










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.22 no.3 Chapingo sep./dic. 2016
https://doi.org/10.5154/r.rchscfa.2015.04.016
Articulo Cientifico
Efectos del sitio y la condición de agostadero en la densidad de hormigueros de escamoles (Liometopum apiculatum Mayr)
1Universidad Autónoma de San Luis Potosí (UASLP), Programa Multidisciplinario de Posgrado en Ciencias Ambientales. Av. Manual Nava núm. 201, Zona Universitaria. C. P. 78290. San Luis Potosí, México.
2 Universidad Autónoma de San Luis Potosí (UASLP), Instituto de Investigación de Zonas Desérticas. Altair núm. 200. Fraccionamiento del Llano. C. P. 78377. San Luis Potosí, México.
3 Universidad Autónoma de San Luis Potosí (UASLP), Coordinación Académica Región Altiplano Oeste. Carretera Santo Domingo km 1.0. C. P. 78600. Salinas de Hidalgo, San Luis Potosí, México.
El efecto de la condición de agostadero en la densidad de hormigueros de la hormiga de escamoles (Liometopum apiculatum Mayr) se evaluó en tres sitios diferentes (favorable, regular y desfavorable), en un espacio dividido en dos patrones contrastantes de aprovechamiento y régimen de tenencia comunal (ejido) y privada (rancho) en Charcas, San Luis Potosí, México. La vegetación se midió utilizando el método de cuadrantes centrados en un punto, espaciados sobre transectos; en cada cuadrante se midió el individuo más próximo al punto correspondiente a cada estrato de la vegetación y se registró la distancia al hormiguero más cercano (si existía). Las especies se agruparon de acuerdo con su valor forrajero (deseables, menos deseables e indeseables) y, mediante estimaciones de densidad y biomasa volumétrica, los grupos se contrastaron con un análisis tabular y estadístico descriptivo. La superficie del suelo se evaluó con el método de líneas de intercepción en la proporción de cobertura basal viva y muerta (mantillo), suelo desnudo, rocas, caminos de hormigas y heces. La densidad más alta de los hormigueros (14•ha-1) se registró en el sitio favorable del rancho (mejor condición de agostadero). Con base en la información registrada se encontraron diferencias en la densidad de hormigueros entre sitios de agostaderos contiguos, lo cual a su vez es alterado severamente por la condición de los mismos.
Palabras clave: Densidad de hormigueros; tenencia de la tierra; estado de la vegetación y del suelo; altiplanicie de San Luis Potosí.
The aim of this work was to evaluate the effect of rangeland condition on the nest density of escamoles ant (Liometopum apiculatum) in three different range sites, all Twithin a single (escamoles) collecting area divided into two contrasting use and property regimes, communal and private, in the Municipality of Charcas, San Luis Potosi, Mexico. In order to measure the vegetation, a point-centered quarter method along transects was used; in every quadrant the distance from the point to the closest plant was measured for each vegetation stratum, while the distance to the closest nest (if existing) was also recorded. The species recorded were grouped according to their forage value as desirable, less desirable and undesirable, and using their density and volumetric biomass estimations, these groups were compared through a tabular and descriptive statistical analysis. The soil surface state was evaluated with the line intercept method, in its proportion of live and dead (mulch) basal cover, bare soil, stones, ant trails and feces. The highest density of ant nests (14•ha-1) was recorded in the favorable site of the ranch (best rangeland condition). Marked differences were found in nest density between adjacent rangeland sites, which in turn were severely altered by the condition of these rangelands.
Keywords: Ant nest density; land tenure; vegetation and soil state; San Luis Potosí highland.
Introducción
Un sitio de agostadero es una unidad de suelo con características físicas y bióticas homogéneas que responde de manera similar a un patrón de uso. Con frecuencia, diferentes estados o condiciones de agostadero se producen dentro de un sitio, ya sea espacial o temporalmente, los cuales según Heady y Child (1994) son los estados actuales de la vegetación con relación a la condición potencial del sitio; es decir, con el estado maduro y relativamente estable de la vegetación en ese sitio. El recurso más importante en un agostadero es su cubierta vegetal; por lo tanto, la determinación de su condición proporciona información detallada sobre cómo se ha manejado y las acciones que deben tomarse para un uso óptimo (Heady & Child, 1994; Holechek, Pieper, & Herbel, 2011; Quirk, 2002).
Los cambios de condición producen alteraciones concomitantes en los sistemas de suelo-agua-vegetación (Thurow, 2000). Los ecosistemas de zonas secas presentan características únicas que surgen por la aridez; en estas comunidades hay menor tolerancia al estrés impuesto por los diversos usos, por lo que tardan más tiempo en recuperarse. El deterioro de las zonas secas se debe principalmente a que los usuarios aumentan la demanda de recursos y, a la larga, los degradan al tratar de mejorar sus condiciones de vida (Griffin, 2002).
El mismo problema puede estar ocurriendo con relación a la recolección de pupas (escamoles) de la hormiga Liometopum apiculatum Mayr en el altiplano del estado de San Luis Potosí, donde las actividades económicas primarias predominan. La degradación puede ocurrir sobre todo en los agostaderos de uso común (ejidos) en el municipio de Charcas, donde la recolección de escamoles se ha expandido rápidamente en la última década. Los escamoles son altamente demandados por ser considerados un manjar, pues desde tiempos prehispánicos han sido consumidos por muchos grupos étnicos, y en la actualidad se venden en mercados y restaurantes (Ramos-Elorduy, 2006). Más detalles sobre la biología de esta especie y su uso se han publicado por Lara-Juárez, Aguirre-Rivera, Castillo-Lara, y Reyes-Agüero (2015). Los recolectores locales carecen de los conocimientos tradicionales para la recolecta de escamoles, como sucede en otros estados como Hidalgo y Tlaxcala, e ignoran la relación entre el insecto y su entorno, en términos de alimentación, descanso, refugio del sol o reproducción y construcción de hormigueros; la recolección parece excesiva y descuidada, lo que junto con el exceso de pastoreo puede ser la causa del deterioro rápido del recurso (Lara-Juárez et al., 2015).
El objetivo de este estudio fue relacionar la estructura del agostadero, mediante el efecto de sitio y condición, con la densidad de hormigueros de la especie L. apiculatum en un área de recolección de escamoles. Con este fin, la estructura de la vegetación, el estado de la superficie del suelo y el número de hormigueros activos asociados se evaluaron en un área categorizada de acuerdo con su historia de uso (ejido y rancho privado), tomando en cuenta tres variantes fisonómicas de vegetación (sitios probables) que se extienden sobre ambos tipos de tenencia de la tierra.
