










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versão On-line ISSN 2007-4018versão impressa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.22 no.3 Chapingo Set./Dez. 2016
https://doi.org/10.5154/r.rchscfa.2015.10.048
Series de incremento del área basal en árboles dominantes de Pseudotsuga menziesii (Mirb.) Franco muestra periodicidad de acuerdo con patrones climáticos globales
1Colegio de Postgraduados, Campus Montecillo. Carretera México-Texcoco km 36.5. C. P. 56230. Montecillo, Texcoco, Estado de México, México.
2Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Centro Nacional de Investigación Disciplinaria en Relación Agua, Suelo, Planta, Atmósfera (CENID RASPA). km 6.5 Margen Derecha Canal Sacramento. C. P. 35140. Gómez Palacio, Durango. México.
Las especies forestales como Pseudotsuga menziesii (Mirb.) Franco son sensibles al clima y muestran anillos de crecimiento claramente definidos. La selección cuidadosa de árboles dominantes con fuste circular permite el análisis de tendencias de crecimiento arbóreo. En este estudio se utilizaron mediciones directas del incremento del área basal (IAB) para explicar las periodicidades biológicas y elaborar predicciones del crecimiento en el abeto Douglas-fir que crece en el oeste de México. Para eliminar el efecto de la edad en el crecimiento de los árboles se hizo un análisis en términos de la edad del cámbium. Los resultados mostraron correlación significativa (P < 0.05) entre IAB y la precipitación de enero a julio. Además, se encontraron periodicidades de 7, 21, 27 y 60 años en el crecimiento de los árboles; el periodo de 60 años fue determinante para la construcción de un modelo ARIMA (0,1,1) para realizar predicciones del IAB en las próximas décadas. Las Proyecciones del crecimiento proponen una reducción del IAB en árboles maduros dominantes en las próximas décadas. Dicha reducción es un resultado inesperado, debido a que el IAB en árboles dominantes permanece constante hasta una edad biológica de senescencia. Los resultados concuerdan con una tendencia general de reducción en el crecimiento en otros bosques del mundo debido a estrés hídrico, lo cual sugiere que la variabilidad climática futura puede empeorar la condición de salud del abeto Douglas-fir de los bosques del norte de México.
Palabras clave: Dendrocronología; anillos de crecimiento; productividad forestal; bosques templados; análisis de series de tiempo.
Tree species like Pseudotsuga menziesii (Mirb.) Franco (Douglas-fir) are sensitive to climate cycles and show well defined growth rings. The careful selection of dominant trees with circular Ttrunk section allow the analysis of tree growth trajectories. In this study, we used direct measurements of basal area increment (BAI) to explain biological periodicity and forecast basal area growth of Douglas-fir growing in Western Mexico. To remove the age effect on tree growth we also ran the analysis in terms of cambial age. Results showed significant (P < 0.05) correlation between BAI and precipitation from January to July. We found periodicities in tree growth of 7, 21, 27 and 60 years. However, the 60-year period, was determinant to build an ARIMA model (0,1,1), to forecast BAI for the next decades. Tree growth projections suggest reduced BAI in mature dominant trees for the next decades. Decreased tree-growth is an unexpected result, as BAI in dominant trees remains constant up to the biological age. Our finding is concurrent with a general decrement in tree growth in other forests of the world due to water stress, which suggests that the future climatic variability may worsen health conditions of Douglas-fir forests in North Mexico.
Keywords: Dendrochronology; tree-ring; forest productivity; temperate forests; time series analysis
Introducción
Debido al aumento de la variabilidad del clima en las últimas tres décadas (Hansen, Sato, & Ruedy, 2012), se requiere una mayor comprensión sobre la relación entre el ancho de anillos de árboles, el crecimiento del área basal y las variables climáticas con la finalidad de proponer mejores prácticas forestales. El establecimiento de la relación entre las mediciones de los anillos de los árboles y los incrementos en el área basal (IAB) con las variables climáticas permite la identificación de patrones de crecimiento. El crecimiento forestal depende de la especie, edad, densidad de masa, calidad del suelo y clima (Skovsgaard & Vanclay, 2008), y el efecto combinado de estos factores se registra en el ancho de anillos de la especie (Esper, Benz, & Pederson, 2012; Matisons, Elferts, & Brūmelis, 2013). Los anillos de los árboles de especies sensibles al clima conservan información acerca de los patrones cíclicos de crecimiento y de las relaciones con el clima, proporcionando información útil para propósitos de modelación y predicción (Matisons et al., 2013; Návar & Lizárraga-Mendiola, 2013).
Una de las especies más importantes para los propósitos de la dendrocronología ha sido el abeto Douglas (Pseudotsuga menziesii [Mirb.] Franco), debido a su sensibilidad para captar señales de eventos cíclicos del clima (Cerano, Villanueva, Valdéz, Méndez, & Constante, 2011; González-Elizondo et al., 2005; Griesbauer & Scott, 2010; Gugger, González-Rodríguez, Rodríguez-Correa, Sugita, & Cavender-Bares, 2011), pero la confiabilidad de esa periodicidad no se ha estudiado en modelos de predicción. El análisis de serie de tiempo de la especie abeto Douglas ayudaría a confirmar los eventos extremos pasados en patrones de circulación atmosférica, escasez de recursos hídricos e impacto socioeconómico en la población (Cerano et al., 2011; Návar & Lizárraga-Mendiola, 2013). Recientemente, el uso del análisis de series de tiempo, combinado con isótopos de madera, y la dendrocronología han demostrado ser útiles para describir la relación de la fisiología de los árboles y la variabilidad del clima de acuerdo con el aumento de CO2 atmosférico (Monserud & Marshall, 2001; Roden, Johnstone, & Dawson, 2011; Swetnam & Lynch, 1993).
Los procedimientos básicos de la dendrocronología utilizan el índice de ancho de anillos (IAA) para analizar las tendencias en el crecimiento del árbol; sin embargo, cuando se seleccionan árboles dominantes con troncos de sección circular, las mediciones directas de IAB están altamente correlacionadas con IAA, permitiendo el estudio de las tendencias de crecimiento. En comparación con el ancho de anillo, el IAB es más conveniente para medir las tasas de crecimiento de los árboles, ya que se trata de una variable bidimensional (Biondi & Qeadan, 2008). Cuando la serie de anillos de árboles se transforman en incremento del área basal (IAB), el uso práctico es que el IAB tiene unidades en cm2•año-1 que permiten la interpretación directa de las tendencias de crecimiento; en cambio, los valores IAA son unidades relativas y adimensionales (Rubino & McCarthy, 2000).
Las herramientas del análisis de series de tiempo se utilizan con frecuencia en estudios econométricos; sin embargo, debido a la variación climática reciente y sus efectos en el crecimiento de los árboles, el uso del análisis de series de tiempo se debe revaluar como una herramienta poderosa para comprender las relaciones de cambio climático y crecimiento de árboles.
