










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.17 no.spe Chapingo ene. 2011
https://doi.org/10.5154/r.rchscfa.2010.08.062
El estímulo del macho cabrío incrementa la función reproductiva de las cabras criollas del semidesierto mexicano independientemente del régimen de fotoperiodo
The male effect stimulus increases reproductive activity of mexican criollo goats irrespective of the photoperiodic regime
Romana Melba Rincón-Delgado1; Carlos Fernando Aréchiga1¶; Francisco Javier Escobar1; José Manuel Silva1; Jairo Iván Aguilera-Soto1; Marco Antonio López-Carlos1; Heriberto Rodríguez1; César Alberto Meza-Herrera2; Javier Valencia3.
1 Universidad Autónoma de Zacatecas. Unidad Académica de Medicina Veterinaria y Zootecnia. Zacatecas, Zac. C. P. 98000. MÉXICO. Tel. (011-52-478) 985-0202). Correo-e: arechiga@uaz.edu.mx o arechiga.carlos@gmail.com (¶Autor para correspondencia).
2 Universidad Autónoma Chapingo-Unidad Regional Universitaria de Zonas Áridas, Bermejillo, Durango. MÉXICO.
3 Universidad Nacional Autónoma de México. Cd. Universitaria C. P. 04410. México, D. F.
Recibido: 23 de agosto, 2009
Aceptado: 23 de septiembre, 2009
RESUMEN
Se evaluó la influencia de un macho cabrío sexualmente activo sobre el inicio de la actividad reproductiva (actividad hipotalámica y ovárica) y la sucesión de los ciclos estrales en cabras criollas expuestas a fotoperiodo artificiales alternados (decreciente/creciente) en un rango de 13.4 a 10.6 horas de luz por día hasta completar 6 ciclos foto-periódicos de 90 d por cada ciclo: fotoperiodo creciente (n=3); fotoperiodo decreciente (n=3). Las cabras criollas procedentes del semidesierto zacatecano (n=30) fueron asignadas de manera aleatoria a dos grupos: 1) cabras expuestas a un macho cabrío sexualmente activo (n=15); 2) cabras no expuestas al macho cabrío (ausencia de macho cabrío; n=15). Dentro de cada grupo experimental existían cabras ovariectomizadas (OVX, n=5), cabras ovariectomizadas e implantadas con estradiol (OVX + E2, n=5), y cabras ovariointactas (testigo, n=5). Se recolectaron muestras de suero sanguíneo de las cabras OVX y OVX + E2, cada cuatro semanas, durante 6 h en intervalos de 15 min (es decir, 24 muestras/día), para determinar la frecuencia (FREQ), la amplitud (AMP) y la concentración (CONC) de la hormona luteinizante (LH). En las cabras testigo (no tratadas), se obtuvieron 2 muestras de sangre a la semana para cuantificar los niveles de progesterona en suero sanguíneo mediante un radioinmunoanálisis (RIA). Las cabras con implantes de E2 (cabras OVX + E2) mostraron un incremento en la frecuencia de los pulsos de la hormona luteinizante en comparación con las cabras OVX sin implante de E2 (2.0±0.5 vs.0.7 ± 0.1 LH pulsos/6 h). La presencia de un macho sexualmente activo aumenta la frecuencia, la amplitud y la concentración de la hormona luteinizante en cabras OVX en comparación con cabras no expuestas a machos (Frecuencia: 3.2 ± 0.4 vs. 0.7 ± 0.1 pulsos/6 h; Amplitud: 1.6 ± 0.1 vs. 0.8 ± 0.3 ng·mL-1; Concentración 5.3 ± 0.6 vs. 2.0 ± 0.9 ng·mL-1) (P<0.001). En las cabras testigo (con los ovarios intactos), la presencia y estímulo del "efecto macho" indujo una mayor función reproductiva (actividad lútea) y un incremento considerable en la sucesión de los ciclos estrales, mostrando un mayor número de días en actividad lútea (44 ± 9.05 vs. 32.3 ± 20.6); menor número de días en anestro (4.0 ± 4.8 vs. 51.9 ± 28) y un mayor número de fases lúteas (15.7 ± 4.4 vs. 11.3 ± 8.7). Todo ello sin afectar los niveles séricos de la hormona progesterona (6.8 ± 0.8 vs. 7.3 ± 0.5) (P>0.05). En conclusión, la presencia de un macho cabrío sexualmente activo estimuló una mayor actividad ovárica de las cabras, disminuyendo la duración del anestro estacional, incluso bajo condiciones de fotoperiodo artificial-controlado ascendente. Esta estrategia pudiera permitir incrementar la función reproductiva y de esa manera reducir la temporada de anestro o inactividad reproductiva de las cabras criollas del norte de México.
Palabras clave: Cabras, fotoperiodo, progesterona, hormona luteinizante, presencia de macho.
ABSTRACT
Influence of a sexually-active male buck was evaluated on the onset of reproductive function (hypothalamic and ovarian activity) and estrous cycles progression in Criollo goats from the northen Mexican desert, exposed to an alternated (decreasing/increasing) artificially controlled photoperiod fluctuating within 13.4 to 10.6 light hours per day until fulfilling 6 photoperiodic cycles of 90 d each cycle: ascending (n=3); and descending (n=3). Mexican-native Criollo goats (n=30) were randomly assigned into 2 treatment groups: 1) goats exposed to a sexually-active male buck (n=15); 2) goats not exposed to a male buck (absence of a male buck; n=15). Each experimental group of goats included ovariectomized goats (OVX, n=5), ovariectomized and estradiol-implanted goats (OVX + E2, n=5), and intact-ovaries goats (Control, n=5). Blood samples were taken from OVX and OVX + E2 goats, every four weeks, during 6 h at 15 min intervals (i.e., 24 samples/day), to determine frequency (FREQ), amplitude (AMP), and concentration (CONC) of luteinizing hormone (LH). For Control goats, blood samples were taken twice every week in order to quantify serum-progesterone levels through radioimmunoanalysis (RIA). Goats implanted with E2 (OVX + E2-goats) showed an increased LH pulse frequency compared to OVX-goats without an E2 implant (2.0 ± 0.5 vs. 0.7 ± 0.1 LH pulses/ 6 h). Presence of a sexually-active male buck increased frequency, amplitude and concentration of LH in OVX goats compared to goats not exposed to males (Frequency: 3.2 ± 0.4 vs. 0.7 ± 0.1 pulses/6 h; Amplitude: 1.6 ± 0.1 vs. 0.8 ± 0.3 ng·mL-1; Concentration: 5.3 ± 0.6 vs. 2.0 ± 0.9 ng·mL-1) (P<0.001). In Control goats (intact ovaries), male exposure increased luteal activity and estrous cycle progression refected as a greater number of days in luteal activity (44 ± 9.05 vs. 32.3 ± 20.6); a reduced number of days in anestrous (4.0 ± 4.8 vs. 51.9 ± 28) and a greater number of luteal phases (15.7 ± 4.4 vs. 11.3 ± 8.7) without compromising serum progesterone levels (6.8 ± 0.8 vs. 7.3 ± 0.5) (P>0.05). In conclusion, presence of a sexually-active male buck induced a greater ovarian activity in Criollo goats, shortening seasonal anestrous even during an ascendent controlled-photoperiod. Such strategy, using a sexually-active male buck might be helpful to increase ovarian activity and reproductive function during the seasonal anestrous of Criollo goats from dry and arid areas of northern Mexico and similar regions of the world.