Materiales y métodos
Área de estudio
El área de estudio comprende la zona de recolecta de escamoles, que incluye ejidos y ranchos ubicados en la parte noreste del municipio de Charcas, en el norte del estado de San Luis Potosí (29.5 km de la ciudad de Charcas hasta el área de estudio, Figura 1), donde tal actividad económica comenzó recientemente y actualmente se lleva a cabo de forma activa. La litología superficial de la zona es heterogénea, lo que a su vez da lugar a agostaderos de matorrales dominados por especies asociadas a sustratos ígneos o sedimentarios. La temperatura media ponderada y precipitación registrada en las estaciones meteorológicas más cercanas (Laguna Seca, 38 años; Charcas, 54 años) se muestran en la Figura 2 (Instituto Nacional de Estadística, Geografía e Informática [INEGI], 2002).
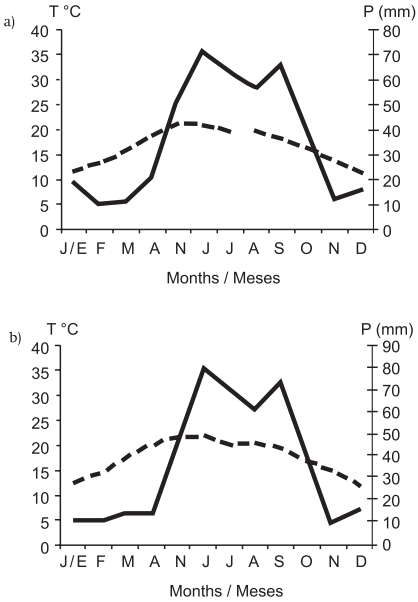
Figura 2 Distribución de la temperatura (---) y la precipitación (—) en la región del área de estudio en San Luis Potosí (tomado de García, 2004). a) Laguna Seca (2,030 m, 16.8 ºC, 439.2 mm), b) Charcas (2,021 m, 17.6 ºC, 446.5 mm).
De acuerdo con el enfoque de Bolaños y Aguirre (2000) se hicieron visitas de reconocimiento en la zona y, con base en mapas temáticos y ortofotos, se localizó un área de geomorfología relativamente homogénea (paisaje), que incluye ranchos y ejidos (Figura 3), lo que hizo posible tener dos zonas contiguas con condición de agostadero e historia de uso contrastante (Figura 4). Posteriormente se identificaron diferencias edáficas y de fisionomía de la vegetación; estas diferencias fueron suficientes para suponer que correspondían a tres sitios de agostadero, mostrando diferencias notables en términos de densidad de hormigueros. Los sitios fueron considerados como sitios favorables, regulares y desfavorables para el establecimiento de hormigueros. Por lo tanto, el área de muestreo incluyó una porción del potrero Cerro Blanco en el ejido Francisco I. Madero, así como parte del potrero Siete Vueltas perteneciente al rancho Laguna Seca.
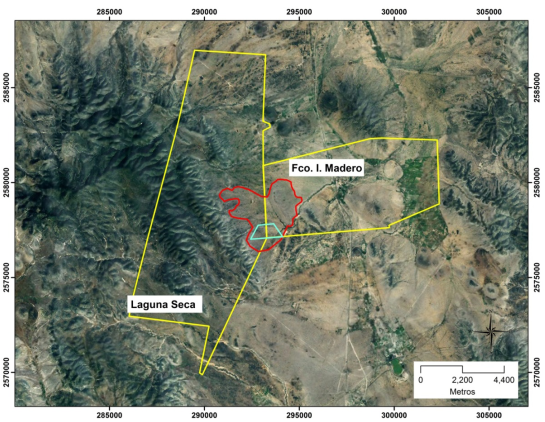
Figura 3 Ubicación de la zona de estudio, rancho Laguna Seca (privado) y ejido Francisco I. Madero (tierra comunal), Charcas, San Luis Potosí (en amarillo, límite de la propiedad; en rojo, límite del paisaje; en azul, área de muestreo)

Figura 4 Límite entre la fracción de Cerro Blanco del ejido Francisco I. Madero (izquierda) y el potrero Siete Vueltas del rancho Laguna Seca (derecha). a) La pérdida de suelo en las tierras comunales ha formado un escalón sobre la línea de la cerca. b) En las tierras comunales se observa menor densidad de vegetación y mayor cantidad de estiércol. c) Aproximación a la diferencia en el nivel del suelo, alrededor de 10 a 12 cm, generada por el sobrepastoreo.
Método de muestreo de la vegetación
Los estudios de campo se llevaron a cabo entre octubre y diciembre del año 2011, durante una sequía de más de dos años. Con el fin de estimar los atributos estructurales de la vegetación y la densidad de hormigueros, se utilizó el método de cuadrantes centrados en un punto a lo largo de transectos, con ajustes para vegetación con varios estratos (Brower, Zar, & von Ende, 1998).
Los transectos se definieron en las imágenes para cada uno de los tres sitios de agostadero, perpendiculares a la cerca que separa el rancho del ejido, y espaciados al menos 100 m entre sí para evitar la superposición.
Como las áreas eran distintas, la cantidad y longitud de transectos fue diferente para cada sitio, pero igual para cada condición. El número de puntos de muestreo fue el mismo en ambas condiciones y se distribuyeron a lo largo de transectos y geotransectos situados en intervalos de 100 m para evitar registrar el mismo hormiguero desde puntos adyacentes, ya que el radio de actividad de las hormigas en torno a su nido fue de aproximadamente 45 a 50 m; los puntos de muestreo se localizaron por lo menos a 50 m de distancia de la cerca (Figura 5). Antes de iniciar las mediciones de campo, las plantas presentes se enumeraron y clasificaron en cuatro estratos de acuerdo con su forma de vida (herbáceas, arbusto bajo, alto y arborescente); posteriormente, en cada punto de muestreo se tomaron, de forma secuencial, mediciones de las especies pertenecientes a cada estrato. Además, se recogieron muestras de especímenes de herbario para corroborar y respaldar las identificaciones de campo, y se depositaron en el herbario Isidro Palacios del Instituto de Investigación de Zonas Desérticas de la Universidad Autónoma de San Luis Potosí. Con base en el enfoque de Vallentine (1990) y Bolaños y Aguirre (2000), las especies vegetales se registraron y clasificaron como deseables, menos deseables e indeseables, de acuerdo con su valor forrajero y reacción sucesional al pastoreo.

Atributos de plantas y hormigueros
En una secuencia por estrato, en cada uno de los cuatro cuadrantes de los 52 puntos de muestra, se registró la distancia desde el punto hasta el hormiguero más cercano, así como la planta más cercana, obteniendo su altura, diámetros mayor y menor de su área basal y dosel. La biomasa volumétrica se estimó indirectamente con la fórmula del volumen de cono truncado, a partir de los datos siguientes:
V = 1/3 • π • h (R2 + r2 + R • r)
Finalmente, se registró el grado de la defoliación apreciado visualmente. Los datos provenientes de un total de 832 plantas y 208 hormigueros se registraron en los cuadrantes de muestreo.