Este estudio tuvo como objetivo analizar las tendencias para IAB de bosques mixtos de abeto Douglas utilizando modelos de la series de tiempo para predecir el crecimiento de árboles para las próximas décadas. Nuestra hipótesis es que la serie del IAB de P. menziesii en la Sierra Madre Occidental refleja eventos climáticos regionales y es confiable para construir un modelo con la finalidad de explicar y predecir el crecimiento del área basal de los árboles.
Materiales y métodos
Área de estudio
El sitio de estudio se encuentra en la Sierra Madre Occidental en el estado de Chihuahua (26° 27’ 59.14’’ N y 106° 21’ 14.95’’ O). La altitud oscila entre 2,650 a 2,730 m, el clima es sub-húmedo templado con una temperatura media anual de 12 ° C y precipitación de 780 mm (Figura 1). Las especies de árboles asociados del sitio de estudio son Picea chihuahuana Martínez, Pinus arizonica Engelm., Pinus durangensis Martínez y Pinus ayacahuite Ehrenb. (García-Arévalo, 2008; González, González, & Márquez, 2007). No se registró aprovechamiento forestal en el área de estudio; P. menziesii, especie que ha estado bajo conservación, es una especie en peligro de extinción de acuerdo con la norma mexicana (NOM-059-SEMARNAT-2010). La altura total del abeto Douglas en el estrato alto oscila entre 23 y 45 m, con DAP (diámetro a la altura del pecho) de 60-104 cm. La masa mixta del sitio de estudio tiene una existencia estimada de 8.4 m3•ha-1 de tronco de la especie estudiada.
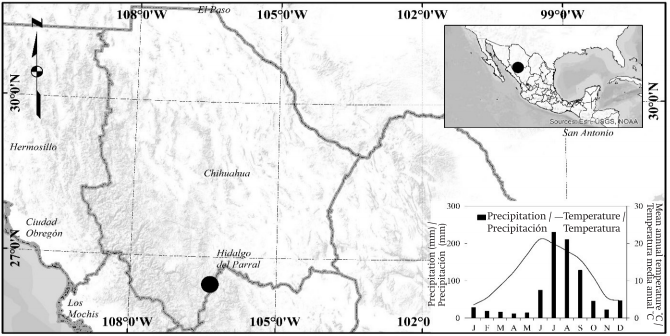
Muestreo del núcleo de la madera
En el verano de 2012 se tomaron muestras de 20 árboles sanos y dominantes cuidadosamente seleccionados por su sección circular del tronco, utilizando un taladro de 12 mm (Haglöf, BS009, Alemania). Para cada árbol se recolectó una muestra en dirección transversal a la pendiente, a 1.3 m por encima del nivel del suelo. En el sitio de estudio, el abeto Douglas está presente en masas mixtas y abiertas donde la cubierta del dosel se extiende de 60 a 80 %. La recolección de árboles dominantes y maduros garantiza menor variación en el IAB como el crecimiento del área basal del altiplano de abeto Douglas y se mantienen constantes hasta la edad de 300 años (Poage & Tappeiner, 2002). Se evitaron árboles viejos cercanos a la edad biológica de la especie, ya que tienen tasas de crecimiento bajas y mala relación con la variación del clima (Black, Colbert, & Pederson, 2008).
Mediciones de muestras
Las muestras se secaron al aire y pulieron con papel de lija para un mejor contraste de los anillos de los árboles. El ancho de anillo se midió con un sistema de medición Velmex (Bloomfield, NY, EE.UU.) con una precisión de 0.001 mm (Robinson & Evans, 1980).
Las muestras de núcleos de crecimiento se procesaron siguiendo las técnicas dendrocronológicas (Stokes & Smiley, 1968). La mayoría de las muestras de núcleos de madera incluyeron el anillo central (75%); la edad de los árboles sin anillo central se estimó por regresión de acuerdo con la tendencia de anillos de árboles con edad conocida. El número total de mediciones de anillos fue 2,480. La edad promedio de los árboles muestreados fue de 140 años, variando desde 67 a 203 años.
La intercorrelación de la serie de mediciones de anillos de árboles de los núcleos de madera se evaluó estadísticamente con el software COFECHA (Holmes, 1983). Un modelo exponencial negativo se utilizó para estandarizar la serie de anillos para cada árbol y para desarrollar una serie de ancho de anillos, utilizando el software Arstan (Cook & Holmes, 1984). La curva “spline” para índices de anillos de árboles se ajustó con una resolución de 10 años.
Incremento del área basal (IAB) y datos de precipitación
El incremento de área basal (IAB) se calculó de acuerdo con la siguiente fórmula, donde R es el radio del tallo (cm) y n es el año de la formación del anillo:
Los aumentos en el IAB en etapas maduras tienden a ser estables, siempre y cuando el árbol no se encuentre cerca de los límites de la etapa juvenil o senescencia biológica (Poage & Tappeiner, 2002; Weiner & Thomas, 2001). A pesar de que la edad de los árboles de la muestra varió de 64 a 204 años, su edad está lejos de la senescencia biológica para la especie de estudio, la cual es de aproximadamente 300 años (Villanueva-Diaz et al., 2010). Con el fin de elaborar una serie del IAB representativo para el sitio de estudio, promediamos los valores del IAB en una resolución anual para todas las muestras. La duración máxima de la serie del IAB fue de 203 años (1807 a 2010). El IAB y los anillos del árbol estandarizados con la serie (IAA) se analizaron para comprobar una correlación significativa (P < 0.05). Por otra parte, las series de IAA e IAB estaban relacionadas con la precipitación regional, para lo cual se utilizó el programa DENDROCLIM 2002 (Biondi & Waikul, 2004). Se utilizó la información de las cuatro estaciones meteorológicas más cercanas, según lo informado por la Comisión Nacional del Agua (Instituto Mexicano de Tecnología del Agua [IMTA], 2013). Además, para eliminar el efecto de diferentes edades de árboles, los datos del año natural a la edad cambial se reordenaron y se ajustó una ecuación polinómica de cinco para estimar el IAB esperado según la edad media de los árboles (Briffa & Melvin, 2011) en nuestra muestra.
Análisis espectral
Los ciclos del IAB se identificaron mediante un análisis espectral. La series de tiempo se descompuso en la suma de las ondas de seno y coseno con diferentes amplitudes y duraciones (Jenkins & Watts, 1968; Priestly, 1981):
donde:
xt = |
Datos |
a0 = |
Plazo medio: |
m = |
Número de frecuencias en la descomposición de Fourier |
ak = |
Coeficientes de los cosenos |
wk = |
Frecuencias de Fourier: |
t = |
Tiempo subíndice |
bk = |
Coeficientes de los senos |
Las estadísticas de Kappa de Fisher y Kolmogorov-Smirnov de Bartlett se calcularon para la serie del IAB asegurándose de que la serie y el espectro no fueran “ruido blanco” y fueran estadísticamente significativas. Además, la densidad espectral y las gráficas del periodograma se desarrollaron y estudiaron para determinar la presencia de los ciclos significativos de baja frecuencia. La estimación de la densidad espectral se produjo por suavizamiento del periodograma.
