecuaciones.
Los resultados de la comparación entre especies de un mismo género son consistentes con los reportados por Corral-Rivas et al. (2007a) y Corral-Rivas et al. (2007b), quienes observaron también diferencias significativas en modelos desarrollados para estimar el diámetro normal y el volumen a partir de las dimensiones del tocón, y en funciones compatibles de ahusamiento y volumen en cinco especies del género Pinus en una región forestal de Durango, respectivamente. Corral-Rivas et al. (2004) también reportaron diferencias significativas en el crecimiento en altura dominante en cinco de las especies del género Pinus analizadas en este estudio. Por su parte, Quiñonez-Barraza et al. (2014) encontraron diferencias en la altura a la que ocurren los puntos de inflexión de ecuaciones de perfil en algunas de las especies consideradas en este trabajo.
Las principales causas de las diferencias estadísticas observadas en el volumen de las especies estudiadas se relacionan con diferencias en el perfil del fuste (Corral-Rivas et al., 2007b; Quiñonez-Barraza et al., 2014) y con requerimientos específicos de calidad de estación de las especies analizadas (Corral-Rivas et al., 2004).
Comparación de sistemas de ecuaciones aditivas para la estimación de volumen entre ecorregiones
El valor del estadístico F calculado en la comparación realizada para evaluar si los sistemas de ecuaciones para una misma especie difieren entre las UMAFOR fue menor que el valor critico (P < 0.1) en todas las especies analizadas, con excepción de P. strobiformis y Q. rugosa (Cuadro 5). Por tanto, se rechaza la hipótesis nula de la formulación de un único modelo para todas las UMAFOR en 10 de las 12 especies estudiadas. Los resultados de las pruebas adicionales realizadas para evaluar si las diferencias son causadas únicamente por dos o por más de las UMAFOR no se muestran en este artículo debido al espacio que ocuparían; sin embargo, indican que en 111 de las 285 comparaciones pareadas no fue posible rechazar la hipótesis nula de un modelo único para el par de UMAFOR analizado (P > 0.1), mientras que en el resto (61 % de los pares) se rechaza la hipótesis nula de un modelo único para los pares de UMAFOR analizados.
Cuadro 5 Resultados de la suma adicional de cuadrados no lineales utilizada para evaluar diferencias regionales en el volumen de las 12 especies estudiadas en 11 unidades de manejo forestal regional (UMAFOR) del estado de Durango.
| Especie | UMAFOR | N | SSE C | df C | SSE R | df R | F | P > F |
|---|---|---|---|---|---|---|---|---|
| 1 | 7 | 1,072 | 25.04 | 1,040 | 28.51 | 1,068 | 5.10 | <0.0001 |
| 2 | 11 | 1,704 | 39.10 | 1,661 | 41.14 | 1,700 | 2.20 | <0.0001 |
| 3 | 5 | 727 | 29.63 | 719 | 31.38 | 723 | 10.56 | <0.0001 |
| 4 | 9 | 1,170 | 16.33 | 1,162 | 20.23 | 1,166 | 69.31 | <0.0001 |
| 5 | 9 | 1,368 | 20.79 | 1,360 | 26.68 | 1,364 | 96.23 | <0.0001 |
| 6 | 10 | 1,244 | 16.60 | 1,236 | 21.02 | 1,240 | 82.37 | <0.0001 |
| 7 | 3 | 333 | 2.00 | 325 | 2.54 | 329 | 22.00 | <0.0001 |
| 8 | 2 | 222 | 0.83 | 214 | 0.86 | 218 | 1.99 | 0.10 |
| 15 | 6 | 653 | 19.15 | 645 | 22.31 | 649 | 26.66 | <0.0001 |
| 41 | 10 | 1,284 | 28.40 | 1,276 | 31.59 | 1,280 | 35.84 | <0.0001 |
| 42 | 2 | 193 | 1.65 | 185 | 6.09 | 189 | 124.60 | <0.0001 |
| 66 | 2 | 125 | 0.91 | 117 | 0.96 | 120 | 2.10 | 0.10 |
*No se rechaza la hipótesis nula de un modelo único (P ≥ 01). N = Número de individuos, SSEC = Suma de cuadrados del error con el modelo completo, dfC = Grados de libertad asociados con el modelo completo, SSER = Suma de cuadrados del error con el modelo reducido, df R = Grados de libertad asociados con el modelo reducido. 1 = Pinus cooperi, 2 = P. durangensis, 3 = P. arizonica, 4 = P. leiophylla, 5 = P. teocote, 6 = P. engelmannii, 7 = P. lumholtzii, 8 = P. strobiformis, 15 = P. herrerae, 41 = Quercus sideroxyla, 42 = Q. durifolia y 66 = Q. rugosa.
