










Serviços Personalizados
Journal
Artigo
 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versão On-line ISSN 2007-4018versão impressa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.22 no.1 Chapingo Jan./Abr. 2016
https://doi.org/10.5154/r.rchscfa.2015.04.015
Artículos
Germinación y ruptura de latencia en semillas de Daphne giraldii Nitsche (Thymelaeaceae) provenientes del noroeste de China
1 Gansu Agricultural University, College of Agronomy. Lanzhou 730070, China.
2Hexi University, China Key Laboratory of Hexi Corridor Resources Utilization. ZhangYe, 734000, China.
3Gansu Taikang Pharmaceutical Company. Wuwei, 733000, China.
Daphne giraldii Nitsche (Thymelaeaceae) es un arbusto de hoja perenne utilizado ampliamente como planta ornamental y en la industria farmacéutica de China. La planta exhibe latencia que retrasa y reduce la germinación. Este estudio determinó los efectos del tratamiento químico, la estratificación y el tratamiento de siembra de semillas para interrumpir la latencia de D. giraldii. Los resultados mostraron que el pretratamiento químico fue el menos eficaz. El mejor porcentaje de germinación (GP) fue 52.33 % y solamente se observó una tasa de germinación (GR) de 1.19 después de 70 días de la estratificación de semillas a una temperatura de 5 °C. Esto indica que el comportamiento de la germinación de D. giraldii no se vio afectado significativamente por la estratificación. La latencia fisiológica de semillas de D. giraldii se mitigó durante la siembra de semillas. Las semillas enterradas a 100 cm de profundidad por 170 días germinaron exitosamente con 86.5 % e índice de germinación de 10.11, por lo que fue el tratamiento más eficaz para mitigar la latencia de las semillas. Esta información puede ser útil para restaurar y conservar otros arbustos que crecen en el noroeste de China y dilucidar su supervivencia en condiciones extremas similares.
Palabras clave: Estratificación; ácido giberélico; 6-benciladenina; siembra de semilla; capacidad de germinación
Daphne giraldii Nitsche (Thymelaeaceae) is a perennial evergreen shrub that is widely used in the ornamental plant and pharmaceutical industries in China. It exhibits dormancy, which delays and reduces germination. This study determined the effects of chemical treatment, stratification and burial treatment for overcoming the seed dormancy of D. giraldii. Results showed that chemical pre-treatment was less effective in breaking dormancy. The best germination percentage (GP) was 52.33 %, a germination rate (GR) of 1.19 only observed after 70 d of seed stratification at 5 °C treatment, indicating that the germination behavior of D. giraldii was not significantly affected by stratification. The physiological dormancy of D. giraldii seeds was alleviated during burial. Seeds buried at 100 cm depth for 170 days showed the highest germination success with GP of 86.5 % and GR of 10.11, which was the most effective treatment to alleviate seed dormancy. This information may be useful to restore and conserve other shrubs grown in Northwestern China and elucidate their survival under similar extreme environments.
Keywords: Stratification; gibberellic acid; 6-benzyladenine; burial; germination capacity
Introducción
El género Daphne incluye 44 especies de arbustos de hoja perenne distribuidos en el noroeste de China, desde Sichuan y Shanxi hasta las provincias de Gansu. Daphne giraldii Nitsche (Thymelaeaceae) es un arbusto de color café, de crecimiento lento, con una altura máxima de 2 m y tallos flexibles. La planta a menudo crece en tierras vírgenes en laderas, en ocasiones en barrancos, pastizales, lindes del bosque y matorrales en elevaciones de 2,400 m a 2,500 m en las montañas de Qilian en China (Zhao, 2007). Daphne giraldii tiene un gran valor ornamental debido a sus hermosas flores amarillas, foliolos de color verde oscuro y frutos rojos, redondos y con cáscara. La planta resiste el frío, los álcalis, y muestra una gran adaptación a los entornos en el noroeste de China; también se utiliza para el diseño de paisajes. El tallo y la corteza seca de la raíz de D. giraldii son conocidos como "ZuShima" (Zhao, Jin, & Zhang, 2012), los cuales se emplean comúnmente en la medicina tradicional china para el tratamiento de dolores y reumatismo, particularmente para artritis reumatoide (Li, Wu, & Yin, 2002). Debido al creciente consumo de D. giraldii, como planta ornamental y medicinal, es necesaria la producción mediante el desarrollo de métodos para su propagación. La propagación de D. giraldii se hace principalmente mediante esquejes y semillas; sin embargo, los esquejes tienen menor supervivencia y las semillas son sometidas a latencia (Wang, Yan, & Mao, 2012), lo cual no es comercialmente eficiente y por lo tanto, no se recomiendan para la propagación a gran escala. Las semillas de muchas especies de arbustos no pueden germinar, incluso si se siembran en condiciones con buena humedad, oxígeno y suelo (Mark, Tony, & Andrew, 2012). Este fenómeno se conoce como latencia, el cual es muy importante en la naturaleza y es un mecanismo adaptativo que garantiza la supervivencia de algunas especies a través de periodos de estrés ambiental (Baskin & Baskin, 2004; Gusano, Gomez, & Dicenta, 2004; Kermode, 2005); sin embargo, las semillas latentes requieren tratamiento antes de que se siembren. Los tratamientos de presiembra como la estratificación por frío y la siembra de semillas (Merritt, Turne, Clarke, & Dixon, 2007; Travlos, Economou, & Karamanos, 2007) se han utilizado para reducir la dureza de semillas, y mejorar la germinación y tasa de emergencia. Los reguladores químicos de crecimiento tales como 6-benciladenina (6-BA) y ácido giberélico exógeno (GA3) juegan un papel importante en la liberación de la latencia y en la promoción de la germinación al aumentar el potencial de crecimiento del embrión (Kucera, Cohn, & Leubner, 2005; Siddiqui, Mujib, & Maqsood, 2011). El regulador de crecimiento, la temperatura de la estratificación y la siembra de semillas son tres factores que pueden afectar potencialmente la latencia y la germinación. Muchos investigadores estudiaron la influencia de estos factores y encontraron que las semillas de Diren (Melastoma dodecandrum Lour) inmersas en 250 a1,000 mg·L-1 de GA3 o 50 a 200 mg·L-1 de 6-BA durante 24 h puede aumentar significativamente el porcentaje e índice de germinación (Tang, Wei, Yang, Liang, & Wei, 2012). Las semillas de diversas especies necesitan diferentes temperaturas de estratificación (-5 a10 °C) y tiempos (30 a150 días) para la ruptura de la latencia (Walck & Hidayati, 2004; Walck, Hidayati, & Okagami, 2002; Tang et al., 2012). Las semillas de Artemisia ordosica Krasch y Ceratocarpus arenarius L. enterradas a una profundidad de 2 a12 cm en arena o suelo de turba pudieron mejorar la eficacia de los porcentajes de germinación de semillas (Liu, Zhang, Yin, & Zhang, 2013; Wolfgang, 2002). La emergencia de plántulas y el establecimiento exitoso de una población de plantas están regulados principalmente por la ruptura de la latencia. Por ello, la latencia de semillas podría ser un importante factor limitante para la propagación de D. giraldii, ya que no se ha reportado un método ni una técnica exitosa para superar la latencia de sus semillas. Los objetivos de este estudio fueron mejorar la germinación de semillas para la producción de plántulas de D. giraldii en viveros o en semillas sembradas en campo para uso a gran escala y encontrar el método más eficaz para la ruptura de la latencia de semillas con el fin de acelerar la germinación. Específicamente, se estudiaron los efectos de diversos tratamientos de ruptura de latencia (inmersión en GA3, 6-BA, estratificación por frío y siembra de semillas) sobre el rendimiento de la germinación de semillas de D. giraldii.