Key words: Goats, photoperiod, progesterone, ovarian activity, male effect.
INTRODUCCIÓN
Un mundo global demanda un incremento en la producción de cabras y en la implementación de tecnologías reproductivas (Aréchiga et al., 2008; Aréchiga y Rincón, 1998). Las cabras criollas muestran una estacionalidad reproductiva, aun en altiplanos áridos de México, como se puede ver mediante la evaluación del aparato reproductor (Valencia et al., 1986, 1990), midiendo los niveles de progesterona sérica en cabras no preñadas (Escobar et al., 1997) o mediante la evaluación de los patrones de actividad reproductiva (Chemineau et al., 2004). Durante el anestro estacional, los machos cabríos mostraron una actividad reproductiva y un estímulo sexual reducido. Los tratamientos fotoperiódicos han demostrado ser exitosos ya que aumentan la actividad sexual de las cabras que se localizan en latitudes subtropicales (Delgadillo et al., 2002, 2003, 2004) así como en machos cabríos criollos expuestos a ciclos fotoperiódicos alternando luz-oscuridad (16 h L:8 h D); con o sin tratamiento con melatonina (Delgadillo et al., 1995, 2001, 2002, 2004, 2006). Se propuso una combinación de fotoperiodo y efecto macho para regular la estacionalidad reproductiva de las cabras (Delgadillo et al., 2003, 2004,2006, 2009). De hecho, el "efecto macho" ha sido reconocido como una técnica valiosa para inducir la ovulación fértil sincronizada durante el anestro post-parto y estacional tanto en cabras como en ovejas (Gelez y Fabre- Nys, 2004; Restall, 1992; Scaramuzzi y Martin, 2008; Walkden-Brown et al., 1999). Los machos sexualmente activos expuestos a ovejas (Ungerfeld et al., 2004) y cabras prepúberes (Amoah y Bryant, 1984; Mellado et al., 2000) y adultas sincronizaron la actividad sexual ya sea por anestro estacional o por lactancia (Pellicer-Rubio et al., 2007; Veliz et al., 2002, 2006a, 2006b). Un gran número de cabras expuestas a un macho sexualmente activo mostraron una conducta estral dos o tres días después de ser reunidos con el macho cabrío (Chemineau, 1983, 1987) y ovularon al reaccionar a un macho mediante el aumento de la secreción de GnRh y de la hormona luteinizante (Chemineau et al., 1986a, 1986b, 2006).
Además de eso, existen vínculos claros entre la disponibilidad del combustible metabólico (glucosa, piruvato y lactato) y la función reproductiva (Ebling, 2005). De hecho, los cambios en los niveles de sangre o en las hormonas metabólicas son señales importantes que informan sobre el estado nutricional de los mamíferos (Gámez-Vázquez et al., 2008; Meza-Herrera et al., 2007, 2008). Una explicación es que la reacción a los suplementos alimenticios altera la glucosa, la insulina, la leptina o IGF-I y posiblemente otras hormonas metabólicas que a su vez puede afectar la función reproductiva (Guerra-García et al., 2009; Meza-Herrera et al., 2004, 2008, 2010a, 2010b; Scaramuzzi et al., 2006). Así mismo, los suplementos bajo condiciones de pastoreo han mejorado la tasa de ovulación y de preñez de las cabras criollas expuestas al efecto macho (De Santiago-Miramontes et al., 2008; Fitz-Rodriguez et al., 2009). No obstante, se ha resaltado que la "innovación" del macho podría ser más importante que el aislamiento del macho (Delgadillo et al., 2009). La presencia del macho también ha inducido a la actividad reproductiva en implantación de melatonina en cabras del mediterráneo provocando un pequeño retraso en la reactivación de la actividad reproductiva durante la temporada de reproducción natural (Zarazaga et al., 2009). De igual manera, se ha observado un intervalo de 2-3 meses a partir del solsticio de verano para la reanudación de la actividad ovárica en cabras expuestas ya sea a un foto periodo natural o a un ciclo controlado de fotoperiodo artificial con una duración de 6 a 12 meses (Escobar, 1997). Por ellos, "el efecto macho" puede aumentar la actividad reproductiva de las cabras criollas mexicanas a pesar de la influencia del fotoperiodo.
El presente trabajo evaluó el efecto de la presencia o la ausencia de un macho cabrío sexualmente activo en la actividad ovárica (lútea) y la sucesión del ciclo estral en ovejas criollas expuestas a una alimentación y a un fotoperiodo controlado con duración de 6 meses en el Centro-Norte de México. Por otra parte, dentro de cada grupo experimental se agregó un grupo de cabras ovariectomizadas (OVX) y un grupo de cabras ovariectomizadas con implante de estradiol (OVX + E2) con el fin de determinar el efecto macho sobre los cambios en la respuesta neuroendocrina que conforma el principal determinante neuroendocrino del estado reproductivo estacional.