Modelos ARMA y ARIMA de Box-Jenkins
Los modelos ARMA (p, q) desarrollados por Box-Jenkins dependen del segundo orden estacionario; la presencia de los términos autorregresivos p (AR) y parámetros de medias móviles q (MA) ayudan a explicar el comportamiento de la variable de respuesta (Box & Jenkins, 1970).
donde
Zt = |
Desviación media en el tiempo t de series estacionarias igualmente espaciadas |
ϕi y ϕp = |
Términos autorregresivos (ϕp(B) = (1-ϕiBi-…-ϕpBp)) |
B = |
Operador backshift (θq(B) = (1-θjBj-…-θqBq)) |
θj and θq = |
Términos de medias móviles |
ut = |
Variables independientes aleatorias con media cero y varianza (ruido blanco) |
Los modelos estacionarios de segundo orden son la media [E(Zt) =µ], varianza [Var(Zt) = σ2] y covarianza [Cov(Zt,Zs) = γk] sin depender del tiempo. Una extensión a los modelos ARMA es la categoría de medias móviles autorregresivas integradas o el proceso ARIMA con diferencia dth. El término d es el número de veces que la serie debe ser diferenciada para ser estacionaria. La tendencia estocástica se puede eliminar al aplicar un operador de diferencia ∇d:
Donde Xt es la series de tiempo diferenciada d para transformarse en serie estacionaria y Zt es la serie original. La prueba de raíz unitaria aumentada de Dickey-Fuller demuestra que la series de tiempo es estacionaria, poniendo a prueba la hipótesis H0: ρ = 1 y la alternativa H1: ρ < 1. Si la no estacionariedad se asocia con una varianza de variable podemos utilizar una transformación de energía de tipo InZt (Pankrats, 1983). Debe tomarse en cuenta que cuando se necesita eliminar la tendencia y varianza de variable se requiere la transformación de valores reales a logaritmos antes de la diferenciación.
Los modelos ARIMA pueden describir los ciclos para los datos anuales. Es conveniente utilizar un operador de diferencia de ciclo para lograr la estacionariedad del mismo:
Donde Wt es la series de tiempo diferenciada D (duración del ciclo C). Los modelos ARIMA de efectos cíclicos pueden ser propuestos en el siguiente modelo ARIMA general (p, d, q) x (P, D, Q)C:
donde
ϕp (B) = |
Modelo AR no cíclico |
θq (B) = |
Modelo MA no cíclico |
ΦP(BC) = |
Modelo AR cíclico de duración C |
ΘQ(BC) = |
Modelo MA cíclico de duración C |
|
Modelos de diferenciación para compuestos cíclicos y no cíclicos |
En este estudio analizamos la serie del IAB mediante modelos autorregresivos integrados de medias móviles (modelos ARIMA de Box-Jenkins), para encontrar ciclos grandes o periodicidad de crecimiento de árboles.
Estimación de los componentes del modelo ARIMA
La modelización ARIMA implicó la comparación de las gráficas de función de autocorrelación estimada (FAC) y autocorrelación parcial (FACP) con FAC y FACP teórica. El patrón de la decadencia de esas funciones y los desfases con los valores significativos son los componentes para determinar el orden de los parámetros para el modelo ARMA (Box & Jenkins, 1970). La FAC se utilizó para determinar el orden (q) de media móvil (MM), y la FACP, para determinar el orden (p) del modelo autorregresivo. Complementariamente se utilizó el método de correlación canónica (SCAN) más pequeño. Este proceso es útil para identificar las órdenes de un modelo ARMA estacionario y no estacionario. En general este método consiste en la identificación de un patrón rectangular de un cuadro que contiene los valores propios de una matriz de producto del vector Ym,t = (Zt, Zt-1 ,... Zt-m ) donde Zt es la series de tiempo, de tal manera que m = Pmin ,...,Pmax y j = qmin ,...,qmax ; donde m y j identifican el posible orden del proceso MA y AR, respectivamente (Tsay & Tiao, 1984). Finalmente, los parámetros se estimaron mediante mínimos cuadrados condicionales (MCC) (Statistical Analysis System [SAS], 2011).
Estrategia para diagnóstico, control y estimación
Los resultados del análisis espectral se consideraron para evaluar los diferentes componentes cíclicos (C) del proceso ARIMA y generar un modelo apropiado para la comprensión del comportamiento del crecimiento de los árboles. La fiabilidad estadística del modelo se verificó a través de una prueba de Chi-cuadrada (prueba de Ljung-Box) al asegurar que los choques aleatorios y ut fueron estadísticamente independientes (no autocorrelacionados) y “ruido blanco”. La predicción se calculó con la estimación de memoria infinita, también llamada estimación condicional. El término condicional se utiliza porque la predicción se calcula al suponer que los valores desconocidos de la serie de respuesta, antes del inicio de los datos, son iguales a la media de la serie. Se realizó una estimación del IAB para el próximo periodo de 15 años y hasta el año 2090 con un nivel de confianza de 95 %.
Resultados y discusión
Serie de anillos de árboles, IAB y precipitación
Los resultados mostraron que nuestra estrategia para la selección de árboles dominantes con sección del tronco circular es confiable para los propósitos dendrocronológicos. La intensidad de la señal en ARSTAN fue alta con un valor de 0.90, lo que indica la existencia de una respuesta común en todos los árboles muestreados (Cook, 1983, 1987). La intercorrelación entre la serie y la sensibilidad media calculada con COFECHA fue adecuada (P < 0.01), con valores de 0.67 y 0.27, respectivamente. Estos valores indican que las mediciones de anillos muestran una tendencia común e idoneidad para el registro transversal de la serie (Figura 2a). La correlación entre IAA y AIB fue estadísticamente significativa (r = 0.80; P < 0.01) lo que indica que en este caso, las mediciones directas IAB son confiables para describir los cambios en el crecimiento de los árboles y útiles para el análisis de series de tiempo (Figura 2b). IAA y IAB también se correlacionaron significativamente (P < 0.05) con la precipitación para el periodo de enero a julio, con valores de 0.73 y 0.70, respectivamente. Estos resultados también confirman la sensibilidad del abeto Douglas a la variabilidad del clima e idoneidad para utilizar su serie de anillos con el fin de predecir series de crecimiento, como se observó en árboles de P. menziesii que crecen en Durango y Zacatecas, México (González-Elizondo et al., 2005).