Materiales y métodos
Recolección de semillas de D. giraldii y condiciones de almacenamiento
Frutas recién maduras, conformadas por una nuececilla (en lo sucesivo llamada semilla) cubierta por una cáscara roja, se recolectaron el 20 de agosto de 2013. Los frutos se obtuvieron a una altitud de 2,450 m, municipio de Buer en las montañas de Qilian con las siguientes coordenadas geográficas: 36° 43' LN y 97° 25' LE, con veranos relativamente cálidos e inviernos fríos en la parte noroeste de China. Las semillas se separaron de la fruta y se secaron después de pasar 6 días en el laboratorio (20 °C a 22 °C). Posteriormente, las semillas se limpiaron y colocaron en bolsas de papel a 25 ± 1 °C y se almacenaron en el laboratorio hasta su uso. Las semillas utilizadas para el tratamiento químico y tratamiento de estratificación se almacenaron durante aproximadamente 8 días, y las semillas utilizadas para los diferentes experimentos de siembra se almacenaron por 35 a 75 días. Las semillas de D. giraldii tienen un diámetro de 5 a 6 mm, son de forma redonda y de color café.
Efecto del tratamiento químico en la latencia de D. Giraldii
Antes del tratamiento experimental, las semillas recolectadas se examinaron para eliminar las que estuvieran manchadas, descoloridas y dañadas. Todas las semillas se lavaron y las semillas flotantes fueron retiradas de la superficie del agua. Las semillas se esterilizaron mediante inmersión en 75 % de alcohol durante 30 min y se enjuagaron tres veces con agua destilada. Posteriormente, la cáscara de las semillas se retiró, rompiendo la cáscara sin dañar el endospermo y el embrión. En consecuencia, el experimento se realizó de la siguiente manera: (1) las semillas fueron inmersas en 100, 150, 200 y 250 mg·L-1 de solución GA3 por 8, 16 y 24 h; (2) las semillas fueron inmersas en 15, 30, 45 y 60 mg·L-1 de solución 6-BA por 8, 16 y 24 h; (3) las semillas fueron inmersas durante 8, 16 y 24 h usando las siguientes combinaciones 100 mg·L-1 GA3 + 15 mg·L-1 6-BA, 150 mg·L-1 GA3 + 30 mg·L-1 6-BA, 200 mg·L-1 GA3 + 45 mg·L-1 6-BA, y 250 mg·L-1 GA3 + 60 mg·L-1 6-BA. Para cada tratamiento se prepararon tres réplicas de 50 semillas. Para el tratamiento testigo se utilizó agua destilada. Después de los tratamientos, las semillas se colocaron en cajas de Petri de 9 cm de diámetro con papel filtro humedecido en 5 mL de agua destilada a temperatura de día/noche de 25/22 °C en un fotoperiodo de 12 h proporcionado por luces fluorescentes (70 a 110 µmol·m-2·s-1, 400-600 nm) y humedad relativa del 50 %. Las cajas de Petri fueron revisadas diariamente hasta iniciar la germinación. Al final del experimento, las semillas se consideraron germinadas si la radícula alcanzaba 1 mm de longitud. La germinación se registró diariamente hasta su cese con un mínimo de 10 días. El porcentaje (GP) e índice de germinación (GR) se calcularon de acuerdo con las siguientes fórmulas (Olmez, Goktur, & Temel, 2007):
GP = n/m *100
donde:
n = Número de semillas germinadas
m = Número de semillas viables e iniciadas
GR = [ (n1* v1) + (n2 * v2) + (ni * vi)] / M
donde:
ni = Número de días para cada conteo
vi = Número de semillas germinada en cada conteo
M = Número total de semillas germinadas.
Efecto de la estratificación en la latencia de D. Giraldii
Las semillas de D. giraldii se colocaron en cajas de Petri de vidrio y se mezclaron con arena húmeda (semillas:arena = 1:6, v/v). La humedad de la arena y las semillas se examinaron continuamente para evitar el secado y la mala aireación. Las placas de Petri de 50 semillas se prepararon por triplicado y se colocaron en una incubadora en la oscuridad a -5 °C, 0 °C, 5 °C y 10 °C durante 30, 45, 60, 70 y 90 días. Las pruebas de germinación se realizaron después de estratificar las semillas por varios periodos a diversas temperaturas.
Efecto de la siembra de semilla en la latencia de D. giraldii
Las semillas fueron preparadas como se describió previamente, el 01 y el 21 de octubre y el 11 de noviembre de 2013, tres paquetes con 50 semillas se colocaron por separado en bolsas de nylon de malla fina. Estos paquetes se enterraron en suelo franco arenoso proveniente de la Universidad de Hexi en Zhangye a profundidades de 20, 40, 60, 80, 100 y 120 cm. No se proporcionó riego suplementario. Los paquetes se recuperaron el 10 de abril de 2014 (las semillas permanecieron enterradas en el suelo durante 190, 170 y 150 días); posteriormente, las semillas se colocaron en cajas de Petri de 9 cm con papel filtro humedecido en agua destilada. El tratamiento testigo fueron semillas limpias colocadas en bolsas de papel y almacenadas en el laboratorio a 25 ± 1 °C. Se llevaron a cabo pruebas de germinación como se describió anteriormente.