Los resultados del análisis de la significancia de los parámetros tampoco se muestran por motivos de espacio, pero se observó que los parámetros asociados a la variable dicotómica utilizada para diferenciar entre UMAFOR (β 01, β 11, β 21) fueron significativos (P < 0.05) en la mayoría de los ajustes del modelo completo. De acuerdo con este análisis, solo en 15 comparaciones realizadas entre UMAFOR (5 % de los casos) no fue posible rechazar la hipótesis nula de la formulación de un modelo único.
En relación con el análisis anterior, resulta interesante la inexistencia de diferencias regionales en el volumen de P. strobiformis; dicho resultado puede estar relacionado con una similitud de condiciones ecológicas de los sitios donde esta especie se desarrolla, ya que crece normalmente en áreas húmedas y con sombra suficiente (Aguilera, 2001). Las diferencias encontradas en volumen dentro de una especie que crece en más de una UMAFOR se atribuyen a diferencias en la densidad residual, causadas principalmente por la aplicación de los tratamientos silvícolas (Larson, 1963), al tipo de suelo y a la precipitación. Castedo, Barrio, Parresol, y Álvarez-González (2005) recomendaron una ecuación diferente para modelar la relación altura-diámetro de Pinus pinaster Ait, en algunas regiones forestales del norte de España. Calama et al. (2003) proponen también diferentes ecuaciones de índice de sitio para Pinus pinea L. en regiones forestales de España. Hasta la fecha, en México no se han reportado estudios que evalúen diferencias regionales en algún tipo de modelo de crecimiento forestal, por lo que existe la necesidad de realizar otros trabajos que confirmen los resultados encontrados, especialmente para la especie P. strobiformis, debido a que en este estudio se contó con un tamaño de muestra adecuado solo para dos UMAFOR (1006 y 1008).
El Cuadro 6 presenta los estimadores de los parámetros derivados del ajuste, el coeficiente de determinación ajustado (R2 aj) y la raíz del error medio cuadrático (REMC) para las distintas ecuaciones que integran los sistemas de ecuaciones aditivas desarrolladas en este trabajo. Estos sistemas de ecuaciones explicaron en promedio 97, 34 y 96 % de la varianza observada en el volumen rollo total del árbol, volumen de ramas y volumen total árbol, respectivamente.
Cuadro 6 Parámetros estimados en los sistemas de ecuaciones aditivas de volumen desarrollados para 12 especies maderables en el estado de Durango.
| Especie | Parámetros | vrta cc | vramas cc | vta cc | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| β 0 | β 1 | β 2 | β 3 | REMC | R 2 aj. | REMC | R 2 aj. | REMC | R 2 aj. | |
| 1 | 0.000058 | 1.934384 | 0.986875 | 0.000059 | 0.131 | 0.98 | 0.105 | 0.35 | 0.165 | 0.97 |
| 2 | 0.000061 | 1.970438 | 0.916467 | 0.000042 | 0.140 | 0.98 | 0.070 | 0.34 | 0.153 | 0.98 |
| 3 | 0.000063 | 1.973039 | 0.908113 | 0.000031 | 0.198 | 0.98 | 0.072 | 0.18 | 0.208 | 0.98 |
| 4 | 0.000062 | 1.989640 | 0.884421 | 0.000044 | 0.111 | 0.98 | 0.081 | 0.31 | 0.132 | 0.98 |
| 5 | 0.000071 | 1.903127 | 0.939725 | 0.000048 | 0.119 | 0.98 | 0.078 | 0.29 | 0.14 | 0.97 |
| 6 | 0.000057 | 1.984667 | 0.922042 | 0.000061 | 0.097 | 0.99 | 0.086 | 0.33 | 0.132 | 0.98 |
| 7 | 0.000088 | 1.893291 | 0.862257 | 0.000084 | 0.086 | 0.97 | 0.081 | 0.36 | 0.131 | 0.95 |
| 8 | 0.000070 | 1.913666 | 0.894689 | 0.000036 | 0.102 | 0.98 | 0.053 | 0.27 | 0.112 | 0.98 |
| 15 | 0.000061 | 1.900093 | 0.984575 | 0.000064 | 0.135 | 0.98 | 0.097 | 0.40 | 0.185 | 0.97 |
| 41 | 0.000050 | 1.974851 | 0.928771 | 0.000065 | 0.115 | 0.97 | 0.111 | 0.27 | 0.157 | 0.96 |
| 42 | 0.000030 | 1.964724 | 1.101723 | 0.000168 | 0.084 | 0.95 | 0.185 | 0.48 | 0.196 | 0.89 |
| 66 | 0.000053 | 1.981578 | 0.887948 | 0.000078 | 0.065 | 0.95 | 0.046 | 0.55 | 0.084 | 0.94 |
1 = Pinus cooperi, 2 = P. durangensis, 3 = P. arizonica, 4 = P. leiophylla, 5 = P. teocote, 6 = P. engelmannii, 7 = P. lumholtzii, 8 = P. strobiformis, 15 = P. herrerae, 41 = Quercus sideroxyla, 42 = Q. durifolia y66 = Q. rugosa. vrta cc = Volumen rollo total del árbol con corteza (m3), vramas cc = Volumen de ramas con corteza (m3), vta cc = Volumen total del árbol con corteza (m3), REMC = Raíz del error medio cuadrático (m3) y R2aj = Coeficiente de determinación ajustado.