MATERIALES Y METODOS
Ubicación, animales y tratamientos
Este estudio se llevó a cabo en la Universidad de Zacatecas, Centro-Norte de México, latitud norte 23° 00' y longitud oeste 102° 44' y a 2,150 m sobre el nivel del mar. El experimento incluyo a cabras criollas, (n=30) provenientes de una región que se localiza en las zonas áridas y semiáridas del estado de Zacatecas en el Centro norte de México. Estas cabras no preñadas cuentan con un peso promedio de 39 ± 4.5 kg. Además, se introdujo en el estudio un macho cabrío sexualmente activo. Los animales se colocaron dentro de cámaras fotoperiódicas controladas (es decir, controlando las horas de luz por día) con 350 lux directamente a la altura de los ojos de las cabras. Todas las cabras fueron expuestas a un fotoperiodo artificial controlado alterno (ascendente/descendente) en un rango de 13.4 a 10.6 horas de luz por día durante cada 90 d-ciclo hasta alcanzar los 6 ciclos fotoperiódicos: ascendente (n=3); y descendiente (n=3). El principal objetivo fue simular los acontecimientos de dos fotoperiodos naturales consecutivos durante un año, y evaluar el efecto de un fotoperiodo específico sobre la función reproductiva de las cabras. Tanto las hembras como los machos fueron alimentados con una ración que cubrió sus necesidades y mantuvo su condición corporal por arriba del 3 (es decir, una escala de 0 a 5).
Diseño experimental
Se formaron dos grupos de cabras al azar: un grupo con cabras no expuestas a la presencia de un macho cabrío sexualmente activo (grupo control, n=15) y el otro, con cabras expuestas a un macho cabrío sexualmente activo (grupo experimental, n=15). Cada grupo (n=15), estaba conformado por cabras ovariectomizadas (cabras OVX, n=5), cabras ovariectomizadas y con implante de estradiol (cabras OVX+E2, n=5), y cabras ovariointactas o cabras control (CG, n=5). Ambos grupos fueron expuestos a un fotoperiodo artificialmente controlado. En consecuencia, se incluyeron las variaciones anuales en horas luz por día en un fotoperiodo mensual artificialmente controlado con el fin de obtener dos ciclos fotoperiódicos al año y evaluar las variaciones en respuesta a la presencia o ausencia de un macho cabrío sexualmente activo. Así mismo, el macho cabrío también fue expuesto a un tratamiento fotoperiódico corto-largo mensual alterno (luz-oscuridad) [foto periodo largo: 16 h L/8 h D; y fotoperiodo corto: 8 h L/16 h D] como previamente se comentó por Delgadillo et al., (1995). Se introdujo un macho cabrío sexualmente activo al grupo experimental (cabras expuestas al macho cabrío), el cual estuvo presente durante el segundo ciclo fotoperiódico y permaneció con las cabras durante el día dependiendo del tratamiento fotoperiódico recibido. Las cabras testigo se mantuvieron permanentemente sin la presencia de un macho.
Recolección de muestras de sangre
Se recolectaron muestras de sangre dos veces por semana para determinar los niveles de progesterona (P4). Estas muestras se obtuvieron por punción de vena yugular, utilizando tubos Vacutainer, y después fueron centrifugadas (1,500 x g, 15 min, 4 °C). Se separó el suero sanguíneo y éste fue almacenado a una temperatura de -20 °C, hasta que se realizó un radioinmunoanálisis.
Las muestras con niveles de progesterona por arriba del 1 ng/mL en el suero sanguíneo fueron consideradas como evidencias de un cuerpo lúteo activo y por lo tanto, de las ovulaciones anteriores. Se tomaron muestras de sangre cada 4 semanas durante un muestreo intensivo (6 h, cada 15 min; 24 muestras por cabra) para la determinación de la LH. Se obtuvieron muestras por punción de vena yugular usando tubos Vacutainer y después fueron centrifugadas (1,500 x g, 15 min, 4 °C). Se separó el suero sanguíneo y fue almacenado a una temperatura de -20 °C hasta que se llevó a cabo un radioinmunoanálisis.
Radioinmunoensayo de progesterona
Los niveles de progesterona fueron determinados por un radioinmunoensayo en fase sólida (Coat- A-Count, Los Angeles, CA). El ensayo de sensibilidad fue de 0.01 ng/mL y los coeficientes de variación intra e inter análisis fueron de 6.7 % y 10.6 %, respectivamente. Las muestras con niveles de progesterona por arriba de 1 ng/mL de suero sanguíneo fueron consideradas como evidencia de la presencia del cuerpo lúteo y el acontecimiento de ovulaciones anteriores.
Radioinmunoensayo de hormona luteinizante (LH)
Las concentraciones de la hormona luteinizante fueron determinadas por un radioinmunoensayo en fase liquida en tubos por triplicado. El ensayo sensibilidad de la LH fue de 0.1 ng/mL, y los coeficientes de variación intra e inter análisis fueron de 6.2 % y 7.08 %, respectivamente. La frecuencia de pulso, la amplitud y la concentración de la LH fueron determinadas mediante el programa Pulsar, el cual fue desarrollado por Merina y Watcher (1982) y reportado por Hoefler y Hallford, (1987).
Análisis estadísticos
Los análisis estadísticos se llevaron a cabo por medio del procedimiento GLM de SAS utilizando un modelo de efectos fijos (SAS, 2000). Se realizó una prueba de Tukey para determinar las diferencias de la media.
RESULTADOS Y DISCUSIÓN
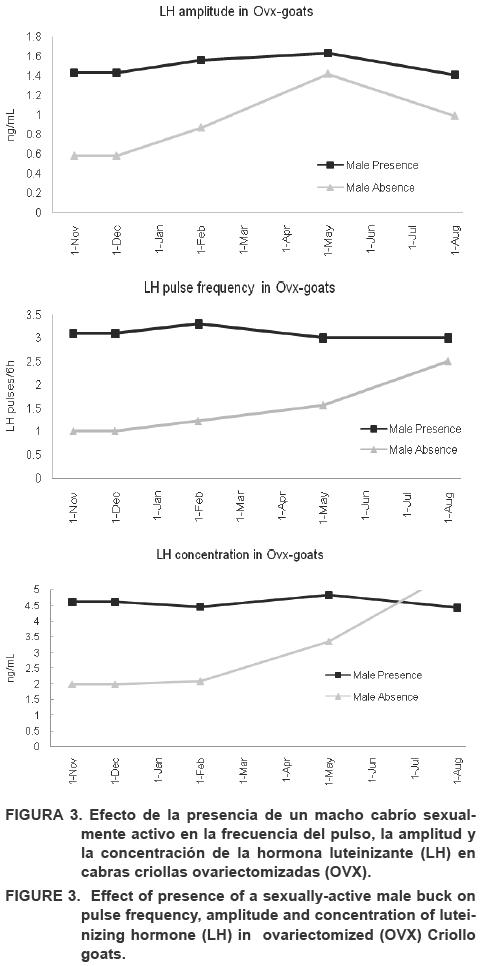
La presencia de un macho cabrío sexualmente activo provocó una reducción en la temporada de anestro de las cabras criollas, así como, el aumento en el número de días con actividad luteal, y por consecuencia, un aumento en el número de fases luteales con tendencia a altos niveles de progesterona durante un ciclo fotoperiòdico artificial descendiente como ocurre de forma natural durante el solsticio de invierno (Figura 1). Cuando se introdujo un macho cabrío sexualmente activo estos efectos permanecieron por más tiempo a pesar del ciclo fotoperiòdico artificial ascendiente (solsticio de verano), el cual supone ser un periodo de inactividad reproductiva. Un gran número de cabras permaneció con el ciclo mientras que se observó una mayor sucesión en el ciclo estral (Figure 1-2).