Cubierta del suelo
El estado o condición de la cubierta del suelo se midió utilizando el método de línea de intercepción (Brower et al., 1998), para estimar la proporción de área total ocupada por la cubierta basal de plantas (mantillo), piedras y estiércol, así como suelo desnudo y caminos de hormigas. Con este fin, cinco líneas (5 m de longitud cada una) se colocaron en cada sitio y condición de agostadero (tres sitios, dos condiciones = 30 en total), utilizando una cuerda tensada entre dos estacas; las líneas se dispusieron en una forma alterna y paralela a los transectos de muestreo, a 10 m de distancia de éstos.
Análisis de datos
La densidad absoluta y relativa, y la biomasa volumétrica por hectárea se calcularon para cada especie de planta, así como la densidad de hormigueros y la proporción de cada categoría de cobertura identificada en la superficie del suelo. Los valores estructurales del total de especies registradas se promediaron, para respaldar los respectivos análisis tabulares. Los sitios y condiciones se compararon utilizando sólo los datos de plantas con más de 10 registros numéricos. Se utilizó un diseño completamente al azar con arreglo factorial 2×3: el factor A fue la condición de agostadero (rancho privado y ejido) y el factor B fue el sitio de agostadero (favorable, regular y desfavorable). Las cifras para suelo desnudo, mantillo, vegetación, piedras y estiércol, expresados en porcentajes, se transformaron a arcsen y se analizaron con el procedimiento GLM de SAS (Statistical Analysis System, 1999), con un nivel de significancia de P < 0.05; se utilizó la prueba de Tukey para comparar los valores medios.
Resultados y discusión
En el Cuadro 1 se muestran las especies de plantas registradas y evaluadas en el municipio de Charcas, San Luis Potosí. Con el mismo tamaño de muestra y diseño de muestreo, en el ejido se registraron sólo 22 especies en comparación con las 35 especies registradas en el rancho. La composición de toda la flora del ejido también fue registrada en el rancho, aunque el ejido carece de varias especies forrajeras arbustivas y herbáceas que todavía están presentes en el rancho. Como resultado del historial de pastoreo contraste, la densidad total de especies en el ejido fue sólo el 42.8 % de la registrada en el rancho; por el contrario, la biomasa volumétrica en el rancho fue sólo 27.92 % de la registrada en el ejido. Esto se explica por el predominio de especies más grandes y de larga vida no aptas para el pastoreo en los sitios de agostadero muy deteriorados del ejido. Esto también concuerda con el estudio de Campbell, Jarillo-Rodríguez, López-Ortiz, y Castillo-Gallegos (2013) realizado en Veracruz, México; los autores señalan que el aumento de ganado en la pradera conduce a la abundancia creciente de especies de plantas no forrajeras, mientras que los zacates forrajeros perennes tienden a disminuir. Por otra parte, las especies herbáceas abundantes, aunque de menor tamaño, tales como los zacates deseables si se registraron en el rancho (Cuadros 2 y 3). Finalmente, sólo dos hormigueros activos se registraron en el ejido (en el sitio favorable), en contraste con los 14 encontrados en el rancho. En este sentido, Rojas y Fragoso (2000) señalan que la ausencia de comunidades de hormigas en el centro del Desierto Chihuahuense es un indicador de deterioro. Del mismo modo, Hoffmann (2000) menciona que la riqueza de especies de hormigas se correlaciona negativamente con la intensidad de pastoreo; mientras que estudios en sistemas silvopastoriles intensivos realizados en Colombia por Rivera, Armbrecht y Calle (2013) indican que la riqueza de especies de hormigas puede aumentar con la presencia de vegetación leñosa como Leucaena leucocephala (Lam.) De Wit. Los hormigueros de algunas hormigas cosechadoras como Messor y Pogonomyrmex están documentados como la representación de parches ricos en nutrientes y como el apoyo de mayor biomasa y riqueza de especies de plantas anuales en la periferia de la entrada a los hormigueros (Whitford, Barness, & Steinberger, 2008). Bestelmeyer y Wiens (1996) encontraron forraje de condición de agostadero altamente degradado a alteración menos grave en un suelo degradado extendido del Chaco en el norte de Argentina, debido a prácticas tradicionales de pastoreo, cambios en la cubierta superficial del suelo, desde dispersa hasta estructuralmente compleja, así como mayor riqueza de especies de hormigas en la mejor condición de agostadero.
Cuadro 1 Especies registradas en los agostaderos muestreados del municipio de Charcas, San Luis Potosí, México.
| Especies | Forma de vida | |||
|---|---|---|---|---|
| Herbácea | Arbusto bajo | Arbusto alto | Arborescente | |
| Acacia schaffneri (S. Watson) F.J. Herm. | x | |||
| Agave salmiana Otto ex Salm-Dick | x | |||
| Aristida adscensionis L. | x | |||
| Berberis trifoliolata Moric. | x | |||
| Bouteloua gracilis (Kunth) Lag. ex Griffiths | x | |||
| Bouteloua reederorun Columbus | x | |||
| Bouvardia ternifolia (Cav.) Schldl. | x | |||
| Calliandra eriophylla Benth. | x | |||
| Condalia Cav. | x | |||
| Cylindropuntia imbricata (Haw.) F. Knuth | x | |||
| Dalea bicolor Humb. & Bonlp. ex Willd. | x | |||
| Dasyochloa pulchella (Kunth) Willd. ex Rydb. | x | |||
| Echinocactus platyacanthus Link & Otto | x | |||
| Euphorbia heterophylla L. | x | |||
| Ferocactus pilosus (Salm-Dick) Werderm | x | |||
| Hilaria cenchroides Kunth | x | |||
| Ipomoea orizabensis (Pelletan) Ledeb. ex Steud. | x | |||
| Jatropha dioica Sessé ex Cerv. | x | |||
| Larrea tridentata (Sessé & Moc. ex DC.) Cov. | x | |||
| Leptochloa dubia (Kunth) Nees | x | |||
| Menodora coulteri A. Gray | x | |||
| Mimosa biuncifera Benth. | x | |||
| Muhlenbergia villosa Swallen | x | |||
| Opuntia cantabrigiensis Lynch | x | |||
| Opuntia leucotricha DC. | x | |||
| Opuntia rastrera F. Weber | x | |||
| Opuntia streptacantha Lem. | x | |||
| Parthenium incanum Kunth. | x | |||
| Prosopis laevigata (Willd.) M. Johnston | x | |||
| Rhus microphylla Engelm. | x | |||
| Salvia ballotiflora Benth. | x | |||
| Senna bauhinioides (A. Gray) H.S. Irwin & Barneby | x | |||
| Solanum ehrenbergii (Bitter) Rydb. | x | |||
| Sporobolus airoides (Torrey) Torrey | x | |||
| Yucca L. | x | |||
Cuadro 2 Estructura de la vegetación en sitios del suelo (favorable, adecuado, poco favorable) en el rancho Laguna Seca, considerado con mejor condición de agostadero para el establecimiento de hormigueros de escamoles (Liometopum apiculatum).