Análisis estadístico
Los datos se analizaron con pruebas de ANOVA y rango múltiple de Duncan (Duncan, 1955) y se determinaron los efectos de cada pretratamiento sobre GP y GR con el paquete software SPSS 10.0 (Statistical Package for the Social Sciences [SPSS], 2000). Los datos de GP se transformaron con la función arcoseno antes del análisis. La prueba de comparación múltiple de Tukey (prueba DSH) se llevó a cabo para determinar los tratamientos con diferencias significativas (P = 0.05). Los datos de GP y GR obtenidos en pruebas sobre tratamiento químico se analizaron por medio de un ANOVA de tres factores, mientras que los datos obtenidos en los tratamientos de estratificación y siembra de semillas se analizaron por medio de un ANOVA de dos factores. Las figuras fueron creadas con Origin 8.0 (OriginLab Corporation, 2007) cuando el ANOVA mostró efectos significativos (P ≤ 0.05).
Resultados y discusión
Efectos del tratamiento químico en la latencia de D. giraldii
En las Figuras 1 y 2 se muestra los GP(en un periodo de 10 días) y GR de las semillas tratadas químicamente. Los tratamientos químicos fueron eficaces; los valores de GP y GR de las semillas frescas aumentaron mediante el tratamiento GA3 (Figuras 1 y 2).El porcentaje de germinación de las semillas tratadas con 100, 150, 200 y 250 mg·L-1 de GA3 por 16 h fue de 17.3, 15.8, 15.1 y 14.7 %, respectivamente, mientras que la tasa de germinación más alta de las semillas tratadas con las mismas soluciones y las mismas horas (16) fue de 0.61 (DE = 0.03). Todos los tratamientos fueron significativamente diferentes del testigo tratado con agua destilada (P < 0.05). Existe una tendencia similar de variación de GP y GR mediante el tratamiento GA3 durante 8 y 24 h. Los análisis estadísticos mostraron que el tratamiento 6-BA también pudo aumentar GP y GR significativamente en comparación con el testigo (P < 0.05). Las semillas remojadas en soluciones de 6-BA (30 mg·L-1) durante 16 h germinaron a un máximo de 17.11 % (DE = 1.15, Figura 1), ligeramente inferior a la máxima del tratamiento GA3 (17.3 %, DE = 0.75, Figura 1), pero no se observaron diferencias significativas entre ellos (P > 0.05). Entre todos los tratamientos químicos, los más altos GP y GR fueron 26.33 % (DE = 2.02) y 0.87 (DE = 0.06), respectivamente, que se obtuvieron en las semillas sumergidas en 150 mg·L-1 GA3 + 30 mg·L-1 6-BA durante 8 h en condiciones de laboratorio (Figuras 1 y 2). Los análisis estadísticos también mostraron que GP y GR aumentaron con el incremento en las concentraciones de GA3 + 6-BA y posteriormente disminuyeron significativamente (P < 0.05) con el aumento sostenible de estas soluciones químicas. En el Cuadro 1 se muestra que la interacción entre el tratamiento químico y el tiempo de duración, el tratamiento químico y concentración química afectaron GP y GR (P < 0.05) significativamente, mientras que la interacción entre tiempo de duración y concentración química no tuvo efecto en GP y GR (P > 0.05). A lo largo de los experimentos, la solución mixta de GA3 y 6-BA mejoró de forma significativa la germinación de semillas de D. giraldii en comparación con los otros tratamientos; sin embargo, esta combinación no superó por completo la latencia de las semillas.
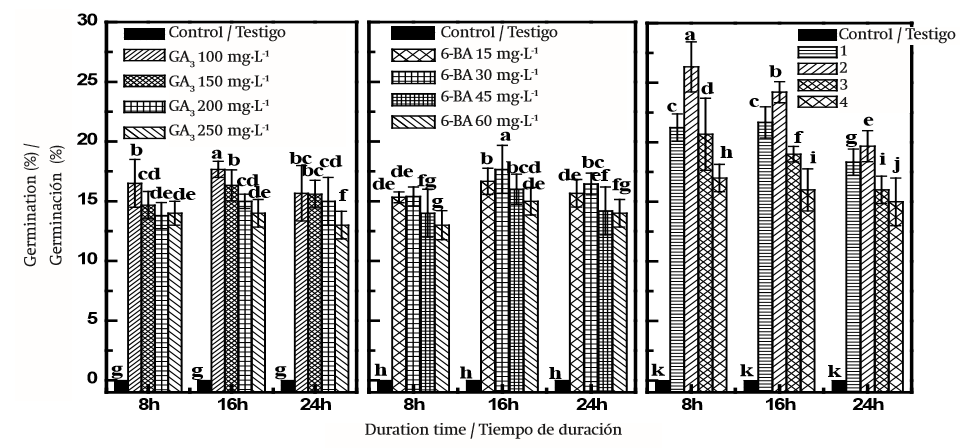
Figura 1. Porcentaje de germinación acumulada de semillas de Daphne giraldii germinadas en diferentes tiempos. 6-BA: 6-benziladenina; GA3: Ácido giberélico. Tratamientos: 1) 100 mg·L-1 GA3 + 15 mg·L-1 6-BA, 2) 150 mg·L-1 GA3 + 30 mg·L-1 6-BA, 3) 200 mg·L-1 GA3 + 45 mg·L-1 6-BA, 4) 250 mg·L-1 GA3 + 60 mg·L-1 6-BA. Los valores (media ± EE, n = 3) marcados con la misma letra (a-k) en el mismo cuadro no son significativamente diferentes mediante la diferencia mínima significativa de Tukey (P ≤ 0.05).