Conclusiones
Los resultados de las pruebas estadísticas correspondientes al método de la suma adicional de cuadrados no lineales y de la significancia de los parámetros indican la necesidad de emplear modelos específicos para cada una de las especies analizadas, y regionales en 10 de ellas. Las pruebas adicionales para evaluar si la existencia de diferencias es causada únicamente por dos o por las nueve especies del género Pinus, señalan que cada una requiere su propio modelo, mientras que las comparaciones entre las especies del género Quercus indican que Q. sideroxyla y Q. rugosa no difieren significativamente. Por otra parte, solo en 15 de 285 pares (5 % de los casos) de las UMAFOR no fue posible rechazar la hipótesis de un modelo único para evaluar diferencias regionales en el volumen de las especies. De acuerdo con los resultados se recomienda el empleo de los modelos específicos y regionales desarrollados en este trabajo para las especies siguientes: P. cooperi, P. durangensis, P. arizonica, P. leiophylla, P. teocote, P. engelmannii, P. lumholtzii y Q. sideroxyla; y de un modelo estatal para P. strobiformis y Q. rugosa, hasta que se cuente con más datos que permitan confirmar este resultado.
Agradecimientos
Al Programa de Fomento a la Organización Social, Planeación y Desarrollo Regional Forestal (PROFOS) de la CONAFOR por financiar la colecta de la información de campo utilizada en este trabajo.
REFERENCIAS
Aguilera, R. M. (2001). Pinus ayacahuite Ehren, paquete tecnológico. México: CONAFOR. [ Links ]
Barrio-Anta, M., Balboa, M., Castedo, D. F., Diéguez-Aranda, U., & Álvarez-González, J. G. (2006). An ecoregional model for estimating volume, biomass and carbon pools in maritime pine stands in Galicia (northwestern Spain). Forest Ecology and Management, 223, 24‒34. doi: 10.1016/j.foreco.2005.10.073 [ Links ]
Bates, D. M., & Watts, D. G. (1988). Nonlinear regression analysis and its applications. New York, USA: Wiley. [ Links ]
Calama, R., Cañadas, N., & Montero, G. (2003). Inter-regional variability in site index models for even-aged stands of stone pine (Pinus pinea L.) in Spain. Annals of Forest Science, 60, 259-269. doi: 10.1051/forest:2003017 [ Links ]
Castedo, F., Barrio, M., Parresol, B. R., & Álvarez-González, J. G. (2005). A stochastic height-diameter model for maritime pine ecoregions in Galicia (north-western Spain). Annals of Forest Science, 62, 455-465. doi: 10.1051/forest:2005042 [ Links ]
Corral-Rivas, J. J., Álvarez-González, J. G., Ruíz-González, A. D., & Gadow, Kv. (2004). Compatible height and site index models for five pine species in El Salto Durango (Mexico). Forest Ecology and Management , 201, 145-160. doi: 10.1016/j.foreco.2004.05.060 [ Links ]
Corral-Rivas, J. J., Barrio, A. M., Diéguez-Aranda, U., & Aguirre, C. O. (2007a). Use of stump diameter to estimate diameter at breast height and tree volume for major pines species in El Salto, Durango (Mexico). Forestry, 80, 29‒40. doi: 10.1093/forestry/cpl048 [ Links ]
Corral-Rivas, J. J., Diéguez-Aranda, U., Castedo, F., & Corral-Rivas, S. (2007b). A merchantable volume system for major pine species in El Salto, Durango (Mexico). Forest Ecology and Management , 238, 118‒129. doi: 10.1016/j.foreco.2006.09.074 [ Links ]
Corral-Rivas, S., Álvarez-González J. G., Crecente-Campo, F., & Corral-Rivas, J.J. (2014). Local and generalized height-diameter models with random parameters for mixed, uneven-aged forests in Northwestern Durango, Mexico. Forest Ecosystems, 1(6). doi: 10.1186/2197-5620-1-6 [ Links ]