| Composición botánica | Biomasa (m3·ha-1) | Total | Densidad (plantas·ha-1) | Total | ||||
|---|---|---|---|---|---|---|---|---|
| Favorable | Adecuado | Poco favorable |
Favorable | Adecuado | Poco favorable |
|||
| Acacia schaffneri | 117.8 | 341.5 | 2,250.4 | 2,709.8 | 647.0 | 67.6 | 3,277.7 | 3,992.3 |
| Agave salmiana | 0.20 | 0.0 | 34.3 | 34.5 | 19.8 | 0.0 | 17.7 | 37.5 |
| Aristida adscensionis | 0.0 | 0.0 | 1.8 | 1.8 | 0.0 | 0.0 | 40,270.0 | 40,270.0 |
| Salvia ballotiflora | 0.0 | 4.7 | 0.0 | 4.7 | 0.0 | 59.5 | 0.0 | 59.5 |
| Berberis trifoliolata | 0.0 | 0.0 | 6.1 | 6.1 | 0.0 | 0.0 | 73.7 | 73.7 |
| Bouteloua gracilis | 3E-02 | 9E-02 | 0.0 | 12E-02 | 7,000.0 | 388.6 | 0.0 | 7,388.6 |
| Bouteloua reederorun | 0.0 | 0.0 | 2.2E-02 | 2.2E-02 | 0.0 | 0.0 | 1,761.1 | 1,761.1 |
| Bouvardia ternifolia | 0.0 | 0.0 | 11E-02 | 11E-02 | 0.0 | 0.0 | 14.6 | 14.6 |
| Echinocactus platyacanthus | 0.0 | 8E-04 | 0.0 | 8E-04 | 0.0 | 10.1 | 0.0 | 10.1 |
| Calliandra eriophylla | 0.0 | 0.0 | 1.2 | 1.2 | 0.0 | 0.0 | 180.6 | 180.6 |
| Cylindropuntia imbricata | 0.0 | 0.0 | 30.4 | 30.4 | 0.0 | 0.0 | 4.9 | 4.9 |
| Condalia sp. | 0.0 | 9E-02 | 0.0 | 9E-02 | 0.0 | 7.2 | 0.0 | 7.2 |
| Dalea bicolor | 2.8 | 8.6 | 18.0 | 29.4 | 115.7 | 66.5 | 41.08 | 223.2 |
| Euphorbia heterophylla | 0.0 | 0.0 | 1.1E-02 | 1.1E-02 | 0.0 | 0.0 | 13.9 | 13.9 |
| Ferocactus pilosus | 0.0 | 2E-03 | 0.0 | 2E-03 | 0.0 | 8.0 | 0.0 | 8.0 |
| Mimosa biuncifera | 0.0 | 1.8 | 0.0 | 1.8 | 0.0 | 44.0 | 0.0 | 44.0 |
| Menodora coulteri | 4E-02 | 1.7 E-02 | 0.0 | 5.8E-02 | 1,011.0 | 853.0 | 0.0 | 1,864.0 |
| Hilaria cenchroides | 2E-03 | 1E-04 | 0.0 | 2.6E-03 | 0.9 | 0.8 | 0.0 | 1.7 |
| Ipomoea orizabensis | 0.0 | 0.0 | 2E-04 | 2E-04 | 0.0 | 0.0 | 2.4 | 2.4 |
| Jatropha dioica | 13E-02 | 0.1 | 0.8 | 1.0 | 31.9 | 120.0 | 657.4 | 809.3 |
| Larrea tridentata | 47.7 | 82.4 | 12.5 | 142.6 | 288.3 | 205.5 | 131.5 | 625.3 |
| Leptochloa dubia | 0.4 | 0.0 | 0.0 | 0.4 | 14.2 | 0.0 | 0.0 | 14.2 |
| Muhlenbergia villosa | 3E-03 | 0.0 | 0.0 | 3E-03 | 0.9 | 0.0 | 0.0 | 0.9 |
| Opuntia leucotricha | 0.0 | 0.0 | 21.7 | 21.7 | 0.0 | 0.0 | 243.5 | 243.5 |
| Opuntia cantabrigiensis | 0.0 | 0.0 | 14E-02 | 14E-02 | 0.0 | 0.0 | 69.1 | 69.1 |
| Opuntia rastrera | 0.0 | 1E-02 | 17.1 | 17.1 | 0.0 | 13.0 | 1,144.8 | 1,157.8 |
| Opuntia streptacantha | 0.0 | 0.0 | 149.1 | 149.1 | 0.0 | 0.0 | 277.9 | 277.9 |
| Parthenium incanum | 1.0 | 0.5 | 0.0 | 1.5 | 17.7 | 1.2 | 0.0 | 18.9 |
| Prosopis laevigata | 112.9 | 271.4 | 98.3 | 482.6 | 725.5 | 697.0 | 546.8 | 1,969.3 |
| Rhus microphylla | 0.0 | 9.7 | 0.0 | 9.7 | 0.0 | 220.0 | 0.0 | 220.0 |
| Senna bauhinioides | 0.0 | 1E-03 | 0.0 | 1E-03 | 0.0 | 20.9 | 0.0 | 20.9 |
| Solanum ehrenbergii | 0.0 | 0.0 | 1E-03 | 1E-03 | 0.0 | 0.0 | 5.1 | 5.1 |
| Yucca spp. | 2,774.9 | 6,093.5 | 897.1 | 9,765.5 | 5,493.6 | 5,248.1 | 1,598.9 | 12,340.6 |
| Dasyochloa pulchella | 0.0 | 0.0 | 2E-03 | 2E-03 | 0.0 | 0.0 | 153.8 | 153.8 |
| Sporobolus airoides | 0.0 | 1.6 | 4.1 | 5.7 | 0.0 | 109.1 | 321.0 | 430.1 |
| Total | 3,057.8 | 6,816.2 | 3,543.3 | 13,417.4 | 15,366.5 | 8,141.8 | 50,807.6 | 74,315.9 |
Cuadro 3 Estructura de la vegetación en sitios del suelo (favorable, adecuado, poco favorable) del ejido Francisco I. Madero, considerado con peores condiciones de agostadero, para el establecimiento de hormigueros de escamoles (Liometopum apiculatum).