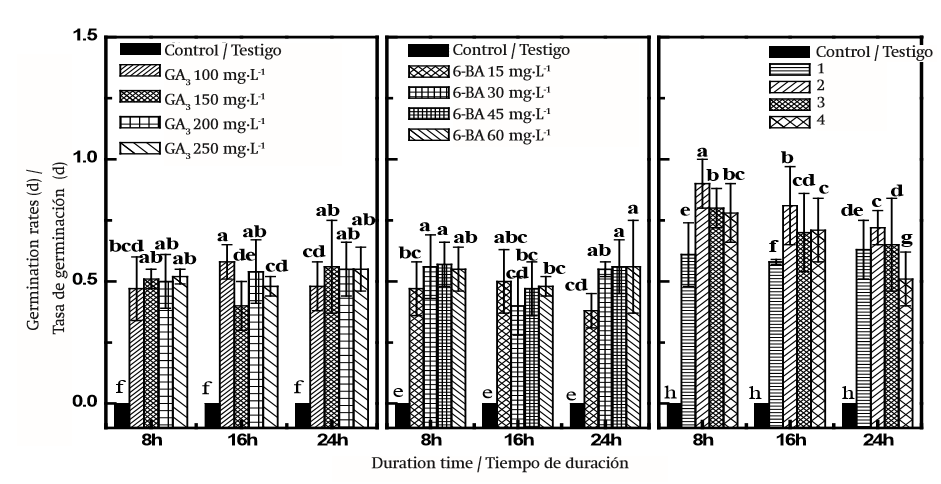
Figura 2. Tasa de germinación acumulada de semillas de Daphne giraldii germinadas en diferentes tiempos. 6-BA: 6-benziladenina; GA3: Ácido giberélico. Tratamientos: 1) 100 mg·L-1 GA3 + 15 mg·L-1 6-BA, 2) 150 mg·L-1 GA3 + 30 mg·L-1 6-BA, 3) 200 mg·L-1 GA3 + 45 mg·-1 6-BA, 4) 250 mg·L-1 GA3 + 60 mg·L-1 6-BA. Los valores (media ± SE, n = 3) marcados con la misma letra (a-h) en el mismo cuadro no son significativamente diferentes mediante la diferencia mínima significativa de Tukey (P ≤ 0.05).
Efecto de la estratificación sobre la latencia de D. giraldii
Como se muestra en la Figura 3, la estratificación, obviamente, aumentó la germinación total de semillas de D. giraldii. Cuando la duración de la estratificación se incrementó de 30 días a 70 días a 5 °C, tanto GP y GR aumentaron significativamente (P < 0.05), y disminuyeron significativamente (P < 0.05) cuando se estratificó por 90 días, lo que muestra la misma tendencia en comparación con el tratamiento a 10 °C. Las semillas estratificadas por 30, 50, 70 y 90 días a 5 °C obtuvieron un GP de 12, 22, 52.33 y 44.67 %, respectivamente. El GP y GR más altos se obtuvieron con este tratamiento; los valores fueron un poco más altos que los valores obtenidos a 10 °C. Estos resultados fueron significativamente diferentes de los resultados del tratamiento a -5 °C y 0 °C y de los obtenidos de semillas no estratificadas (tratamiento testigo). Las semillas con el tratamiento de estratificación por 50 días a 0 °C tuvieron un porcentaje de germinación bajo y significativamente diferente en comparación con las semillas estratificadas por 30 días a 0 °C, ya que las primeras no germinaron (P < 0.05); sin embargo, no hubo diferencia significativa (P > 0.05) con el aumento del tiempo (70 a 90 días). Estos resultados mostraron la misma tendencia que los de GR. Por otra parte, la variación significativa confirmada en este estudio se puede atribuir al tratamiento de semillas estratificadas por 70 días a 5 °C, 0 °C y 10 °C, donde los valores de GP y GR aumentaron, mientras que las semillas a -5 °C no germinaron en ningún periodo de estratificación.
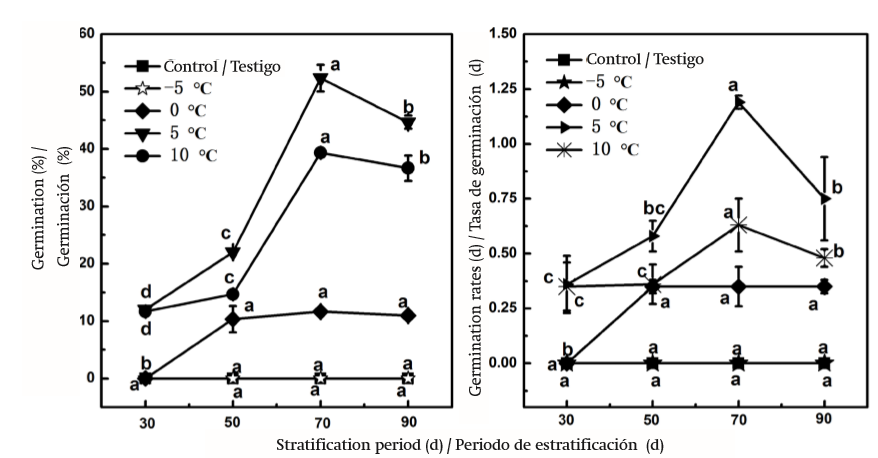
FIGURA 3. Efectos de estratificación en la germinación de semillas de Daphne giraldii. Los valores (media ± EE, n = 3) marcados con la misma letra (a-d) en el mismo cuadro no son significativamente diferentes mediante la diferencia mínima significativa de Tukey (P ≤ 0.05). El testigo fue semillas frescas.
Efecto de la siembra de semilla sobre la latencia de D. giraldii
Los efectos de profundidad de siembra y duración de siembra de semillas sobre los valores de GP y GR se resumen en la Figura 4 y Cuadro 2. Las semillas enterradas a 20 cm por 190 días tuvieron una germinación de 11.3 %, mientras que las semillas enterradas a 100 cm por 170 días tuvieron una germinación de 86.5 %. Las semillas de D. giraldii enterradas a 100 cm de profundidad por 170 días sobrepasaron el 80 % de germinación; la mayoría de las semillas germinaron rápidamente en aproximadamente una semana. En las profundidades restantes, GP disminuyó en el caso de todos los tratamientos y las semillas del tratamiento testigo (obtenidas del laboratorio a 25 ± 1 °C) no germinaron; estos resultados fueron similares que los de GR. Las semillas enterradas a 20 cm de profundidad tuvieron un GP and GR bajo. En el Cuadro 2 se muestra que los efectos del tiempo y profundidad de siembra y la interacción entre estos factores afectaron significativamente (P < 0.05) el GP y GR de semillas de D. giraldii. Un efecto significativo de la interacción entre el tiempo y la profundidad de siembra fue causado aparentemente por un aumento gradual en la profundidad de siembra en el momento adecuado (Figura 4 Cuadro 2).