Un macho cabrío se vuelve sexualmente activo al ser expuesto a tratamientos fotoperiódicos alternos mensuales (16 h L:8 h D; y viceversa; Delgadillo et al., 2001; 2004a; 2004b; 2006). En este estudio, las cabras implantadas con E2, expuestas a un macho cabrío sexualmente activo, mostraron un aumento en la secreción de la LH (LH pulsos/6 h), algo similar ocurrió durante la temporada reproductiva. Por lo tanto, la retroalimentación negativa del estradiol (E2) en el hipotálamo estuvo ausente durante los días de fotoperiodo ascendente (Karsch et al., 1984). La concentración de la LH, la frecuencia de los pulsos y la amplitud de las cabras OVX implantadas con E2 fue similar a aquellas cabras que estaban en estacionalidad reproductiva (Chemineau, 1983; Chemineau et al., 1988, Henniawati et al., 1995; Restall, 1992). En general, la presencia de un macho cabrío sexualmente activo aumenta los niveles en suero de la LH (Cuadro 1).
Niveles de hormona en la LH
La presencia de un macho cabrío sexualmente activo aumenta significativamente (P<0.01), la amplitud, la concentración y la frecuencia del pulso de la LH en cabras OVX expuestas a un fotoperiodo artificial. Sin embargo, no se mostraron variaciones en la LH dentro de los tratamientos fotoperiódicos artiiciales (Cuadro 1-4) (2-3).
Cabras ovariectomizadas
Las cabras OVX expuestas a la presencia de un macho cabrío presentaron un aumento mayor en la frecuencia del pulso (pulsos/ 6 h), la amplitud y la concentración, (P<0.01) de la LH, que las cabras no expuestas a un macho cabrío (Frecuencia: 3.2 ± 0.4 vs. 0.7 ± 0.1 pulsos de la LH cada 6 h; Amplitud: 1.6 ± 0.1 vs. 0.8 ± 0.3 ng/mL; Concentración: 5.3 ± 0.6 vs.2.0 ± 0.9 ng/mL). Véanse los Cuadros 1-4.
Las cabras OVX mostraron una disminución en los niveles de la LH. No obstante, cuando a las cabras se les implanto E2, los niveles de la LH aumentaron (frecuencia del pulso, amplitud y concentración), mientras que la presencia de un macho cabrío sexualmente activo aumento considerablemente los niveles de la LH (P<0.01). Por ello, parece ser que el estradiol ovárico juega un papel importante en una retroalimentación positiva y en la activación del eje hipotálamo-hipofisario-gonadal subsecuente a la temporada de anestro en cabras criollas. Sin embargo, el estímulo E2 fue insuficiente debido a que tal efecto fue provocado solo cuando un macho sexualmente activo se encontraba presente. Tal efecto podría ser un efecto sinérgico en la actividad reproductiva de las cabras criollas, el cual parece haber desarrollado mecanismos adaptativos para solucionar de mejor manera los retos ambientales (Meza-herrera et al., 2006, 2007).
Cabras OVX e implantadas con E2
Las cabras OVX e implantadas con E2 (OVX+E2) expuestas a un macho cabrío sexualmente activo solo generan diferencias en la frecuencia del pulso de LH (LH pulsos/6 h) comparadas con cabras no expuestas a un macho cabrío (P<0.05) (Cuadro 2). De hecho, no se observaron diferencias significativas (P>0.05) en relación a la amplitud de la LH (1.2 ± 0.2 vs.1.1 ± 0.5 ng/ mL), o a la concentración (3.6 ± 0.1 vs. 3.4 ± 1.3 ng/mL), (Cuadros 3-4).
Cabras testigo
Las cabras testigo (cabras ovariointactas), expuestas a la presencia de un macho cabrío sexualmente activo presentaron una actividad luteal mayor (Figuras 1-2), así como una sucesión mayor en los ciclos estrales que las cabras no expuestas a un macho cabrío (Figuras 1-2). Esta conducta reproductiva fue evidente debido a un aumento en el número de días en actividad luteal (44 ± 9.05 vs. 32.3 ± 20.6; P<0.05), una disminución en el número de días en anestro (4.0 ± 4.8 vs. 51.9 ± 28; P<0.05), una disminución en la secreción de progesterona (6.8 ± 0.8 vs. 7.3 ± 0.5; P<0.05), así como un aumento en la fases luteales (15.7 ± 4.4 vs. 11.3 ±8.7); (P<0.05) (Figuras 1-2).
Las cabras testigo no expuestas a machos mostraron una actividad reproductiva solo durante la influencia de los ciclos fotoperiódicos artificiales en el solsticio de invierno (disminución del fotoperiodo con menos horas luz durante el día). Por otro parte, las cabras expuestas a la presencia de un macho cabrío sexualmente activo mostraron de manera práctica una actividad reproductiva durante todo el periodo experimental, independientemente de la influencia de un fotoperiodo artificial reducido o prolongado (fotoperiodo en el solsticio de verano e invierno, respectivamente). Por consiguiente, la presencia de un macho cabrío sexualmente activo en las cabras indujo a una actividad reproductiva mayor (ovárica y/o luteal) a comparación de las cabras que no fueron expuestas a la presencia de un macho. Esta actividad reproductiva fue medida por un mayor número de ciclos estrales, y una mayor sucesión en los ciclos estrales, incluso durante los periodos de actividad reproductiva reducida como en los fotoperiodo mayores (días largos). Se observó refractariedad durante el primer ciclo fotoperiódico artificial pero no en el segundo cuando hubo ausencia del macho dentro del grupo.