| Composición botánica | Biomasa (m3·ha-1) | Total | Densidad (plantas·ha-1) | Total | ||||
|---|---|---|---|---|---|---|---|---|
| Favorable | Adecuado | Poco favorable |
Favorable | Adecuado | Poco favorable |
|||
| Acacia schaffneri | 499.8 | 5,907.0 | 3,200.8 | 9,607.6 | 2,606.4 | 3,418.7 | 4,710.0 | 10,735.1 |
| Agave salmiana | 0.0 | 0.0 | 23.7 | 23.7 | 0.0 | 0.0 | 210.6 | 210.6 |
| Echinocactus platyacanthus | 0.0 | 9E-04 | 0.0 | 9E-04 | 0.0 | 6.5 | 0.0 | 6.5 |
| Bouteloua gracilis | 4E-04 | 9.8E-05 | 2.5E-03 | 3E-03 | 13.1 | 7.9 | 10.4 | 31.4 |
| Bouteloua reederorun | 0.0 | 0.0 | 2E-03 | 2E-03 | 0.0 | 0.0 | 35.4 | 35.4 |
| Cylindropuntia imbricata | 50.0 | 101.0 | 141.3 | 292.3 | 0.1 | 0.5 | 1.1 | 1.70 |
| Dalea bicolor | 0.0 | 0.0 | 12E-02 | 12E-02 | 0.0 | 0.0 | 5.6 | 5.6 |
| Euphorbia heterophylla | 0.0 | 0.0 | 4.4E-03 | 4.4E-03 | 0.0 | 0.0 | 26.0 | 26.0 |
| Hilaria cenchroides | 0.0 | 1E-04 | 0.0 | 1E-04 | 0.0 | 6.7 | 0.0 | 6.7 |
| Jatropha dioica | 26E-02 | 0.0 | 1.3 | 1.6 | 363.9 | 0.0 | 403.0 | 766.9 |
| Larrea tridentata | 158.2 | 207.6 | 289.6 | 655.4 | 1,512.7 | 1,452.3 | 5,366.6 | 8,331.6 |
| Menodora coulteri | 1.3E-03 | 3.8E-03 | 0.0 | 5.1E-03 | 94.7 | 115.2 | 0.0 | 209.9 |
| Opuntia leucotricha | 0.0 | 0.0 | 3.9E-03 | 3.9E-03 | 0.0 | 0.0 | 12.9 | 12.9 |
| Opuntia rastrera | 3.0 | 5.0 | 5.5 | 13.5 | 9.4 | 46.4 | 208.4 | 264.2 |
| Opuntia streptacantha | 0.0 | 0.0 | 279.8 | 279.8 | 0.0 | 0.0 | 771.9 | 771.9 |
| Parthenium incanum | 11E-02 | 7E-02 | 0.0 | 0.18 | 20.1 | 12.7 | 0.0 | 32.8 |
| Parthenium sp. | 0.0 | 0.1 | 0.0 | 0.1 | 0.0 | 19.1 | 0.0 | 19.1 |
| Prosopis laevigata | 0.0 | 31.9 | 9,294.3 | 9,326.2 | 0.0 | 450.0 | 1,213.1 | 1,663.1 |
| Senna bauhinioides | 4E-04 | 1.9E-03 | 2.7E-03 | 5E-03 | 17.0 | 36.9 | 114.9 | 168.8 |
| Sporobolus airoides | 0.0 | 0.1 | 1.0 | 1.1 | 0.0 | 153.0 | 262.5 | 415.5 |
| Dasyochloa pulchella | 1E-03 | 0.0 | 2E-03 | 3E-03 | 428.3 | 0.0 | 1,927.1 | 2,355.4 |
| Yucca sp. | 872.4 | 1,230.6 | 25,744.4 | 27,847.3 | 2,530.6 | 2,257.4 | 949.4 | 5,737.4 |
| Total | 1,583.8 | 7,483.4 | 38,982.0 | 48,049.2 | 7,596.4 | 7,983.3 | 16,228.9 | 31,808.6 |
El pastoreo excesivo y prolongado en el ejido ha dado lugar a cambios profundos en la estructura de la vegetación, reduciendo las diferencias originales entre los sitios de agostadero. Por lo tanto, sólo los datos registrados en el rancho parecen válidos para el contraste estructural entre sitios de agostadero reconocidos por sus diferencias fisonómicas y la abundancia aparente de hormigueros (Cuadro 2). Los resultados del levantamiento completo, sin la eliminación de aquellas especies con menos de 10 registros y con cifras promedio probablemente sesgadas (especialmente con respecto a la densidad), revelan varias diferencias en la composición de plantas, biomasa volumétrica y densidad de plantas de especies comunes en dos o tres sitios de agostadero. Estas diferencias entre los sitios de agostadero se observan también en las cifras estructurales acumuladas, en particular en el caso de la densidad y en la abundancia de hormigueros: 12 en el sitio favorable (6•ha-1), dos en el sitio regular y ninguno en el sitio desfavorable (Figura 6).
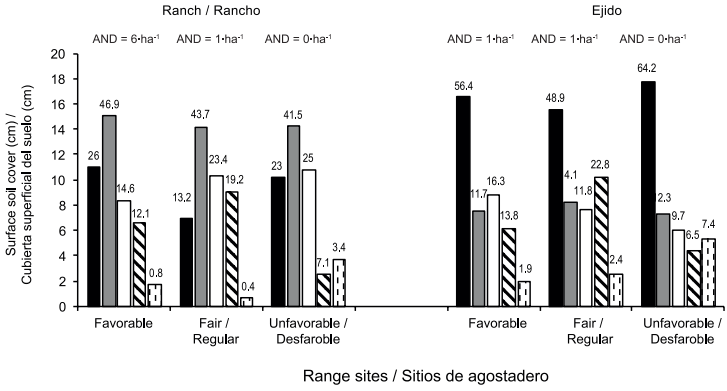
Figura 6 Efecto de la condición de agostadero y sitio tentativo en la cubierta superficial del
suelo del rancho Laguna Seca y ejido Francisco I. Madero en
Charcas, San Luis Potosí (m = líneas Canfield de 5 m cada una).
Suelo desnudo( ), mantillo
(
), mantillo
( ), área
basal de la vegetación (
), área
basal de la vegetación ( ), rocas
(
), rocas
( ) y heces
(
) y heces
( ). AND =
Densidad de hormigueros. Los números en la parte superior de cada
barra indican porcentajes transformados.
). AND =
Densidad de hormigueros. Los números en la parte superior de cada
barra indican porcentajes transformados.