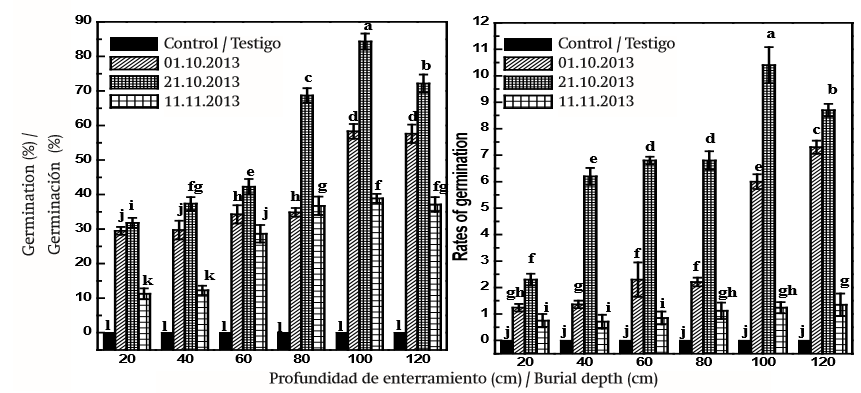
Figura 4. Porcentaje y tasa de germinación de semillas de Daphne giraldii durante diversos periodos. Los valores (media ± EE, n = 3) marcados con la misma letra (a-l) en el mismo marco no son significativamente diferentes mediante la diferencia mínima significativa de Tukey (P ≤ 0.05).
Cuadro 2. ANOVA del porcentaje (GP) e índice (GR) de germinación de semillas de Daphne giraldii como resultado de la variación de profundidad y tiempo de siembra realizado en 2013 en campo abierto (split-plot ANOVA).

Menos del 1 % de las semillas enterradas fueron consideradas muertas dentro de los lotes de semillas exhumadas el 10 de abril de 2014. Las temperaturas medias diarias en el lugar de siembra de octubre de 2013 hasta principios de abril de 2014 fueron de entre -20 °C y 9 °C y aumentaron ligeramente en marzo. Este hallazgo indica que el nivel de latencia aumentó en respuesta a este intervalo de temperatura exterior.
Los problemas relacionados con la latencia de semillas a menudo limitan el uso de algunas especies en la producción de plántulas. Diversas especies de plantas tienen diferentes clases de latencia de semillas, y éstas se pueden dividir en grupos morfológicos, morfofisiológicos, fisiológicos, físicos y combinatorios (Baskin & Baskin, 2004; Gerhard, 2005; Geissler & Gzik, 2010). Xing, Guo, y Wang (2003) reportaron que la planta Stellera chamaejasme L., que pertenece a la familia Thymelaeaceae, tiene la latencia física y fisiológica, que la germinación más alta fue menor de 50 % en virtud de diversos tratamientos de presiembra. Nuestros experimentos muestran que las semillas de D. giraldii pertenecen a la clase fisiológica de acuerdo con el sistema de clasificación de latencia de semillas de Baskin (Baskin & Baskin, 2004). Estos resultados apoyan la conclusión de Wang et al. (2012) de que las semillas frescas de D. giraldii tienen este tipo de latencia.
Algunos investigadores (Airi, Bhatt, Bhatt, Rawal, & Dhar, 2009; Azad, Rahman, & Matin, 2011) han estudiado distintos tratamientos de presiembra para la germinación de semillas para la ruptura de la latencia y, por consiguiente, mejorar el índice de germinación y acelerar el proceso de germinación. La latencia de semillas puede ser interrumpida mediante tratamientos químicos, estratificación gradual y siembra de semillas (Merritt et al., 2007; Ooim, Auld, & Whelan 2006; Travlos et al., 2007). Los resultados de este estudio proporcionan varios puntos de vista no reportados sobre la latencia y germinación de semillas de D. giraldii. El nivel de latencia de un lote de semillas no puede evaluarse directamente, pero se puede medir indirectamente mediante las pruebas de germinación. Los resultados del presente estudio muestran que los GP y GR de semillas de D. giraldii aumentaron significativamente (P < 0.05) en los diferentes pretratamientos en comparación con testigo. Los pretratamientos mostraron que el tratamiento químico no fue muy eficaz para la ruptura de la latencia (Figuras 1 y 2), aunque GA3 y 6-BA pueden aumentar y sincronizar la germinación de semillas de muchas plantas (Chisha, Woodward, & Price, 2007; Nadjafi, Bannayan, Tabrizi, & Rastgoo, 2006). Tang et al. (2012) y Yang, Ye, Wang, y Yin (2009) reportaron que con la inmersión de semillas de M. dodecandrum y Ardisia crenata Sims durante 24 h en 250 mg·L-1 de GA3 o 50 mg·L-1 de 6-BA se podría alcanzar significativamente de 40 a 80 % de germinación . Sin embargo, los datos de este estudio mostraron que la germinación de semillas tratadas con soluciones de mezcla de GA3 y 6-BA en distintos momentos tiene diferencias significativas en comparación con el tratamiento testigo (semillas frescas) y mayor germinación que la obtenida con otros tratamientos químicos individuales. La mejor condición de germinación de semillas tratadas con químicos tuvo un porcentaje de germinación bajo (menos de 30 %) así como el índice de germinación (menos de 1.5). Las semillas frescas de D. giraldii sin tratamiento previo no germinaron, pero los pretratamientos químicos mejoraron ligeramente la germinación, la cual reveló que el embrión mostró algún grado de latencia que fue interrumpido parcialmente por dichos tratamientos. Los tratamientos químicos no fueron completamente eficaces para la ruptura de la latencia, lo que podría atribuirse a las diferencias en los mecanismos que imponen la latencia con otras especies. Baskin y Baskin (2001) indicaron que la latencia de Plumbaginaceae y Juncaginaceae es fisiológica y puede romperse mediante estratificación por frío. La estratificación es utilizada ampliamente para la ruptura de la latencia fisiológica y mejorar la germinación de semillas de numerosas especies (Baker, Steadman, Plummer, & Dixon, 2005; Farshad, Hojat, & Mahmood, 2012). En el presente estudio, las semillas de D. giraldii estratificadas por periodos respondieron favorablemente a los tratamientos químicos. Las semillas estratificadas por 70 días a 5 °C aumentaron el porcentaje de germinación (52.33 %) e índice de germinación (1.19), mientras que la germinación más baja (0 %) se encontró en el tratamiento de estratificación a -5 °C durante 90 días y a 0 °C durante 30 días; Sechenbater y Am (2002) reportaron un resultado similar. GA3 y 6-BA promueven la germinación de semillas, y la estratificación, la ruptura de la latencia a un nivel mayor, de este modo, las semillas de D. giraldii se clasifican por tener latencia fisiológica (Baskin & Baskin, 2004; Nikolaeva, 1977). Estudios fisiológicos recientes han demostrado que la latencia fisiológica incluye un embrión y un recubrimiento, y su suma e interacción determinan el grado de toda la latencia fisiológica de la semilla (Chen, Kuo, & Chien, 2008). En este estudio, la estratificación por frío tiene mejores efectos que el tratamiento químico en la ruptura de la latencia. Un porcentaje de germinación mayor fue alcanzado en pretratamientos de semillas de D. giraldii mediante la estratificación por frío a 5 °C durante 70 días, que fue mejor en comparación con el tratamiento a 10 °C. Esto puede explicar que tal condición puede ser beneficiosa para ablandar el recubrimiento de la semilla en arena húmeda y eliminar los inhibidores de germinación a una temperatura adecuada (5 °C) (Packa, Kwiatkowski, & Graban, 2014).