Los ciclos fotoperiódicos artificiales (con una duración de 6 meses) fueron implementados para simular dos fotoperiodos naturales en un año, alternando ciclos fotoperiódicos ascendentes y descendentes (Escobar et al., 1997). El rango de horas luz por día que se utilizó en este estudio fue de 13.4 a 10.6 h L/d y fue determinado basándose en estudios previos donde se utilizaron cabras del mismo sitio, observando una influencia de fotoperiodo dependiente en la actividad ovárica de estas cabras criollas Escobar et al., 1997). El fotoperiodo artificial descendente que se obtuvo en las cámaras ambientales controladas puede estimular la función reproductiva en cabras criollas. Sin embargo, durante la temporada reproductiva no hubo continuidad en la sucesión del ciclo estral. La introducción y la presencia de un macho cabrío sexualmente activo aumenta el estímulo de la actividad reproductiva de las cabras. La introducción de un macho durante un fotoperiodo artificial en el solsticio de verano generó actividad ovárica en un intervalo promedio de 12.4 d. Se observaron resultados similares en ovejas (Martin et al., 1986) y cabras bajo foto periodos naturales (Chemineau, 1987), y artificialmente controlados (Chemineau et al., 1986 b).
La disponibilidad de alimentos, aunque no se considera como un factor crítico para la regulación, es un factor modulador en la función reproductiva de las cabras criollas (Urrutia-Morales et al., 2009). No obstante, otros autores han observado un aumento en la actividad reproductiva y en la taza de preñez al implementar un estímulo del "efecto macho" (De Santiago-Miramontes et al., 2008; Fitz-Rodriguez et al., 2009). Así, la presencia de un macho cabrío sexualmente activo permite un aumento positivo en la función reproductiva de cabras nativas independientemente del tratamiento fotoperiódico, tal y como se ha mencionado anteriormente por Malpaux et al. (1997) y Delgadillo et al. (2002). Este efecto parece ser independiente de la proporción macho-hembra (Carrillo et al., 2007) así como la continuidad y la discontinuidad del "efecto macho" (Rivas-Muñoz et al., 2007). A pesar de que, recientemente, ha sido mayormente relacionado con la "innovación" del macho (Delgadillo et al., 2009).
La inducción de la actividad sexual en un macho cabrío ha sido reportada previamente utilizando distintos tratamientos. Uno de estos tratamientos se basa en tratamientos fotoperiódicos alternos (con una duración de dos meses; 16 h L/8 h D) presentado por Delgadillo y Chemineau (1992) en el norte de México (latitud 26° 23'N; longitud 104° 47' O). Delgadillo et al. (1993) también reportó una activación sexual en fotoperiodos controlados y tratamientos de melatonina en machos cabríos (Chemineau et al., 1999; Flores et al., 2000). Para el presente estudio, el macho cabrío recibió un tratamiento fotoperiódico alterno mensual de 16 h L/8 D para prevenir cambios estacionales en el eje hipotálamo-hipofisario-gonadal, y mejorar el peso testicular y la producción de semen (Delgadillo et al., 1995).
La introducción de un macho estimula la sucesión del ciclo estral previniendo o reduciendo el anestro estacional. Con respecto a esto, 5-d después de que se introdujo un macho con las cabras en temporada anovulatoria, aumento la tasa de ovulación a un 2-3 d durante su apogeo (Chemineau, 1983). Para las cabras ovariointactas, el efecto de la presencia de un macho cabrío sexualmente activo aumentó la actividad luteal (44 ± 9.1 vs. 32.3 ±20.6 d), redujo el anestro (4.0 ± 4.8 vs. 51.9 ± 28 d), y no se observaron diferencias en los niveles séricos de progesterona (6.8 ± 0.8 vs.7.3 ± 0.5), sin embargo, se mostró una diferencia notoria en el número de fases luteales observadas (15.7 vs. 11.3). En el presente estudio, las fases luteales aumentaron según lo determinado por la patrones de secreción de progesterona, mostrando una mejor sucesión en el ciclo estral, un aumento en el número de ciclos estrales, así como, un mejor resultado reproductivo; todo esto independientemente del régimen fotoperiódico.
El macho cabrío permaneció de 8 a 10 h por día interactuando con las cabras dependiendo del tratamiento fotoperiódico, provocando una actividad ovárica constante. Así mismo, se ha observado un intervalo de 2-3 meses a partir del solsticio de verano (21 de junio) para la reanudación de la actividad ovárica en cabras expuestas a un fotoperiodo natural o a ciclos controlados de fotoperiodos artificiales con duración de 6 a 12 meses (Escobar et al., 1997). Esto indica que a las cabras les puede tomar de dos o tres meses para desarrollar refractariedad. La presencia de un macho, también induce la actividad reproductiva en cabras hembras del Mediterráneo implantadas con melatonina causando un retraso pequeño en la reactivación de la actividad reproductiva durante la temporada de reproducción natural (Zarazaga et al., 2009).
El tratamiento fotoperiódico artificialmente controlado que se utiliza en cabras parece ser complicado y poco práctico. No obstante, una alternativa podría ser, exponer al macho cabrío a un tratamiento fotoperiódico (16 h L/8 h D), ya que el macho solo requiere de una sola cámara de ambiente controlado. Tales prácticas de manejo representan una verdadera alternativa para las cabras bajo un sistema extensivo de producción. De esta forma la temporada de reproducción es controlada de la forma más rentable y en el momento más óptimo del año.
CONCLUSIONES
La presencia de un macho cabrío sexualmente activo aumentó la actividad ovárica, la actividad en el ciclo estral y la sucesión del ciclo estral en las cabras criollas que se encuentran en condiciones subtropicales, a pesar de la influencia de un fotoperiodo controlado mayor. Además, la presencia de un macho cabrío aumento la amplitud, la concentración y la frecuencia del pulso de la LH en cabras ovariectomizadas independientemente del régimen fotoperiódico. La presencia de un macho ante cabras OVX e implantadas con E2 provocaron un aumento en la frecuencia del pulso de la LH como se observó durante la temporada de reproducción. Por ello, un tratamiento fotoperiódico controlado en machos podría ser una alternativa para programar la temporada de reproducción con ventanas de tiempos mediante la introducción de un macho cabrío sexualmente activo expuesto a periodos alternos de 16 h L; 8 h D. Estas tecnologías reproductivas requieren de un equipo y una infraestructura mínima en un sistema de producción extensivo típico de zonas áridas y semiáridas del mundo.