Las especies probablemente relacionadas positivamente con los escamoles son Bouteloua gracilis (Kunth) Lag. ex Griffiths, Menodora coulteri A. Gray, Hilaria cenchroides Kunth, Leptochloa dubia (Kunth) Nees, Muhlenbergia villosa Swallen y Parthenium incanum Kunth (Cuadro 2), todas mostrando una respuesta decreciente al pastoreo y, por lo tanto, consideradas como deseables. En otros sitios del Desierto Chihuahuense, Bestelmeyer y Wiens (2001) también encontraron mayor abundancia de varias especies de hormigas en agostaderos dominados por B. gracilis que en sitios dominados por otras especies de plantas, tales como B. eriopoda (Torr.) Torr., Prosopis glandulosa Torrey y Yucca elata Engelm. La peor condición de agostadero (ejido) evidenció alta abundancia de gobernadora (Larrea tridentata [Sessé & Moc. ex DC.] Cov), especie indeseable, lo cual es consistente con los resultados de Whitford y Steinberger (2011) sobre la invasión y expansión de L. tridentata en agostaderos muy sobrepastoreados del sur de Estados Unidos y norte de México. Para obtener una estimación más sólida de la estructura y dinámica de la vegetación, las especies de plantas con menos de 10 registros se retiraron del análisis; las especies restantes se clasificaron en deseables, menos deseables e indeseables. Con la notable excepción de Aristida adscensionis L., una especie clasificada generalmente como menos deseable y presente sólo en el rancho, la composición de las especies más abundantes (es decir, aquellas con 10 o más registros) fue idéntica en ambas áreas a pesar de su condición de agostadero contrastante (Cuadros 4 y 5 ). Por el contrario, en los valores de densidad de plantas deseables (Cuadro 4) se observaron diferentes patrones como reacción a la distinta carga animal, tipos de ganado y selectividad diversa. Cuatro de las seis especies clasificadas como deseables tuvieron una densidad notablemente decreciente (M. coulteri, B. gracilis, B. reederorun Columbus y Dalea bicolor Humb. & Bonlp. ex Willd.); las otras dos especies fueron relativamente poco afectadas por el deterioro, de manera similar que las especies indeseables Jatropha dioica Sessé ex Cerv. y Cylindropuntia imbricata (Haw.) F. Knuth. Por otra parte, el resto de las especies menos deseables e indeseables tuvieron densidades varias veces mayores en la peor condición de agostadero del ejido; es decir, su abundancia ha aumentado claramente como reacción al exceso de pastoreo. De hecho, Milton y Dean (2010) y Whitford y Steinberger (2011) señalan que el pastoreo excesivo hace que los zacates sean más vulnerables a la invasión de especies indeseables, tales como L. tridentata, C. imbricata y Prosopis spp.
Cuadro 4 Efecto de la condición de agostadero en la densidad de especies de plantas con más de 10 registros en el muestreo realizado en Charcas, San Luis Potosí.
| Valor forrajero | Especies | Mejor condición (rancho) | Peor condición (ejido) | ||
|---|---|---|---|---|---|
| Absoluta (plantas·ha-1) |
Relativa (%) | Absoluta (plantas·ha-1) |
Relativa (%) | ||
| Deseables | Sporobolus airoides | 430.1 | 0.78 | 415.5 | 2.55 |
| Parthenium incanum | 18.3 | 0.03 | 32.8 | 0.20 | |
| Menodora coulteri | 1,863.4 | 3.38 | 209.9 | 1.29 | |
| Bouteloua gracilis | 7,389.0 | 13.44 | 31.4 | 0.19 | |
| Bouteloua reederorun | 1,761.1 | 3.20 | 35.4 | 0.21 | |
| Dalea bicolor | 223.1 | 0.40 | 5.6 | 0.03 | |
| Subtotal | 11,685.0 | 21.25 | 730.8 | 4.49 | |
| Menos deseables | Dasyochloa pulchella | 1,538.5 | 2.79 | 6,214.0 | 38.22 |
| Agave salmiana | 37.2 | 0.06 | 210.6 | 1.29 | |
| Aristida adscensionis | 40,271.3 | 73.26 | - | - | |
| Subtotal | 41,847.1 | 76.12 | 6,424.6 | 39.51 | |
| Indeseables | Larrea tridentata | 625.1 | 1.13 | 8,331.5 | 51.25 |
| Jatropha dioica | 807.0 | 1.46 | 769.2 | 4.73 | |
| Cylindropuntia imbricata | 4.9 | 0.009 | 1.7 | 0.010 | |
| Sub total | 1,437.1 | 2.61 | 9,102.6 | 55.98 | |
| Total | 54,969.5 | 100 | 16,258.1 | 100 | |
Cuadro 5 Efecto de la condición de agostadero en la biomasa volumétrica de especies de plantas con más de 10 registros en el muestreo realizado en Charcas, San Luis Potosí.
| Valor forrajero | Especies | Mejor condición (rancho) | Peor condición (ejido) | ||
|---|---|---|---|---|---|
| Absoluta (m3·ha-1) |
Relativa (%) | Absoluta (m3·ha-1) |
Relativa (%) | ||
| Deseables | Sporobolus airoides | 5.8 | 2.36 | 1.1 | 0.22 |
| Parthenium incanum | 1.5 | 0.61 | 0.1 | 3.6E-02 | |
| Menodora coulteri | 5.8E-02 | 0.02 | 5E-03 | 1E-03 | |
| Bouteloua gracilis | 0.012 | 0.04 | 3E-03 | 6E-04 | |
| Bouteloua reederorun | 2.2E-02 | 8E-03 | 2E-03 | 4E-04 | |
| Dalea bicolor | 29.4 | 11.96 | 0.012 | 2.4E-02 | |
| Subtotal | 36.9 | 15.01 | 1.4 | 0.29 | |
| Menos deseables | Dasyochloa pulchella | 2E-03 | 1E-03 | 3E-03 | 6E-04 |
| Agave salmiana | 34.5 | 14.01 | 23.7 | 4.87 | |
| Aristida adscensionis | 1.8E-02 | 7E-03 | - | - | |
| Subtotal | 34.5 | 14.06 | 23.7 | 4.88 | |
| Indeseables | Larrea tridentata | 142.6 | 58.03 | 318.9 | 65.54 |
| Jatropha dioica | 1.0 | 0.41 | 0.078 | 0.16 | |
| Cylindropuntia imbricata | 30.5 | 12.44 | 141.6 | 29.10 | |
| Sub total | 174.2 | 70.89 | 461.4 | 94.84 | |
| Total | 245.7 | 100 | 486.5 | 100 | |
Por último, vale la pena señalar que en el rancho, con la mejor condición de agostadero, la densidad total de especies deseables y menos deseables fue mayor que en el ejido, con la peor condición de agostadero; lo contrario ocurrió con las de especies indeseables (Cuadro 4). Del mismo modo, Del Curto, Porath, Parsons, y Morrison (2005) mencionan que los agostaderos privados albergan mayor abundancia de plantas deseables y muestran efectos menos graves del exceso de pastoreo en relación con los agostaderos públicos, debido principalmente a mejores prácticas de manejo, tales como una distribución más uniforme del ganado y abrevaderos, pastoreo estacional, duración del pastoreo, y el uso de suplementos y cercas electrificadas. Ramírez y Enríquez (2003) comentan que la riqueza, diversidad y composición de comunidades de hormigas en dos sistemas silvopastoriles tropicales compuestos principalmente por Cynodon plectostachyus (K. Schum.) Pilg., asociados con Prosopis juliflora (Sw.) DC. (2-3 cabezas•ha-1) o con L. leucoephala (4-4.5 cabezas•ha-1), son afectados negativamente por la carga animal.