Los resultados de este estudio mostraron que la latencia fisiológica de semillas de D. giraldii fue mitigada de manera efectiva durante la siembra. La Universidad de Hexi se ubica en el desierto templado y las montañas Qilian; los tipos de suelo en esta zona son franco arcilloso, franco limoso y franco arenoso con 0 a150 cm debajo de la superficie (Li, Qi, Zhao, & Zhang, 1999). Los tratamientos de siembra mitigaron la latencia de las semillas y, por lo tanto, incrementaron el GP y GR de D. giraldii en dicha área. Las semillas enterradas el 21 de octubre de 2013 a 100 cm de profundidad y que se extrajeron el 10 de abril 2014 (las semillas permanecieron en el suelo durante 170 días) tuvieron la germinación más alta con GP de 86.5 % y GR de 10.11. Este tratamiento también fue el más eficaz en el alivio de latencia de las semillas, lo que demuestra que es probable que el periodo comprendido entre 21 de octubre de 2013 y 10 de abril de 2014 sea un periodo predominante, y momento adecuado para la siembra (170 días) y para la liberación de latencia de las semillas de D. giraldii en dicha área. La profundidad de siembra hasta 100 cm mejoró significativamente el porcentaje e índice de germinación; las semillas enterradas a 100 cm de profundidad tuvieron mayor porcentaje e índice de germinación en comparación con aquellas enterradas a 20, 40, 60, 80 y 120 cm. El porcentaje e índice de germinación disminuyeron ligeramente cuando se aumentó de forma continua la profundidad del suelo (120 cm). Esto tal vez sea resultado del bajo nivel de oxígeno a mayores profundidades del suelo; niveles muy bajos de oxígeno a cierta profundidad pueden llevar a la inducción de la latencia secundaria (Malik & Vanden, 1988; Poinar & Columbus, 1992). Las semillas enterradas de D. giraldii podrían estar en un ambiente hipóxico y la germinación de las semillas se vería afectada por la variación en la disponibilidad de oxígeno del suelo relacionado con diferentes profundidades de enterramiento (Yan, Liu, Li, & Ma, 2007). Sin embargo, se requiere más estudio para explorar los niveles de oxígeno en diversas profundidades de enterramiento de esta zona. Los resultados de este estudio son diferentes de los resultados reportados por otros autores en los últimos años. En un estudio del noreste de China, la germinación de semillas de arbustos de A. ordosica mejoró mediante la siembra a 5 cm de profundidad durante seis meses (Liu et al., 2013), mientras que otro estudio en Turquía reveló que las mayores profundidades (> 20 cm) podrían inducir semillas a una latencia secundaria (Mennan, 2003). Las discrepancias anteriores podrían ser el resultado de las variaciones genéticas en las poblaciones estudiadas, las condiciones ambientales, la composición del suelo y los tratamientos aplicados antes de la germinación. Durante el invierno, la superficie de la tierra en el área experimental está congelada porque la temperatura baja a -20 ° C en el exterior (Li, Gao, Wang, & Wang, 2013). A principios de la primavera, la temperatura aumenta de 5 °C a 9 °C y el suelo se moja debido a que el hielo y la nieve se derriten. Así, es probable que la temporada de invierno sea un periodo predominante para la liberación de latencia de las semillas de D. giraldii y la siembra por 170 días afecta y mejora la germinación. La naturaleza fisiológica de la latencia del embrión en D. giraldii aún no está aclarada, pero una posible explicación es dada por Walker (1971), quien indica que la latencia es controlada en cierta medida por algunos inhibidores en la semilla. Estos inhibidores se forman posiblemente en una etapa temprana en el desarrollo de la semilla para evitar el proceso de germinación. En este estudio, la respuesta puede deberse a la mejor degradación de los inhibidores de la germinación en el embrión durante 170 días de siembra en esta área bajo una profundidad adecuada del suelo. Por lo tanto, se recomienda utilizar un tratamiento de enterramiento apropiado para la ruptura de la latencia en futuros estudios de germinación de D. giraldii. Esta información puede ser útil para cultivar y conservar otros arbustos que crecen en las montañas de Qilian en China y dilucidar su supervivencia en condiciones extremas similares.
Conclusiones
A partir de esta investigación se puede concluir que la mala regeneración natural de D. giraldii se atribuye principalmente a la latencia fisiológica. Este estudio indicó que la estratificación de semillas a 5 °C durante 170 días parece ser un método más eficaz para la ruptura de la latencia de semillas de D. giraldii en comparación con el tratamiento químico. Por lo anterior, las semillas fueron enterradas en suelo franco arenoso a 100 cm de profundidad por 170 días para ofrecer el método de ruptura de latencia más conveniente y eficaz para D. giraldii. Estos hallazgos aportan una contribución significativa a los esfuerzos de conservación de esta especie endémica cultivada en el noroeste de China.