AGRADECIMIENTOS
Esta investigación fue parcialmente financiada por SIVILLA-CONACYT con el número 1998401010-4.
LITERATURA CITADA
AMOAH, E. A.; BRYANT, M. 1984. A note on the effect of contact with male goats on occurrence of puberty in female goat kids. Anim. Prod. 38: 141-144. [ Links ]
ARÉCHIGA, C. F.; AGUILERA, J. I.; RINCÓN, R. M.; MÉNDEZ DE LARA, S.; BAÑUELOS R.; MEZA-HERRERA, C. A. 2008. Role and perspectives of goat production in a global world. Tropical and Subtrop. Agroecosystems. 9: 1-14. [ Links ]
ARECHIGA-FLORES, C. F.; RINCÓN-DELGADO, R. M. 1998. Perspectives for implementation of reproductive technologies in goats. XIII Annual National Meeting on Goat Production. Universidad Autónoma de San Luis Potosí, San Luis Potosí, S.L.P., México. 13: 12-37. [ Links ]
CARRILLO, E.; VÉLIZ, F. G.; FLORES, J. A.; DELGADILLO, J. A. 2007. A diminution in the male/female ratio does not reduce the ability of sexually active male goats to induce estrus activity in anovulatory female goats. Tec. Pecu. Mex. 45: 319-328. [ Links ]
CHEMINEAU, P. 1983. Effect on oestrus and ovulation of exposing creole goats to the male at three times of the year. J. Reprod. Fertil. 67, 65-72. [ Links ]
CHEMINEAU, P. 1987. Posibilities for using bucks to stimulate ovarian and oestrus cycles in anovulatory goats. A review. Livestock Production Science, 17: 135-147. [ Links ]
CHEMINEAU, P. BARIL, G.; LEBOEUF, B.; MAUREL, M. C.; ROY F.; PELLICER-RUBIO, M.; MALPAUX, B.; COGNIE, Y. 1999. Implications of recent advances in reproductive physiology for reproductive management of goats. J. Reprod. Fertil. Suppl. 54: 129-142. [ Links ]
CHEMINEAU, P.; DAVEAU, A.; COGNIÉ, Y.; AUMONT G.; CHESNEAU, D. 2004. Seasonal ovulatory activity exists in tropical Creole famale goats and Black Belly ewes subjected to a temperature photoperiod. BMC Physiology 4: 1-12. [ Links ]
CHEMINEAU, P.; LEVY, F.; THIMONIER, J. 1986a. Effects of anosmia on LH secretion, ovulation and oestrus behaviour induced by males in the anovular Creole goats. Anim. Reprod. Sci. 10: 125-132. [ Links ]
CHEMINEAU, P.; MARTIN G. B.; SAUMANDE, J.; NORMAN, T. E. 1988. Seasonal and hormonal control of pulsatile LH secretion in the dairy goat (Capra hircus). J. Reprod. Fertil. 83: 91-98. [ Links ]
CHEMINEAU, P.; NORMAN, T. E.; RAVAULT, J. P.; THIMO-NIER, J. 1986b. Induction and persistence of pituitary and ovarian activity in the out-of-season lactating dairy goat after a treatment combining a skeleton photoperiod, melatonin and the male effect. J. Reprod. Fertil. 78: 497504. [ Links ]
CHEMINEAU, P.; PELLICER-RUBIO, M. T.; LASSOUED, N.; KHALDI, G.; MONNIAUX, D. 2006. Male-induced short oestrous and ovarian cycles in sheep and goats: a working hypothesis. Review. Reprod. Nutr. Dev. 46, 417429. [ Links ]
DE SANTIAGO-MIRAMOENTES, MALPAUX, B.; DELGADILLO, J. A. 2009. Bodycondition is associated with a shorter breeding season and reduced ovulation rate in subtropical goats. Anim. Reprod. Sci. 114: 175-182. [ Links ]
DE SANTIAGO-MIRAMONTES, M. A.; RIVAS-MUÑOZ, R.; MUÑOZ-GUTIÉRREZ, M.; MALPAUX, B.; SCARAMUZZI, R. J.; DELGADILLO, J. A. 2008. The ovulation rate in anoestrous female goats managed under grazing conditions and exposed to the male effect is increased by nutritional supplementation. Anim. Reprod. Sci. 105: 409-416. [ Links ]
DELGADILLO, J. A.; CARRILLO, E.; MORAN, J.; DUARTE, G.; CHEMINEAU, P.; MALPAUX, B. 2001. Induction of sexual activity of male creole goats in subtropical northern Mexico using long days and melatonin. J. Anim. Sci. 79, 2245-2252. [ Links ]
DELGADILLO, J. A.; CHEMINEAU, P. 1992. Abolition of the seasonal release of luteinizing hormone and testosterone in Alpine goats (Capra hircus) by short photoperiodic cycles. J. Reprod. Fertil. 94: 45-55. [ Links ]
DELGADILLO, J. A.; CORTEZ, M. E.; DUARTE, G.; CHEMINEAU, P.; MALPAUX, B. 2004a. Evidence that the photoperiod controls the annual changes in testosterone secretion, testicular and body weight in subtropical male goats. Reprod. Nutr. Dev. 44: 183-193. [ Links ]
DELGADILLO, J. A.; FITZ-RODRÍGUEZ, G.; DUARTE, G.; VÉLIZ, F. G.; CARRILLO, E.; FLORES, J. A.; VIELMA, J.; HERNANDEZ, H.; MALPAUX, B. 2004b. Management of photoperiod to control caprine reproduction in the subtropics. Reprod. Fertil. Dev. 16: 471-478. [ Links ]
DELGADILLO, J. A.; FLORES, J. A.; VÉLIZ, F. G.; DUARTE, G.; VIELMA, J.; HERNÁNDEZ, H.; FERNÁNDEZ, I. G. 2006. Importance of the signals provided by the buck for the success of the male effect in goats. Review. Reprod. Nutr. Dev. 46: 391-400. [ Links ]