En cuanto a la biomasa volumétrica (Cuadro 5), el resultado clave es que en el agostadero con la peor condición (ejido) se registró el doble de la cantidad de biomasa que en el agostadero con la mejor condición (rancho). Del mismo modo, en el ejido casi 95 % de la biomasa correspondió a especies indeseables, mientras que la biomasa de las especies deseables fue insignificante (0.3 %) en este agostadero, y relativamente baja (15 %) en el rancho. El deterioro derivado del pastoreo excesivo conduce a la dominancia de plantas más grandes y menos productivas, con predominio de biomasa menos activa (madera) inadecuada para el pastoreo (Odum & Sarmiento, 1998). Por consiguiente, esta estimación estructural instantánea sin consideraciones de sucesión o dinámica puede conducir a evaluaciones erróneas sobre la salud o condición de un sitio de agostadero.
A diferencia de la densidad, el análisis de biomasa reveló que todas las especies de plantas deseables mostraron una respuesta decreciente al pastoreo excesivo, lo que contrasta con el aumento significativo de C. imbricata, también documentado por Milton y Dean (2010) y Zimmermann (2010); por el contrario, la biomasa de J. dioica parece independiente del deterioro. Al respecto, Fulbright, Lozano-Cavazos, Ruthven III, y Litt (2013) señalan que J. dioica está presente en zonas con antecedentes de por más de 30 años.
Cubierta del suelo
La Figura 6 resume los resultados de los efectos del sitio y la condición de agostadero sobre la cobertura de la superficie del suelo. Independientemente del sitio de agostadero, el ejido tiene en promedio el doble del área de suelo desnudo y estiércol, y sólo 60 % de la cobertura basal viva y menos del 30 % del mantillo registrado en el rancho.
Un ANOVA de dos vías detectó diferencia significativa en la condición de agostadero para la variable suelo desnudo (F1,32 = 48.05, P < 0.0001); los efectos de sitio de agostadero (F2,32 = 2.79, P = 0.0763) e interacciones no fueron significativas (F2,32 = 0.47, P = 0.6269). Los porcentajes más bajos de suelo desnudo se observaron en los tres sitios de agostadero del rancho privado: sitio favorable (26 %), regular (13.2 %) y desfavorable (23.0 %).
El mantillo fue afectado por la condición de agostadero (F1,32 = 55.46, P < 0.0001); los efectos de sitio (F2,32 = 0.16, P = 0.8470) e interacciones (F2,32 = 0.20, P = 0.8184) no fueron significativos. Los porcentajes más bajos de mantillo se registraron en los tres sitios del ejido: sitio favorable (11.7 %), regular (14.1 %) y desfavorable (12.3 %)
La cobertura vegetal basal se vio afectada por la condición de agostadero (F1,32 = 5.18, P < 0.0275); los efectos de sitio (F2,32 = 0.08, P = 0.9253) e interacciones no fueron significativos (F2,32 = 2.11, P = 0.1372). Los porcentajes mayores de área basal se observaron en los tres sitios de agostadero del rancho privado: sitio favorable (14.6 %), regular (23.4 %) y desfavorable, (25.0 %).
El ANOVA detectó efecto de sitio de agostadero para la variable piedras (F2,32 = 4.57, P = 0.0179), pero no se encontraron diferencias significativas para la condición de agostadero (F1,32 = 0.18, P = 0.6708) e interacciones (F2,32 = 0.10, P = 0.9077). Los valores más altos se encontraron en el sitio de agostadero clasificado como regular en las condiciones rancho privado y ejido (22.8 y 19.2 %, respectivamente).
La variable heces mostró efecto de sitio de agostadero (F2,32 = 2.56, P < 0.0029), mientras que los efectos de condición (F2,32 = 7.06, P = 0.1196) e interacción no fueron significativos (F2,32 = 0.39, P = 0.6823). Los porcentajes más bajos de heces se presentaron en los sitios favorables y adecuados de las dos condiciones (rancho y ejido).
Dado que en ambos predios, el ganado ha pastado libre y preferentemente sobre ciertas especies de plantas en los tres sitios de agostadero por la falta de cercas interiores, las diferencias existentes en el estado de la superficie del suelo se derivan parcialmente de diferentes grados de utilización histórica del agostadero. El sitio de agostadero considerado favorable para los escamoles en ambas áreas difiere consistentemente en presentar el registro más bajo de estiércol, un indicador de menor perturbación de ganado; de hecho, cuanto mayor es la intensidad y la continuidad del pisoteo, mayor será la probabilidad de destrucción de hormigas y sus caminos.
Cuando se comparó el efecto de condición de agostadero sobre la cobertura del suelo, el ejido, con sólo dos hormigueros registrados, mostró mayor proporción de suelo desnudo (200 %) y estiércol (224 %), y menor proporción de mantillo (25 %) que en el rancho que tuvo 14 hormigueros y mejor condición; sin embargo, las proporciones de área basal de vegetación y piedras fueron similares en ambas condiciones. Estos resultados concuerdan con los de Nash et al. (2001), quienes mencionan que la abundancia de comunidades de hormigas disminuye a medida que la condición de los sitios de agostadero cambia de buena a pobre, y que esto está directamente relacionado con el incremento de la proporción de suelo desnudo y la reducción de la cobertura vegetal.
Conclusiones
En los agostaderos analizados se documentó la condición contrastante que resultó del patrón de uso distinto. Los sitios de agostadero característicos, en términos de diferencias fisonómicas y densidad aparente de hormigueros (favorable, regular y desfavorable), muestran la misma composición general de plantas y la biomasa total instantánea; sin embargo, la densidad y biomasa de las especies deseables y el número de hormigueros encontrados indican que estas zonas se diferencian entre sí a pesar de compartir la misma historia de uso. La relación entre la densidad de hormigueros de Liometopum apiculatum y el tipo de vegetación, condición de agostadero y cubierta del suelo, se observó mediante el registro de la densidad más alta de los hormigueros (14•ha-1) en el sitio favorable del rancho con mejor condición de agostadero. Por lo tanto, siempre que el agostadero tenga los factores ambientales requeridos por la especie L. apiculatum y su condición sea buena, la densidad y calidad (tamaño y actividad de las hormigas) de los hormigueros tenderán a ser óptimas, ya que los recursos y las condiciones ambientales necesarias están disponibles para que las colonias de insectos puedan desarrollarse con normalidad, incluso bajo una presión de pastoreo racional.
Agradecimientos
Este estudio fue financiado por la Fundación Produce San Luis Potosí, A. C. y CONACyT mediante tres becas concedidas a P. Lara Juárez (368440), F. M. Tristán Patiño (370070) y J. A. Rendón Huerta (208836). Los autores también agradecen al Ing. J. Valle Méndez, dueño del rancho Laguna Seca; a la gente de Cerro Blanco en el ejido Francisco I. Madero, quienes amablemente nos permitieron llevar a cabo el estudio en sus tierras; al Sr. Estardo Camacho por su ayuda con el mapa del área de estudio, y finalmente al Sr. J. García por la identificación de las especies vegetales.