Agradecimientos
Los autores agradecen por el apoyo financiero proporcionado por la organización National Natural Science Foundation de Gansu (Núm. 1506RJZG052, Núm. 1308RJZG156) y el apoyo financiero por parte de la Universidad de Hexi (XZ2013-03). También agradecemos a Pr. Enhe Zhang y Qinlin Wang por la asistencia técnica proporcionada.
REFERENCIAS
Airi, S., Bhatt, I. D., Bhatt, A., Rawal, R. S., & Dhar, U. (2009). Variations in seed germination of Hippophae salicifolia with different presoaking treatments. Journal of Forest Research, 20, 27-30. doi: 10.1007/s11676-009-0005-3 [ Links ]
Azad, M. S., Rahman, M. T., & Matin, M. A. (2011). Seed germination techniques of Phoenix dactylifera: A new experience from Bangladesh. Frontiers of Agriculture in China, 5, 241-246. doi: 10.1007/s11703-011-1086-2 [ Links ]
Baker, K. S., Steadman, K. J., Plummer, J. A., & Dixon, K. W. (2005). Seed dormancy and germination responses of nine Australian fire ephemerals. Plant and Soil, 277, 345-348. doi: 10.1007/S11104-005-7971-9 [ Links ]
Baskin, C. C., & Baskin, J. M. (2004). A classification system for seed dormancy. Seed Science Research, 14, 1-16. doi: 10.1079/SSR2003150 [ Links ]
Baskin, C. C., & Baskin, J. M. (2001). Seeds: Ecology, biogeography, and evolution of dormancy and germination. Plant Ecology, 152, 204-205. doi: 10.1023/A:1011465920842 [ Links ]
Chen, S. Y., Kuo, S. R., & Chien, C. T. (2008). Roles of gibberellins and abscisic acid in dormancy and germination of red bayberry (Myrica rubra) seeds. Tree Physiology, 28, 1431-1439. Obtenido de http://treephys.oxfordjournals.org/content/28/9/1431.full.pdf [ Links ]
Chisha, K. E., Woodward, S., & Price, A. (2007). Comparison of the effect of mechanical scarification and gibberellic acid treatments on seed germination in Pterocarpus angolensis. Southern Hemisphere Forestry Journal, 69, 63-70. doi: 10.2989/SHFJ.2007.69.1.9.171 [ Links ]
Duncan, D. B. (1955). Multiple range and multiple F tests. Biometrics, 11,1-42. Obtenido de http://www.jstor.org/stable/3001478 [ Links ]
Farshad, D., Hojat, G. M., & Mahmood , E. A. (2012). Overcoming seed dormancy of mooseer (Allium hirtifolium) through cold stratification, gibberellic acid, and acid scarificati. Journal of Forestry Research, 23(4), 707-710. doi: 10.1007/s11676-012-0314-9 [ Links ]
Geissler, K., & Gzik, A. (2010). Germination ecology of three endangered river corridor plants in relation to their preferred occurrence. Flora, 205(9), 590-598. doi: 10.1016/j.flora.2010.04.008 [ Links ]
Gerhard, L.G. (2005). Glucanase gene expression in low hydrated seeds as a mechanisms for dormancy release during tobacco after-ripening. The Plant Journal, 41, 133-145. doi: 10.1111/j.1365-313X.2004.02284.x [ Links ]
Gusano, M. G., Gomez, P. M., & Dicenta, F. (2004). Breaking seed dormancy in almond (Prunus dulcis Mill.) D. A. Webb. Scientia Horticulturae, 99, 363-370. doi: 10.1016/j.scienta.2003.07.001 [ Links ]
Kermode, A. R. (2005). Role of abscisic acid in seed dormancy. Journal of Plant Growth Regulation, 24, 319-344. doi: 10.1007/s00344-005-0110-2 [ Links ]
Kucera, B., Cohn, M. A., & Leubner, M. G. (2005). Plant hormone interactions during seed dormancy release and germination. Seed Science Research, 15, 281-307. doi: 10.1079/SSR2005218 [ Links ]
Li, S. H., Wu, L. J., & Yin, H. Y. (2002). Chemical and pharmacological advances of the study on Zushim. Chinese Journal of Traditional Chinese Medicine, 27(6), 401-402. [ Links ]
Li, F. X., Qi, S. Z., Zhao, F. H., & Zhang, Y. D.(1999). Soil basic classification of Linze like area in Hexi Corridor. Chinese Journal of Soil Science, 30, 13-17. Obtenido de http://www.cnki.com.cn/Article/CJFDTotal- TRTB1999S1003.htm [ Links ]
Liu, H. L., Zhang, L. W., Yin, L. K., & Zhang, D. Y. (2013). Effects of temperature, dry storage, and burial on dormancy and germination of seeds of 13 desert plant species from sand dunes in the Gurbantunggut Desert, Northwest China. Arid Land Research and Management, 27, 65-78. doi: 10.1080/15324982.2012.719569 [ Links ]
Li, H. Y., Gao, Z. R., Wang, S., & Wang, H. Y. (2013). Extreme temperature variation of Hexi Corridor in recent 60 years. Arid Land Geography, 38, 1-5. doi: 10.13826/j.cnki.cn65-1103/x.2013.01.001 [ Links ]
Malik, N., & Vanden, B. W. H. (1988). The biology of Canadian weeds Galium aparine L. and Galium spurium L. Canadian Journal of Plant Science, 68, 481-499. doi: 10.4141/cjps88-059 [ Links ]
Mark, K. J., Tony, D. A., & Andrew, J. D. (2012). Projected soil temperature increase and seed dormancy response along an altitudinal gradient: Implications for seed bank persistence under climate change. Plant and Soil, 353, 289-303. doi: 10.1007/s11104-011-1032-3 [ Links ]
Merritt, D. J., Turne, S. R., Clarke, S., & Dixon, K. W. (2007). Seed dormancy and germination stimulation syndromes for Australian temperate species. Australian Journal of Botany, 55, 336-344. doi: 10.1071/BT06106 [ Links ]
Mennan, H. (2003). The effects of depth and duration of burial on seasonal germination, dormancyand viability of Galium aparine and Bifora radians seeds. Journal of Agronomy and Crop Science, 189(5), 304-309. doi: 10.1046/j.1439-037X.2003.00048.x [ Links ]