DELGADILLO, J. A.; FLORES, J. A.; VELIZ, F. G.; HERNÁDEZ, H. F.; DUARTE, G.; VIELMA, J.; POINDRON, P.; CHEMINEAU, P.; MALPAUX, B. 2002. Induction of sexual activity in lactating anovulatory female goats using male goats treated only with artificially long days. J. Anim. Sci. 80: 2780-2786. [ Links ]
DELGADILLO, J. A.; GELEZ, H.; UNGERFELD, R.; HAWKEN, P. A.; MARTIN, G. B. 2009. The 'male effect' in sheep and goats-Revisiting the dogmas. Review. Behav. Brain Res. 2009. 304-314. [ Links ]
DELGADILLO, J. A.; HOCHEREAU-DE REVIERS, M. T.; DAVEAU, A.; CHEMINEAU, P. 1995. Effect of short photo-periodic cycles on male genital tract and testicular parameters in male goat (Capra hircus). Reprod. Nutr. Dev. 35: 549-558. [ Links ]
DELGADILLO, J. A.; LEBOEUF, B.; CHEMINEAU, P. 1993. Maintenance of sperm production in bucks during a third year of short photoperiodic cycles. Reprod. Nutr. Dev. 33: 609-617. [ Links ]
DELGADILLO-SÁNCHEZ, J. A.; FLORES-CABRERA, J. A.; VÉLIZ-DERAS, F. G.; DUARTE-MORENO, G.; VIELMA-SIFUENTES, J.; POINDRON-MASSOT P.; MALPAUX, B. 2003. Control de la reproducción de los caprinos del subtrópico mexicano utilizando tratamientos fotoperiódi-cos y efecto macho. Vet. Mex. 34: 69-79. [ Links ]
EBLING, F. J. 2005. The neuroendocrine timing of puberty. Reproduction. 129: 675-683. [ Links ]
ESCOBAR, F. J.; ZARCO L.; VALENCIA, J. 1997. Effect of photoperiod on reproductive seasonality of criollo goat in Mexico. XXII Anual Meeting of Academy for Reproductive Biology Research. Acapulco, Guerrero, México. [ Links ]
FITZ-RODRÍGUEZ, G.; DE SANTIAGO-MIRAMONTES, M. A.; SCARAMUZZI, R. J.; MALPAUX, B.; DELGADILLO, J. A. 2009. Nutritional supplementation improves ovulation and pregnancy rates in female goats managed under natural grazing conditions and exposed to the male effect. Anim. Reprod. Sci. 116: 85-89. [ Links ]
FLORES, J. A.; VELIZ, F. G.; PÉREZ-VILLANUEVA, J. A.; MARTÍNEZ DE LA ESCALERA, G.; CHEMINEAU, P.; POINDRON, P.; MALPAUX, B.; DELGADILLO, J. A. 2000. Male reproductive conditions is the limiting factor of efficiency in the male effect during seasonal anestrus in female goats. Biol. Reprod. 62: 1409-1414. [ Links ]
GÁMEZ-VÁZQUEZ, H. G.; ROSALES-NIETO, C. A.; BAÑUELOS-VALENZUELA, R.; URRUTIA-MORALES, J.; DÍAZ-GÓMEZ, M. O.; SILVA-RAMOS, J. M.; MEZA-HERRERA, C. A. 2008. Body condition score positively influence plasma leptin concentrations in criollo goats. J. Anim. Vet. Adv. 7, 1237-1240. [ Links ]
GELEZ H.; FABRE-NYS, C. 2004. The "male effect" in sheep and goats: A review of the respective roles of the two olfactory systems. Horm. and Behav. 46: 257-271. [ Links ]
GUERRA-GARCÍA, M.; MEZA-HERRERA, C. A.; SÁNCHEZ-TORRES-ESQUEDA, M. T.; GALLEGOS-SÁNCHEZ, J.; TORRES-HERNÁNDEZ, G.; PRO-MARTÍNEZ, A. 2009. IGF-1 and ovarian activity of goats in divergent body condition and supplemented with non-degradable ruminal protein. Agrociencia. 43, 241-247. [ Links ]
HENNIAWATI, H.; RESTALL, B. J.; SCARAMUZZI, R. J. 1995. Effect of season on LH secretion in ovariectomized Australian cashmere does. J. Reprod. Fertil. 103: 349-356. [ Links ]
HOEFLER, W. C.; HALLFORD, D. M. 1987. Influence of suckling status and type of birth on serum hormone profiles and return to estrus in early-postpartum, spring-lambing ewes. Theriogenology 27: 887-895. [ Links ]
KARSCH, F. J.; BITTMAN, E. L.; FOSTER, D. L.; GOODMAN, R. L.; LEGAN, S. J.; ROBINSON, J. E. 1984. Neuroendocrine basis of seasonal reproduction. Rec. Progress in Horm. Res. 40: 185-232. [ Links ]
MALPAUX, B.; VIGUIÉ, C.; SKINNER, D. C.; THIÉRY, J. C.; CHEMINEAU, P. 1997. Control of the circannual rhythm of reproduction by melatonin in the ewe. Brain Res. Bull. 44: 431-438. [ Links ]
MARTIN, G.; OLDMAN, C.; COGNIE, Y.; PEARCE, D. 1986. The physiological responses of anovulatory ewes to the introduction of rams. Review. Livest. Prod. Sci. 15: 219-247. [ Links ]
MELLADO, M.; OLIVAS, R.; RUIZ, F. 2000. Effect of buck stimulus on mature and pre-pubertal norgestomet-treated goats. Sm. Rum. Res. 36: 269-274. [ Links ]
MEZA-HERRERA, C. A.; SÁNCHEZ, J. M.; CHÁVEZ-PERCHES, J. G.; SALINAS, H.; MELLADO, M. 2004. Protein supplementation, body condition and ovarian activity in goats. Preovulatory serum profile of insulin. South Afric. J. Animal Sci. 34 Suppl. 1: 223-226. [ Links ]
MEZA-HERRERA, C. A.; MARTÍNEZ, L.; ARÉCHIGA, C.; BA-ÑUELOS, R.; RINCÓN, R. M.; URRUTIA, J.; SALINAS, H.; MELLADO, M. 2006. Circannual identification and quantification of constitutive heat shock proteins (HSP 70) in goats. J. Appl. Anim. Res. 29: 9-12. [ Links ]
MEZA-HERRERA, C. A.; BOCANEGRA, J. A.; BAÑUELOS, R.; ARÉCHIGA, C. F.; RINCÓN, R. M.; OCHOA-CORDERO, M. A.; JUÁREZ-REYES, A. S.; CERRILLO-SOTO M. A.; SALINAS, H. 2007. Circannual fluctuations in serum cortisol and glucose concentrations and hair coat growth in goats. J. Appl. Anim. Res. 31: 79-82. [ Links ]