Tendencias de crecimiento de árboles
La media de IAB de P. menziesii fue de 54 cm2•año-1 (DE ±12). Este resultado es similar a los 50 cm2•año-1 reportados para los rodales de abeto Douglas que crecen en el oeste de Oregon, donde la edad de los árboles varía de 100 a 300 años, y en el altiplano a (Poage & Tappeiner, 2002). Los resultados muestran una tendencia a la reducción del IAB, que es un resultado inesperado ya que la muestra se componía de árboles sanos y dominantes con edad promedio de 140 años (intervalo de 67-204 años); esto es 150 años menos que la senescencia natural de la especie. Una reducción en el IAB también se ha reportado para otros bosques que sugieren que el estrés de nutrientes e hídrico son factores de tensión (Gomez-Guerrero et al., 2013; Park et al., 2013; Peñuelas, Canadell, & Ogaya, 2011; Silva, Gómez-Guerrero, Doane, & Horwath, 2015). Una fuerte relación entre el diámetro de crecimiento y la humedad también se han observado en P. menziesii en la Sierra Madre Oriental con grandes desviaciones en el crecimiento del diámetro de acuerdo con episodios secos y húmedos extremos (Arreola-Ortiz & Návar-Cháidez, 2010).
Al analizar los resultados, a una resolución de décadas, las reducciones en IAB con respecto a la década anterior fueron 24 y 28 % para los periodos 1890-1899 y 1950-1959, respectivamente. Los periodos favorables resultaron en un aumento del IAB de aproximadamente 16 % por encima del valor medio (Figura 3a). En una base de porcentaje y valores absolutos, el impacto de los periodos favorables es menor que la de los periodos de sequía porque el beneficio de los periodos húmedos en IAB se limita a la capacidad de almacenamiento de agua de los suelos (Castruita, 2014). El periodo de décadas de sequía de los años 1950 y 1990 también se ha reportado en el análisis de flujo de río en México (Návar, 2012) y las evaluaciones de contenido de humedad del suelo utilizando modelos hidrológicos (Návar, 2015).
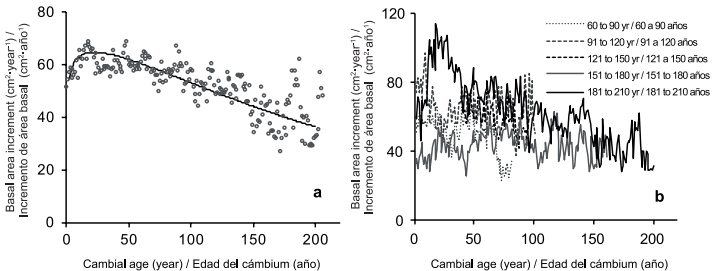
Figura 3 (a) Tendencia del incremento del área basal (IAB) promedio de Pseudotsuga menziesii, de acuerdo con la ecuación de Chapman-Richards P < 0.05 y (b) IAB y edad del cámbium por categorías de edad.
La reducción del IAB en este estudio no se relaciona con la edad de los árboles, debido a que de acuerdo con la edad cambial en Figura 3b, el promedio esperado en el IAB para árboles de 140 años de edad es de 50 cm2•año-1, muy cerca del valor medio de 54 cm2•año-1. Por lo tanto, la tendencia de reducción en el IAB es más probable debido al efecto del clima ya que el IAA y el IAB se correlacionaron estadísticamente, así como la precipitación de enero a julio. Según nuestros resultados, el estrés hídrico por sequías podría ser la razón de las reducciones importantes del IAB en 1893, 1953 y 1996 a valores de 23, 19 y 25 cm2•año-1, respectivamente. Para estos años se han reportado sequías prolongadas; por ejemplo, un periodo seco severo en el año 1950 se reportó en el norte de México (Díaz, Therrell, Stahle, & Cleaveland, 2002) que afectó el sur de Estados Unidos y para mediados de los años 90 también se detectó el mismo problema en árboles de Taxodium mucronatum (Ten.) en el centro de México (Stahle et al., 2009; Stahle et al., 2011).
Las nuevas revisiones de estandarización de mediciones de anillos de árboles sugieren que la selección de árboles dominantes podría dar lugar a una sobrestimación de crecimiento de árboles (Nehrbass Ahles et al., 2014). Se considera que este no es el caso de la muestra de árboles ya que al ordenar los datos de acuerdo con la edad cambial, no se observó una diferencia desproporcionada en el IAB entre las categorías de edad, especialmente después de que la edad del cámbium ha llegado a los 50 años (Figura 3b). Las tendencias decrecientes en el crecimiento del árbol en algunas ocasiones se deben a la inclusión de árboles longevos con tasas de crecimiento bajas (Black et al., 2008); sin embargo, la categoría de mayor edad en este estudio (181-204 años) muestra un IAB esperado de 50 cm2•año-1 a edades cámbium de 150 años, sustentando que la muestra está compuesta por árboles maduros (no viejos) con tasas de crecimiento promedio para esta edad.
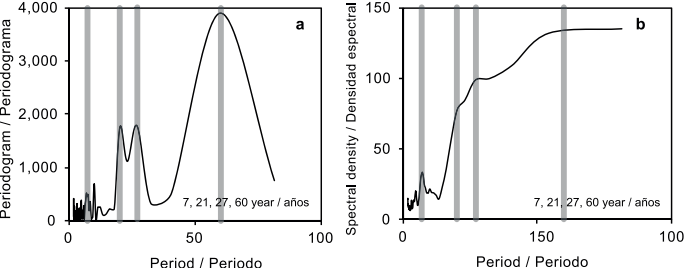
Análisis espectral y periodicidad
Los estadísticos de Kappa de Fisher (P < 0.05) y Kolmogorov-Smirnov de Bartlett fueron significativos (P < 0.0001), lo que indica que la serie y sus espectros no son ruido blanco y el análisis es imparcial (Fuller, 1976). Los resultados indicaron periodicidades de 7, 21, 27 y 60 años (Figura 4). Debido a que se está analizando un proceso biológico, estas periodicidades representan ciclos aproximados; sin embargo, lo que es muy notable es que el último ciclo concuerda con la repetición de sequías en 60 años reportada para el norte de México (Cerano et al., 2011). Aunque es difícil establecer una relación directa con los eventos circulatorios globales, el hecho que merece especial atención es que los ciclos que se encuentran aquí han demostrado ser compatibles con los ciclos del El Niño-Oscilación del Sur (ENSO, por sus siglas en inglés) y Oscilación Decadal del Pacífico (PDO, por sus siglas en inglés) (Hare & Mantua, 2000; Návar & Lizárraga-Mendiola 2013; Trenberth, 1997). Una explicación para esto podría ser la sensibilidad de la especie de estudio en relación con las variaciones de humedad, bajo la influencia de eventos regionales (Griesbauer & Scott, 2010; Návar, 2015). Los resultados coinciden con los ciclos de 7, 21 y 60-años asociados con otros eventos circulatorios globales como ENSO, PDO y AMO (Oscilación multidecadal del Atlántico), respectivamente, según lo propuesto por otros autores (Návar & Lizárraga-Mendiola, 2013; Návar, 2015).