Nadjafi, F., Bannayan, M., Tabrizi, L., & Rastgoo, M. (2006). Seed germination and dormancy breaking techniques for Ferula gummosa and Teucrium polium. Journal of Arid Environments, 64, 542-547. doi: 10.1016/j.jaridenv.2005.06.009 [ Links ]
Nikolaeva, M. G. (1977). Factors controlling the seed dormancy pattern. In Khan, A. A. (Ed.), The physiology and biochemistry of seed dormancy and germination (pp. 51-74). USA: Elsevier/North-Holland. [ Links ]
Olmez, Z., Gokturk, A., & Temel, F. (2007). Effect of cold stratification, sulfuric acid, submersion in hot and top water pretreatment on germination of blad-der-senna (Colutea armena Boiss. & Huet.) seeds. Seed Science and Technology, 35, 266-271. doi: 10.15258/sst.2007.35.2.02 [ Links ]
Ooim, K. J., Auld, T. D., & Whelan, R. J. (2006). Dormancy and the fire-centric focus: Germination of three Leucopogon species (Ericaceae) from south-eastern Australia. Annals of Botany, 98, 421-430. doi: 10.1093/aob/mcl118 [ Links ]
OriginLab Corporation. (2007). Origin 8.0 software. Northampton, MA, USA: Author. [ Links ]
Packa, D., Kwiatkowski, L., & Graban, W. (2014). Germination and dormancy of sida hermaphrodita seeds. Seed science and technology, 42,1-15. doi: 10.15258/sst.2014.42.1.01 [ Links ]
Poinar, G. O., & Columbus, J. T. (1992). The induction of secondary seed dormancy by oxygen deficiency in a barnyard grass Echinochloa crus-galli. Experientia, 48, 904-906. doi: 10.1007/BF02118432 [ Links ]
Sechenbater, M. L., & Am, L. (2002). Effect of plant hormones on seed germination of Prunusm ongolica Maxim. Journal of Inner Mongolia Normal University (Nature Science), 31, 384-387. doi: 10.3969/j.issn.1001-8735.2002.04.017 [ Links ]
Siddiqui, Z., Mujib, A., & Maqsood, M. (2011). Liquid overlaying improves somatic embryogenesis in Catharanthus roseus. Plant Cell Tissue Organ Cult, 104, 247-256. doi: 10.1007/s11240-010-9828-z [ Links ]
Statistical Package for the Social Sciences (SPSS). (2000). SPSS 10.0 software. Chicago, IL, USA: Author. [ Links ]
Tang, H., Wei, J. Q., Yang, Q. H., Liang, H. L., & Wei, X. (2012). Germination and dormancy-breaking of Diren (Melastoma dodecandrum) seeds. Seed Science and Technology, 40, 1-10. doi: 10.15258/sst.2012.40.1.01 [ Links ]
Travlos, I. S., Economou, G., & Karamanos, A. I. (2007). Germination and emergence of the hard seed coated Tylosema esculentum (Burch) A. Schreib in re- sponse to different pre-sowing seed treatments. Journal of Arid Environments, 68, 501-507. doi: 10.1016/j.jaridenv.2006.07.001 [ Links ]
Wang, Q. L., Yan, F., & Mao, Z. H. (2012). Studies on dormancy of Daphne giraldii Nitsche (Thymelaeaceae) seeds. Chinese Horticulture Abstracts, 12, 9-13. Obtenido de http://www.cnki.com.cn/Article/CJFDTOTAL-YUWZ201212008.htm [ Links ]
Walker, M. G. (1971). Changes in germination promotion and inhibition in seed extracts of sub-terranean clover (Trifolium subterraneum L.) related to dormancy and germination. Australian Journal of Biology Science, 24, 897-903. doi: 10.1071/BI9710897 [ Links ]
Walck, J. L., & Hidayati, S. N. (2004). Germination ecophysiology of the western North American species Osmorhiza depauperata (Apiaceae): Implications of preadaptation and phylogenetic niche conservatism in seed dormancy evolution. Seed Science Research, 14, 387-394. doi: 10.1079/SSR2004184 [ Links ]
Walck, J. L., Hidayati, S. N., & Okagami, N. (2002). Seed germination ecophysiology of the Asian species Osmorhiza aristata (Apiaceae): Comparison with its North American congeners and implications for evolution of types of dormancy. American Journal of Botany, 89, 829-835. doi: 10.3732/ajb.89.5.829 [ Links ]
Wolfgang, S., & Gerhard, R. (2003). Variation in seed dormancy of the wetland sedge, Carex elongata, between populations and individuals in two consecutive years. Seed Science Research, 13, 315-322. doi: 10.1079/SSR2003148 [ Links ]
Xing, F., Guo, J. X., & Wang, Y. H. (2003). Seed germination characteristics and regeneration mechanism of Stellera chamaejasme population. Chinese Journal of Applied Ecology, 14, 1851-1854. doi: 10.13287/j.1001-9332.2003.0409 [ Links ]
Yan, Q. L., Liu, Z. M., Li, X. H., & Ma, J. L. (2007). Effects of burial on seed germination characteristics of 65 plant species on Horqin semi-arid steppe. Chinese Journal of Applied Ecology, 18, 777-782. doi: 10.13287/j.1001-9332.2007.0131 [ Links ]
Yang, Q. H., Ye, W. H., Wang, Z. M., & Yin, X. J. (2009). Seed germination physiology of Ardisia crenata var. bicolor. Seed Science and Technology, 37, 291-302. doi: 10.15258/sst.2009.37.2.04 [ Links ]
Zhao, R. N. (2007). Resources of Chinese traditional and herbal drugs in Gansu. Lanzhou, China: Gansu Science and Technology Press. [ Links ]
Zhao, J., Jin, X. J., & Zhang, H. J. (2012). Research progress of Daphne giraldii. Chinese Wild Plant Resources, 31, 12-14. doi: 10.3969/j.issn.1006-9690.2012.06.003 [ Links ]
Recibido: 14 de Abril de 2015; Aprobado: 25 de Noviembre de 2015
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