MEZA-HERRERA, C. A.; ROSS, T.; HALLFORD, D. M.; HAWKINS, D.; GONZÁLEZ-BULNES, A. 2007. Effects of body condition and protein supplementation on LH secretion and luteal function in sheep. Reprod. Dom. Anim. 42: 461-465. [ Links ]
MEZA-HERRERA, C. A.; HALLFORD, D. M.; ORTIZ, J. A.; CUEVAS, R. A.; SÁNCHEZ, J. M.; SALINAS, H.; MELLADO, M.; GONZÁLEZ-BULNES, A. 2008. Body condition and protein supplementation positively affect perio-vulatory ovarian activity by non-LH mediated pathways in goats. Anim. Reprod. Sci. 106: 412-420. [ Links ]
MEZA-HERRERA, C. A.; VELIZ-DERAS, F. G.; WURZINGER, M.; LÓPEZ-ARIZA, B.; ARELLANO-RODRÍGUEZ, G.; RODRÍGUEZ-MARTÍNEZ, R. 2010a. The kiss-1-kisspep-tin-gpr54 complex: A critical modulator of GnRH neurones during pubertal activation. J. Appl. Biomed. 8: 1-9. [ Links ]
MEZA-HERRERA, C. A.; GONZÁLEZ-BULNES, A.; KRIDLI, R.; MELLADO, M.; ARECHIGA-FLORES, C. F.; SALINAS, H.; LUGINBHUL, J. M. 2010b. Neuroendocrine, metabolic and genomic cues signaling the onset of puberty in females. Reprod. Dom. Anim. doi: 10.1111/j.1439-0531-.2009.01355.x [ Links ]
PELLICER-RUBIO, M. T.; LEBOEUF, B.; BERNELAS, D.; FORGERIT, Y.; POUGNARD, J. L.; BONNÉ, J. L.; SEN-TY, E.; CHEMINEAU, P. 2007. Highly synchronous and fertile reproductive activity induced by the male effect during deep anoestrus in lactating goats subjected to treatment with artificially long days followed by a natural photoperiod. Anim. Reprod. Sci. 98: 241-258. [ Links ]
RESTALL, B. J. 1992. The male effect in goats. In: WILSON, R. T.; BOURZA, T. D. Editors, V International Conference on Goats. New Delhi, India. [ Links ]
RIVAS-MUÑOZ, R.; FITZ-RODRÍGUEZ, G.; POINDRON, P.; MAL-PAUX, B.; DELGADILLO, J. A. 2007. Stimulation of estrous behaviour in grazing female goats by continuous or discontinuous exposure to males. J. Anim. Sci. 85: 1257-1263. [ Links ]
SAS. 2000. SAS/STAT® User's Guide (8.1 Edition). SAS Inst. Inc. Cary, NC, USA. [ Links ]
SCARAMUZZI, R. J.; CAMPBELL, B. K.; DOWNING, J. A.; KENDALL, N. R.; KHALID, M.; MUÑOZ-GUTIÉRREZ, M.; SOMCHIT, A. 2006. A review of the effects of supplementary nutrition in the ewe on the concentrations of reproductive and metabolic hormones and the mechanisms that regulate folliculogenesis and ovulation rate. Reprod. Nutr. Dev. 46: 339-354. [ Links ]
SCARAMUZZI, R. J.; MARTIN G. B. 2008. The importance of interactions among nutrition, seasonality and socio-sexual factors in the development of hormone-free methods for controlling fertility. Reprod. Domest. Anim. 43: 129-136. [ Links ]
UNGERFELD, R.; FORSBERG, M.; RUBIANES, E. 2004. Overview of the response of anoestrous ewes to the ram effect. Reprod. Fertil. Dev. 16: 479-490. [ Links ]
URRUTIA-MORALES, J.; MEZA-HERRERA C. A.; ESCOBAR-MEDINA, F. J.; GÁMEZ-VÁZQUEZ, H. G.; RAMÍREZ-ANDRADE, B. M.: DÍAZ-GÓMEZ M. O.; GONZÁLEZ-BULNES, A. 2009. Relative roles of photoperiodic and nutritional cues in modulating ovarian activity in goats. Reprod. Biol. 9: 283-294. [ Links ]
VALENCIA, J.; GONZÁLEZ, J. L.; DÍAZ, J. 1986. Actividad reproductiva de la cabra criolla en México en el examen postmortem del aparato genital. Vet. Méx. 17: 177-189. [ Links ]
VALENCIA, J.; ZARCO, L.; DUCOING, A.; MURCIA, C.; NAVARRO, H. 1990. Breeding season of criollo and granadina goats under constant nutritional levels in the Mexican highlands. In: Livestock Reproduction in Latin America. International Atomic Energy Agency, Viena. 321-333. [ Links ]
VELIZ, F. G.; MORENO, S.; DUARTE, G.; VIELMA, J.; CHEMINEAU, P.; POINDRON, P.; MALPAUX, B.; DELGADILLO, J. A. 2002. Male effect in seasonally anovulatory lactating goats depends on the presence of sexually active bucks, but not estrous females. Anim. Reprod. Sci. 72: 197-207. [ Links ]
VELIZ, F. G.; POINDRON, P.; MALPAUX, B.; DELGADILLO, J. A. 2006a. Maintaining contact with bucks does not induce refractoriness to the male effect in seasonally anestrous female goats. Anim. Reprod. Sci. 92: 300-309. [ Links ]
VELIZ, F. G.; POINDRON, P.; MALPAUX, B.; DELGADILLO, J. A. 2006b. Positive correlation between the liveweight of anestrous goats and their response to the male effect with sexually active bucks. Reprod. Nutr. Dev. 6: 1-6. [ Links ]
WALKDEN-BROWN, S. W.; MARTIN, G. B.; RESTALL, B. J. 1999. Role of male-female interaction in regulating reproduction in sheep and goats. J. Reprod. Fertil. Suppl. 52: 243-257. [ Links ]
ZARAZAGA, L. A.; GATICA, M. C.; CELI, I.; GUZMAN, J. L.; MALPAUX B. 2009. Effect of melatonin implants on sexual activity in Mediterranean goat females without separation from males. Theriogenology 72: 910-918. [ Links ]