Transformación estacionaria del IAB
Las pruebas de raíz unitaria de Dickey-Fuller y función de autocorrelación (FAC) de la serie original indicaron no estacionariedad (Cuadro 1). La transformación del IAB a logaritmo natural y su primera diferencia (Yt) permite que las series se agrupen en torno a la media con una varianza homogénea. La transformación logarítmica es adecuada para disminuir la heterocedasticidad y se propone la diferencia dth cuando la media de las series cambie con el tiempo (Pankrats, 1983). La FAC de la serie original del IAB del abeto Douglas decae a niveles no significativos en tres años, lo que sugiere que el crecimiento de los tres años anteriores se correlaciona con el crecimiento de árboles y diámetro actual. La transformación y primera diferenciación dieron lugar a una tendencia diferente o FAC, con autocorrelación negativa significativa en el retraso de un año (Figura 5).
Cuadro 1 Pruebas de raíz unitaria aumentada de Dickey-Fuller de la serie original del incremento de área basal en Pseudotsuga menziesii.
| Tipo | Desfases | Rho | Pr < Rho | Tau | Pr < Tau | F | Pr > F |
|---|---|---|---|---|---|---|---|
| 0 | 73.40 | 0.0013 | −4.80 | <0.0001 | 26.25 | 0.001 | |
| Única media | 1 | −49.21 | 0.0013 | −5.46 | <0.0001 | 14.97 | 0.001 |
| 2 | −53.23 | 0.0013 | −5.49 | <0.0002 | 11.58 | 0.001 | |
| 0 | −70.75 | 0.0005 | −6.09 | <0.0001 | 32.51 | 0.001 | |
| Tendencia | 1 | −73.06 | 0.0005 | −7.24 | <0.0001 | 18.70 | 0.001 |
| 2 | −89.03 | 0.0005 | −8.04 | <0.0001 | 15.16 | 0.001 |

Modelos ARIMA de estimación e identificación
El patrón de la decadencia para FAC y FACp del IAB transformado propone un modelo MA(1). Las características del modelo se definieron por el aumento en el desfase 1, que posteriormente se reduce a 0 (Figura 5). El hecho de que el pico fue negativo con tendencia exponencial negativa (FACp) indica que θ > 0 (FAC) (Pankrats, 1983). Este resultado fue consistente con el método de correlación canónica (SCAN) más pequeño que propone un modelo ARIMA (0,1,1). Después de considerar FAC y FACp, el modelo seleccionado fue ARIMA (0,1,1). Este modelo tiene un parámetro medio móvil (q) que implica perturbaciones aleatorias, lo que es diferente de los modelos AR (1) o ARMA (1,1) que son propuestos frecuentemente para la serie de ancho de anillo (Meko, 1981). El modelo seleccionado se evaluó con todas las periodicidades encontradas (7, 21, 27 y 60) y sin ellas para evaluar su significancia. Tomando en cuenta los valores de t y P, todos los modelos fueron significativos (C = 7, 21, 27 y 60). El término promedio móvil (θ) para todos los casos fue positivo con valores de 0.46 a 0.76, mientras que el término de media móvil cíclica (Θ) varió de 0.45 a 0.83 (Cuadro 2).
Cuadro 2 Ajuste de los modelos de Pseudotsuga menziesii.
| Model | Parametros* | Error estándar | Valor T | Pr > |t| | Estimación de la varianza | Standard error of the estimation | CIA** | CIB** | ||
|---|---|---|---|---|---|---|---|---|---|---|
| θ 1 | ϕ 1 | ΦP/ΘQ | ||||||||
| ARIMA (0,1,1) | 0.503 | -- | -- | 0.068 | 7.38 | <0.0001 | 0.043 | 0.207 | -49.23 | -46.14 |
| ARIMA (0,1,1) (0,1,1)7 | 0.458 | -- | -- | 0.071 | 6.38 | <0.0001 | 0.050 | 0.225 | -20.01 | -13.92 |
| -- | 0.832 | 0.047 | 17.54 | <0.0001 | -- | -- | -- | -- | ||
| ARIMA (0,1,1) (0,1,1)21 | 0.647 | -- | -- | 0.065 | 9.97 | <0.0001 | 0.054 | 0.232 | -8.81 | -2.91 |
| -- | 0.645 | 0.075 | 8.53 | <0.0001 | -- | -- | -- | -- | ||
| ARIMA (0,1,1) (0,1,1)27 | 0.492 | -- | -- | 0.076 | 6.49 | <0.0001 | 0.063 | 0.251 | 11.75 | 17.56 |
| -- | 0.780 | 0.074 | 10.46 | <0.0001 | -- | -- | -- | -- | ||
| ARIMA (0,1,1) (0,1,1)60 | 0.764 | -- | -- | 0.065 | 11.59 | <0.0001 | 0.060 | 0.246 | 5.72 | 10.97 |
| -- | 0.448 | 0.166 | 2.70 | <0.0001 | -- | -- | -- | -- | ||
*θ1 es el término de media móvil no cíclica, ϕ1 es el modelo autoregresivo no cíclico, ΦP es el modelo autoregresivo cíclico de duración C, y ΘQ es el modelo de media móvil cíclico de duración C.
** Criterio de información de Akaike (AIC) y criterio de información bayesiano (CIB)
Para propósitos de predicción, el modelo seleccionado fue ARIMA (0,1,1) × (0,1,1)60. Dos razones apoyan esta selección: 1) En primer lugar, su fuerza estadística se refleja en la significancia de sus términos, no hay residuos correlacionados, criterio de información de Akaike (CIA) y criterio de información bayesiano (CIB) (valores más bajos), y el ajuste fiable de la fechas; 2) En segundo lugar, la periodicidad de 60 años se ha relacionado con patrones circulatorios globales por otros autores para la especie estudiada (Cerano et al., 2011) y tiene un impacto importante en el crecimiento de los árboles en el área de estudio.
Predicción del IAB
Con el mejor modelo ARIMA estadístico y climático [ARIMA (0,1,1) x (0,1,1)60], se realizó una predicción para el IAB en las próximas ocho décadas utilizando el software SAS 9.3 (SAS, 2011) de acuerdo con las ecuaciones:
Utilizando coeficientes del Cuadro 2:
Este modelo considera al IAB con base en las funciones de las perturbaciones aleatorias, restando 76 % del mismo componente del año anterior, 45 % de los ocurridos hace 60 años y la adición de 34 % de los ocurridos 61 años atrás. A largo plazo, la predicción indica una reducción de 72 % en el IAB con un promedio de 15 cm2•año-1, y una recuperación hacia el año 2025. No obstante, el crecimiento proyectado en 2025 para esta especie está por debajo de la media histórica (54 cm2•año-1), lo que sugiere una reducción neta de crecimiento para los próximos 75 años (Figura 6); sin embargo, la amplitud de los intervalos de confianza limita la interpretación de varias décadas.