Corral-Rivas, S., Álvarez-González, J. G., Corral-Rivas, J. J., & López-Sánchez, C. A. (2015). Characterization of diameter structures of natural forests of northwest of Durango, Mexico. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 21(2), 221-236. doi: 10.5154/r.rchscfa.2014.10.046 [ Links ]
Corral, R. S., Návar, J. J., & Fernández, S. F. (1999). Ajuste de funciones de ahusamiento a los perfiles fustales de cinco Pináceas de la región de El Salto, Durango. Madera y Bosques, 5, 53‒65. Retrieved from http://www.redalyc.org/pdf/617/61750205.pdf [ Links ]
Cunia, T. (1964). Weighted least squares methods and construction of volume tables. Forest Science, 10, 180‒191. [ Links ]
Cámara de Diputados del H. Congreso de la Unión. (2003). Ley de desarrollo forestal sustentable. México: Diario Oficial de la Federación. Retrieved from http://www.diputados.gob.mx/LeyesBiblio/pdf/259_100516.pdf [ Links ]
Diéguez, A. U., Barrio, A. M., Castedo, D. F., Ruíz, G. A. D., Álvarez, T. M. F., Álvarez, G. J. G., & Rojo, A. (2003). Dendrometría. Madrid, España: Fundación Conde del Valle de Salazar-Ediciones Mundi-Prensa. [ Links ]
González, M. S., González, M., & Márquez, M. A. (2007). Vegetación y ecorregiones de Durango. México: Plaza y Valdés Editores-Instituto Politécnico Nacional. [ Links ]
Harvey, A. C. (1976). Estimating regression models whit multiplicative heteroscedastiticity. Econometrica, 44, 461‒465. doi: 10.2307/1913974 [ Links ]
Huang, S., Price, D., & Titus, S. J. (2000). Development of ecoregion-based height-diameter models for white spruce in boreal forests. Forest Ecology and Management , 129, 125-141. doi: 10.1016/S0378-1127(99)00151-6 [ Links ]
Larson, P. R. (1963). Stem form development of forest trees. USA: Society of American Foresters. [ Links ]
López-Serrano, P., López Sánchez, C. A., Solís-Moreno, R., & Corral-Rivas, J. J. (2016). Geospatial estimation of above ground forest biomass in the Sierra Madre Occidental in the State of Durango, Mexico. Forests, 7(70), 1‒13. doi: 10.3390/f7030070 [ Links ]
Martínez-Antúnez, P., Wehenkel, C., Hernández-Díaz, J. C., González-Elizondo, M., Corral-Rivas, J. J., & Pinedo-Álvarez, A. (2013).Effect of climate and physiography on the density of tree and shrub species in Northwest Mexico. Polish Journal Ecology, 61(2), 283-295. Retrieved from https://www.researchgate.net/publication/249963406_Effect_of_climate_and_physiography_on_the_density_of_tree_and_shrub_species_in_Northwest_Mexico [ Links ]
Parresol, B. R. (1999). Assessing tree and stand biomass: A review whit examples and critical comparisons. Forest Science , 45, 573‒593. Retrieved from http://www.sidalc.net/repdoc/A11144i/A11144i.pdf [ Links ]
Quiñonez-Barraza, G., De los Santos-Posadas, H. M., Álvarez-González, J. G., & Velázquez-Martínez, A. (2014). Sistema compatible de ahusamiento y volumen comercial para las principales especies de Pinus en Durango, México. Agrociencia, 48, 553‒567. Retrieved from http://www.scielo.org.mx/pdf/agro/v48n5/v48n5a8.pdf [ Links ]
Schilaegel, B. E. (1982). Acer negundo biomasas componet regression analysis for the Mississipi Delta. Forest Science , 28, 355‒358. [ Links ]
Schumacher, F. X., & Hall, F. D. S. (1933). Logarihmic expression of timber-tree volume. Journal of Agricultural Research, 47(9), 719‒734. Retrieved from https://naldc.nal.usda.gov/download/IND43968352/PDF [ Links ]
Statistical Analysis System Institute Inc. (SAS). (2008). SAS/ETS® 9.2. User’s guide. Cary, NC, USA: Author. [ Links ]
Recibido: 26 de Enero de 2016; Aprobado: 11 de Diciembre de 2016
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