REFERENCIAS
Bestelmeyer, B. T., & Wiens, J. A. (1996). The effect of land use on the structure of ground-foraging ant communities in the Argentina Chaco. Ecological Applications, 6, 1225-1240. doi: 10.2307/2269603 [ Links ]
Bestelmeyer, B. T., & Wiens, J. A. (2001). Local and regional-scale responses of ant diversity to a semiarid biome transition. Ecography, 24, 381-392. doi: 10.1111/j.1600-0587.2001.tb00473.x [ Links ]
Bolaños, A., & Aguirre, J. R. (2000). Evaluación preliminar de los agostaderos del occidente del estado de México. Acta Científica Potosina, 15(2), 74‒97. [ Links ]
Brower, J. E., Zar, J. H., & von Ende, C. N. (1998). Field and laboratory methods for general ecology (4th ed.). Boston, Massachusetts, USA: WCB Inc McGraw-Hill. [ Links ]
Campbell, W. B., Jarillo-Rodríguez, J., López-Ortiz, S., & Castillo-Gallegos, E. (2013). Does stocking rate manipulation promote pasture sustainability in the humid tropics? Rangeland Ecology and Management, 66, 348-355. doi: 10.2111/REM-D-11-00110.1 [ Links ]
Del Curto, T., Porath, M., Parsons, C. T., & Morrison, J. A. (2005). Management strategies for sustainable beef cattle grazing on forested rangelands in the Pacific Northwest. Rangeland Ecology and Management, 58, 119-127. doi: 10.2111/1551-5028(2005)58<119:MSFSBC>2.0.CO;2 [ Links ]
Fulbright, T. E., Lozano-Cavazos, E. A., Ruthven III, D. C., & Litt, A. R. (2013). Plant and small vertebrate composition and diversity 36-39 years after root plowing. Rangeland Ecology and Management, 66, 19-25. doi: 10.2111/REM-D-12-00008.1 [ Links ]
García, E. (2004). Modificaciones al sistema de clasificación climática de Köppen. México: Instituto de Geografía, Universidad Nacional Autónoma de México. [ Links ]
Griffin, G. (2002). Indigenous people in rangelands. In A. C. Grice, & K. C. Hodgkinson (Eds.), Global rangelands: Progress and prospects (pp. 29-41). Wallingford, Oxon, UK: CABI. [ Links ]
Heady, H. F., & Child, R. D. (1994). Rangeland ecology and management. Boulder, Colorado, USA: Westview Press. [ Links ]
Hoffmann, B. (2000). Changes in ant species composition and community organization along grazing gradients in semi-arid rangelands of the Northern territory. Rangeland Journal, 22, 171-189. doi: 10.1071/RJ0000171 [ Links ]
Holechek, J. L., Pieper, R. D., & Herbel, C. H. (2011). Range management (6th ed.). River, New Jersey, USA: Prentice Hall. [ Links ]
Instituto Nacional de Estadística y Geografía (INEGI). (2002). Síntesis de información geográfica del estado de San Luis Potosí. Aguascalientes, México: Autor. [ Links ]
Lara-Juárez, P., Aguirre, R. J. R., Castillo-Lara, P., & Reyes- Agüero, J. A. (2015). Biología y aprovechamiento de la hormiga de escamoles, Liometopum apiculatum (Hymenoptera:Formicidae). Acta Zoológica Mexicana, 31, 251-264. Retrieved from http://www.scielo.org.mx/pdf/azm/v31n2/v31n2a12.pdf [ Links ]
Milton, S. J., & Dean, W. R. J. (2010). Plant invasions in arid areas: Special problems and solutions: A South African perspective. Biological Invasions, 12, 3935-3948. doi: 10.1007/s10530-010-9820-x [ Links ]
Nash, M. S., Whitford, W. G., Bradford, D. F., Franson, S. E., Neale, A. C., & Heggem, D. T. (2001). Ant communities and livestock grazing in the Great Basin, USA. Journal of Arid Environments, 49, 695-710. doi: 10.1006/jare.2001.0824 [ Links ]
Odum, E. P. & Sarmiento, F. O. (1998). Ecología. México: McGraw-Hill Interamericana. [ Links ]
Quirk, M. (2002). Managing grazing. In A. C. Grice, & K. C. Hodgkinson (Eds.), Global rangelands: Progress and prospects (pp. 131-145). Wallingford, Oxon, UK: CABI . [ Links ]
Ramírez, M., & Enríquez, M. L. (2003). Importance and diversity of ants in silvopastoral systems in the Cauca Valley, Colombia. Livestock Research for Rural Development, 15(1), article 5. Retrieved August 17, 2016 from http://www.lrrd.org/lrrd15/1/rami151.htm [ Links ]
Ramos-Elorduy, J. (2006). Threatened edible insects in Hidalgo, Mexico and some measures to preserve them. Journal of Ethnobiology and Ethnomedicine, 2, 51. doi: 10.1186/1746-4269-2-51 [ Links ]
Rivera, L. F., Armbrecht, I., & Calle, Z. (2013). Silvopastoral systems and ant diversity conservation in a cattle-dominated landscape of the Colombian Andes. Agriculture, Ecosystems and Environment, 181, 188-194. doi: 10.1016/j.agee.2013.09.011 [ Links ]
Rojas, P., & Fragoso, C. (2000). Composition, diversity, and distribution of a Chihuahuan Desert ant community (Mapimí, México). Journal of Arid Environments, 44, 213-227. doi: 10.1006/jare.1999.0583 [ Links ]
Statistical Analysis System (SAS Institute). (1999). User’s guide: Statistics (CD/ROM Computer file) 8th version. Cary, NC, USA: Author. [ Links ]
Thurow, T. L. (2000). Hydrologic effects on rangeland degradation and restoration processes. In O. Arnalds, & S. Archer (Eds.), Rangeland desertification (pp. 53-66). Dordrecht, Netherlands: Kluwer Academic Publishers. [ Links ]
Vallentine, J. F. (1990). Grazing management. San Diego, California, USA: Academic Press. [ Links ]
Whitford, W. G., Barness, G., & Steinberger, Y. (2008). Effects of three species of Chihuahuan Desert ants on annual plants and soil properties. Journal of Arid Environments, 72, 392-400. doi: 10.1016/j.jaridenv.2007.07.012 [ Links ]
Whitford, W. G., & Steinberger, Y. (2011). Herbivore-plant interactions and desertification in arid land. In J. Seckbach, and Z. Dubinsky (Eds.), All f lesh is grass, cellular origin, life in extreme habitats and astrobiology (pp. 237-256). New York, USA: Springer. doi: 10.1007/978-90-481-9316-5_11 [ Links ]
Zimmermann, H. (2010). Managing prickly pear invasions in South Africa. In A. Nefzaoui, P. Inglese, & T. Belay (Eds.), Improved utilization of cactus pear for food, feed, soil and water conservation and other products in Africa (pp. 157-165). Mekelle, Ethiopia: Proceedings of International Workshop. Retrieved from http://www.cactusnet.org/documents/Issue12.pdf [ Links ]
Recibido: 14 de Abril de 2015; Aprobado: 16 de Mayo de 2016
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