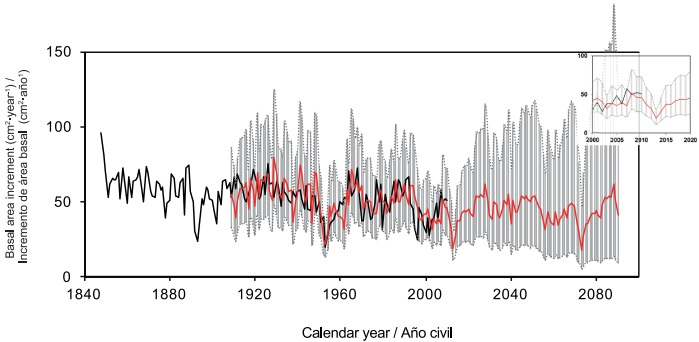
Figura 6 Predicción extendida del incremento del área basal de Pseudotsuga menziesii, a partir del modelo ARIMA. La línea de color negro indica mediciones reales, y la línea roja, las estimaciones. El área gris define el intervalo de confianza al 95 %.
Con base en los resultados se asume que la frecuencia de otros factores estresantes de los bosques, como plagas e incendios se mantienen constantes, debido a ello, la salud del bosque estudiado podría estar en riesgo con efectos negativos concomitantes de otros factores. La prevalencia de bosques saludables requiere un estudio de línea base para el crecimiento de árboles como el llevado a cabo.
Las tendencias de crecimiento de los árboles, encontradas en este estudio, no muestran señales de efectos combinados y benéficos en el aumento de CO2 en la atmósfera y temperatura como Soulé and Knapp (2015) lo han reportado. Nuestra muestra de árbol mostró cuatro fases de crecimiento máximo, que fueron disminuyendo con el tiempo (aproximadamente, 1860-1880, 1920-1940, 1960-1980, y 1980-2000). La principal implicación de estos resultados son los impactos esperados de la variación del clima en el futuro a corto plazo. La predicción del modelo propuesto no está considerando la variación del clima de manera implícita; la tendencia de reducción de crecimiento en árboles puede ser una indicación de una respuesta generalizada de los bosques en otras partes del mundo (Silva, Anand, & Leithead, 2010).
Creemos que los datos para IAB, IAA y precipitación fueron altamente correlacionados porque utilizamos el IAB directo para el año de la formación de los anillos. Independientemente de la edad de los árboles, todos los anillos formados en las mismas condiciones ambientales reflejan el fuerte efecto de la precipitación. Otra razón es que las tasas de crecimiento de los árboles provenían de un amplio abanico de edades. De acuerdo con algunos autores, el análisis mediante edad cambial indicó que el IAB del abeto Douglas tiende a ser menos sensible al clima a medida que el árbol envejece (Black et al., 2008), pero nuestros resultados indican que una muestra compuesta por árboles maduros y dominantes de rodales abiertos es fiable para detectar una señalización común de los árboles. Uno de los mejores métodos para seleccionar una muestra de árboles para fines dendrocronológicos y estimación de crecimiento de árboles es la selección al azar que incluyen diferentes edades y diámetros (Nehrbass‐Ahles et al., 2014). En ese sentido, nuestro método de selección para árboles altos y saludables en masas mixtas y abiertas de abeto Douglas conduce a una muestra heterogénea de edad y diámetro (40-104 cm) que funcionó bien para los propósitos del estudio.
El método utilizado en este estudio muestra potencial para usarse con otras especies forestales con la finalidad de investigar los efectos del cambio climático a escala geográfica. Las tendencias de crecimiento de árboles que encontramos son similares a las reportadas por otros autores, las cuales indicaron que los eventos de sequías extremas como las de 1893 y 1952 fueron geográficamente importantes en el norte de México y suroeste de EE.UU. (Cerano et al., 2011; Stahle et al., 2009).
Aunque el presente estudio no ha sido probado a escala regional, el método podría tener cierto potencial para correlacionar series de IAB con índices de vigor de plantas como el NDVI (índice de vegetación de diferencia normalizada) propuesto por otros autores (Berner, Beck, Bunn, & Goetz, 2013). Las series de IAB deben considerarse como una variable sustitutiva para estudiar los efectos del cambio climático, en particular para las especies forestales sensibles. De hecho, el nuevo proceso de estandarización en la dendrocronología propone estandarizar en términos de IAB estable observado en la edad madura de las especies forestales (Biondi & Qeadan, 2008).
Conclusiones
En este estudio se encontró fuerte correlación entre los índices de anillos de crecimiento, crecimiento de área basal de la especie y precipitación. El análisis de series de tiempo de IAB en abeto Douglas de la Sierra Madre Occidental permitió la identificación de la periodicidad en el crecimiento del árbol, que coincide con los eventos atmosféricos globales como los ciclos de ENSO. La predicción de IAB indica la disminución del crecimiento de los árboles dentro de la presente década, a pesar de que cierto grado de recuperación se alcanzará en el año 2025. No obstante, las proyecciones de crecimiento de árboles a largo plazo no serán tan altas como las observadas en la década de 1960 (60 cm2•año-1). La reducción del IAB en este estudio no se relaciona con la edad de los árboles, ya que también se analizaron las tendencias de acuerdo con la edad del cámbium. El uso de análisis de series temporales del IAB de árboles dominantes y maduros puede ser útil para predecir cambios en el crecimiento del árbol y proponer las prácticas de manejo más adecuadas.
Agradecimientos
Se agradece a la fundación FORD, al Consejo Nacional de Ciencia y Tecnología (CONACYT) y a la Universidad de California por el apoyo financiero proporcionado, a través del programa UC-MEXUS (Proyecto CN-13-546), para llevar a cabo este estudio.
REFERENCIAS
Arreola-Ortiz, M. R., & Návar-Cháidez, J. J. (2010). Análisis de sequías y productividad con cronologías de Pseudotsuga menziesii Rob. & Fern., y su asociación con el niño en el nordeste de México. Investigaciones geográficas, 7-20. Retrieved from http://www.scielo.org.mx/pdf/igeo/n71/n71a2.pdf [ Links ]
Berner, L. T., Beck, P. S. A., Bunn, A. G., & Goetz, S. J. (2013). Plant response to climate change along the forest-tundra ecotone in northeastern Siberia. Global Change Biology, 19(11), 3449-3462. doi: 10.1111/gcb.12304 [ Links ]
Biondi, F., & Qeadan, F. (2008). A theory-driven approach to tree-ring standardization: Defining the biological trend from expected basal area increment. Tree-Ring Research, 64(2), 81-96. doi: 10.3959/2008-6.1 [ Links ]
Biondi, F., & Waikul, K. (2004). DENDROCLIM2002: A C++ program for statistical calibration of climate signals in tree-ring chronologies. Computers & Geosciences, 30(3), 303-311. doi: 10.1016/j.cageo.2003.11.004 [ Links ]
Black, B. A., Colbert, J. J., & Pederson, N. (2008). Relationships between radial growth rates and lifespan within North American tree species. Ecoscience, 15(3), 349-357. doi: 10.2980/15-3-3149 [ Links ]
Box, G. E. P., & Jenkins, G. M. (1970). Time series analisys, forecasting and control. San Francisco, USA: Holden-Day. [ Links ]
Briffa, K. R., & Melvin, T. M. (2011). A closer look at regional curve standardization of tree-ring records: Justification of the need, a warning of some pitfalls, and suggested improvements in its application. In K. M., Hughes, W. T., Swetnam & F. H.,Diaz (Eds.), Dendroclimatology: Progress and Prospects (pp. 113-145). Dordrecht, UK: Springer Netherlands. doi: 10.1007/978-1-4020-5725-0_5 [ Links ]
Castruita, E. L. U. (2014). Variabilidad climática, eficiencia de uso de agua intrínseca y crecimiento del área basal en Bosques del norte de México. (Tesis doctoral, Colegio de Postgraduados, México. Retrieved from http://www.biblio.colpos.mx:8080/jspui/bitstream/handle/10521/2384/Castruita_Esparza_LU_DC_Forestal_2014.pdf?sequence=1 [ Links ]
Cerano, P. J., Villanueva, D. J., Valdéz, C. R., Méndez, G. J., & Constante, G. V. (2011). Sequías reconstruidas en los últimos 600 años para el noreste de México. Revista Mexicana de Ciencias Agrícolas, 2, 235-249. Retrieved from http://www.redalyc.org/articulo.oa?id=263121431006 [ Links ]
Cook, E. R. (1983). A time series analysis approach to tree-ring standardization. Ph. D. dissertation. University of Arizona, USA. [ Links ]
Cook, E. R. (1987). The decomposition of tree-ring series for environmental studies. Tree-Ring Bulletin, 47, 37-59. Retrieved from http://arizona.openrepository.com/arizona/handle/10150/261788 [ Links ]
Cook, E. R., & Holmes, R. L. (1984). Program ARSTAN and users manual. Tucson, AZ, USA: Laboratory of Tree-Ring Research, University of Arizona. [ Links ]
Díaz, S. C., Therrell, M. D., Stahle, D. W., & Cleaveland, M. K. (2002). Chihuahua (Mexico) winter-spring precipitation reconstructed from tree-rings, 1647- 1992. Climate Research, 22(3), 237-244. doi: 10.3354/cr022237 [ Links ]
Esper, J., Benz, M., & Pederson, N. (2012). Influence of wood harvest on tree-ring time-series of Picea abies in a temperate forest. Forest Ecology and Management, 284, 86-92. doi: 10.1016/j.foreco.2012.07.047 [ Links ]
Fuller, W. A. (1976). Introduction to statical time series. New York, USA: John Wiley & Sons, Inc. [ Links ]
García-Arévalo, A. (2008). Vegetación y flora de un bosque relictual de Picea chihuahuana Martínez del norte de México. Polibotánica, 25, 45-68. Retrieved from http://www.scielo.org.mx/pdf/polib/n25/n25a5.pdf [ Links ]
Gomez-Guerrero, A., Silva, L. C. R., Barrera-Reyes, M., Kishchuk, B., Velazquez-Martinez, A., Martinez- Trinidad, T., Horwath, W. R. (2013). Growth decline and divergent tree ring isotopic composition (13C and 18O) contradict predictions of CO2 stimulation in high altitudinal forests. Global Change Biology, 19(6), 1748-1758. doi: 10.1111/gcb.12170 [ Links ]
González-Elizondo, M., Jurado, E., Návar, J., González- Elizondo, M. S., Villanueva, J., Aguirre, O., & Jiménez, J. (2005). Tree-rings and climate relationships for douglas-fir chronologies from the sierra madre occidental, mexico: A 1681-2001 rain reconstruction. Forest Ecology and Management, 213, 39- 53. doi: 10.1016/j.foreco.2005.03.012 [ Links ]
González, E. M. S., González, M. E., & Márquez, M. A. L. (2007). Vegetación y ecorregiones de Durango. México: Plaza y Valdés. [ Links ]
Griesbauer, H., & Scott, G. D. (2010). Assessing the climatic sensitivity of Douglas-fir at its northern range margins in British Columbia, Canada. Trees, 24(2), 375- 389. doi: 10.1007/s00468-009-0407-z [ Links ]
Gugger, P. F., González-Rodríguez, A., Rodríguez-Correa, H., Sugita, S., & Cavender-Bares, J. (2011). Southward Pleistocene migration of Douglas-fir into Mexico: Phylogeography, ecological niche modeling, and conservation of ‘rear edge’ populations. New Phytologist, 189(4), 1185-1199. doi: 10.1111/j.1469-8137.2010.03559.x [ Links ]
Hansen, J., Sato, M., & Ruedy, R. (2012). Perception of climate change. Proceedings of the National Academy of Sciences, 109(37), E2415-E2423. doi: 10.1073/pnas.1205276109 [ Links ]
Hare, S. R., & Mantua, N. J. (2000). Empirical evidence for North Pacific regime shifts in 1977 and 1989. Progress in oceanography, 47(2), 103-145. doi: 10.1016/S0079-6611(00)00033-1 [ Links ]
Holmes, R. L. (1983). Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 43, 69-78. [ Links ]
Instituto Mexicano de Tecnología del Agua (IMTA). (2013). Extractor Rápido de Información Climatológica III (ERIC) version 3.2. Jiutepec, Morelos, México: Author. [ Links ]
Jenkins, G. M., & Watts, D. G. (1968). Spectral analysis and its applications. San Francisco, USA: Holden-Day . [ Links ]
Matisons, R., Elferts, D., & Brūmelis, G. (2013). Pointer years in tree-ring width and earlywood-vessel area time series of Quercus robur-Relation with climate factors near its northern distribution limit. Dendrochronologia, 31(2), 129-139. doi: 10.1016/j.dendro.2012.10.001 [ Links ]
Meko, D. M. (1981). Application of Box-Jenkins methods of time series analysis to the reconstruction of drought from tree rings. USA: University of Arizona. [ Links ]
Monserud, R. A., & Marshall, J. D. (2001). Time-series analysis of δ13C from tree rings. Time trends and autocorrelation. Tree Physiology, 21, 1087-1102. doi: 10.1093/treephys/21.15.1087 [ Links ]
Návar, J. (2012). Modeling annual discharge of six Mexico’s northern rivers. Revista Ambiente & Água, 7, 36-50. doi: 10.4136/ambi-agua.705 [ Links ]
Návar, J. (2015). Hydro-climatic variability and perturbations in mexico’s north-western temperate forests. Ecohydrology, 8, 1065-1072. doi: 10.1002/eco.1564 [ Links ]
Návar, J., & Lizárraga-Mendiola, L. (2013). Hydro-climatic variability and forest fires in Mexico’s northern temperate forests. Geofísica internacional, 52, 5-20. doi: 10.1016/S0016-7169(13)71458-2 [ Links ]
Nehrbass-Ahles, C., Babst, F., Klesse, S., Nötzli, M., Bouriaud, O., Neukom, R., Frank, D. (2014). The influence of sampling design on tree-ring based quantification of forest growth. Global Change Biology, 20(9), 2867-2885. doi: 10.1111/gcb.12599 [ Links ]
Pankrats, A. (1983). Forecasting with univariate Box-Jenkins models. Concepts and cases. USA: John Wiley and Sons. [ Links ]
Park, W. A., Allen, C. D., Macalady, A. K., Griffin, D., Woodhouse, C. A., Meko, D. M., McDowell, N. G. (2013). Temperature as a potent driver of regional forest drought stress and tree mortality. Nature Clim. Change, 3(3), 292-297. doi: 10.1038/nclimate1693 [ Links ]
Peñuelas, J., Canadell, J. G., & Ogaya, R. (2011). Increased water-use efficiency during the 20th century did not translate into enhanced tree growth. Global Ecology and Biogeography, 20(4), 597-608. doi: 10.1111/j.1466-8238.2010.00608.x [ Links ]
Poage, N. J., & Tappeiner, J. C. (2002). Long-term patterns of diameter and basal area growth of old-growth Douglas-fir trees in western Oregon. Canadian Journal of Forest Research, 32(7), 1232-1243. doi: 10.1139/x02-045 [ Links ]
Priestly, M. B. (1981). Spectral analysis and time series. New York, USA: Academic Press Inc. [ Links ]
Robinson, W. J., & Evans, R. (1980). A microcomputer-based tree-ring measuring system. Tree-Ring Bulletin, 40, 59- 64. Retrieved from http://arizona.openrepository.com/arizona/handle/10150/260443 [ Links ]
Roden, J. S., Johnstone, J. A., & Dawson, T. E. (2011). Regional and watershed-scale coherence in the stable-oxygen and carbon isotope ratio time series in tree rings of Coast Redwood (Sequoia sempervirens). Tree-Ring Research, 67(2), 71-86. doi: 10.3959/2010-4.1 [ Links ]
Rubino, D. L., & McCarthy, B. C. (2000). Dendroclimatological analysis of white oak (Quercus alba L., Fagaceae) from an old-growth forest of Southeastern Ohio, USA. Journal of the Torrey Botanical Society, 127(3), 240-250. doi: 10.2307/3088761 [ Links ]
Secretaría de Medio Ambiente y Recursos Naturales (2010). Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Protección ambiental, especies nativas de flora y fauna silvestres de México, categorías de riesgo y especificaciones para su inclusión, exclusión o cambio, y lista de especies en riesgo. México: Diario Oficial de la Federación. [ Links ]
Silva, L. C. R., Anand, M., & Leithead, M. D. (2010). Recent widespread tree growth decline despite increasing atmospheric CO2. PLoS ONE, 5(7), e11543. doi: 10.1371/journal.pone.0011543 [ Links ]
Silva, L. C. R., Gómez-Guerrero, A., Doane, T. A., & Horwath, W. R. (2015). Isotopic and nutritional evidence for species- and site-specific responses to N deposition and elevated CO2 in temperate forests. Journal of Geophysical Research: Biogeosciences, 120(6), 1110-1123. doi: 10.1002/2014JG002865 [ Links ]
Skovsgaard, J. P., & Vanclay, J. K. (2008). Forest site productivity: A review of the evolution of dendrometric concepts for even-aged stands. Forestry, 81(1), 13-31. doi: 10.1093/forestry/cpm041 [ Links ]
Soulé, P. T., & Knapp, P. A. (2015). Analyses of intrinsic water-use efficiency indicate performance differences of ponderosa pine and douglas-fir in response to CO2 enrichment. Journal of Biogeography, 42,144-155. doi: 10.1111/jbi.12408 [ Links ]
Stahle, D. W., Cook, E. R., Diaz, J. V., Fye, F. K., Burnette, D. J., Griffin, D., Heim, R. R. (2009). Early 21st- Century drought in Mexico. Eos, Transactions American Geophysical Union, 90(11), 89-90. doi: 10.1029/2009EO110001 [ Links ]
Stahle, D. W., Diaz, J. V., Burnette, D. J., Paredes, J., Heim, R., Fye, F. K., Stahle, D. K. (2011). Major Mesoamerican droughts of the past millennium. Geophysical Research Letters, 38(5), 1-4. doi: 10.1029/2010GL046472 [ Links ]
Statistical Analysis System (SAS). (2011). SAS/ETS 9.3 User’s Guide. Cary, NC, USA: SAS Institute Inc. [ Links ]
Stokes, M. A., & Smiley, T. L. (1968). An introduction to tree-ring dating, Chicago, USA: University of Arizona Press. [ Links ]
Swetnam, T. W., & Lynch, A. M. (1993). Multicentury, regional-scale patterns of western spruce budworms outbreaks. Ecological Monographis, 63(4), 399-424. doi: 10.2307/2937153 [ Links ]
Trenberth, K. E. (1997). The definition of el nino. Bulletin of the American Meteorological Society, 78(12), 2771- 2777. Retrieved from http://www.cgd.ucar.edu/staff/trenbert/trenberth.papers/i1520-0477-078-12-2771.pdf [ Links ]
Tsay, R. S., & Tiao, G. C. (1984). Consistent estimates of autoregressive parameters and extended sample autocorrelation function for stationary and nonstationary ARMA models. Journal of the American Statistical Association, 79(385), 84-96. doi: 10.2307/2288340 [ Links ]
Villanueva-Diaz, J., Cerano-Paredes, J., Stahle, D. W., Constante-García, V., Vázquez-Salem, L., Estrada-Ávalos, J., & Benavides-Solorio, J. D. (2010). Árboles longevos de México. Revista Mexicana de Ciencias Forestales, 1(2), 1-23. Retrieved from http://www.redalyc.org/articulo.oa?id=63438955002 [ Links ]
Weiner, J., & Thomas, S. C. (2001). The nature of tree growth and the “age-related decline in forest productivity”. Oikos, 94(2), 374-376. doi: 10.1034/j.1600-0706.2001.940219.x [ Links ]
Recibido: 21 de Octubre de 2015; Aprobado: 20 de Julio de 2016
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
