










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.17 no.spe Chapingo ene. 2011
https://doi.org/10.5154/r.rchscfa.2010.09.081
La presencia de hembras estrogenizadas al momento del efecto macho induce la actividad estral de cabras en el semidesierto mexicano
The presence of estrogenized females at the moment of male effect induce oestrous activity of goats in the mexican semi-desert
María de los Ángeles De Santiago-Miramontes1; Sandra Marcelino-León1; Juan Ramón Luna-Orozco2; Raymundo Rivas-Muñoz3; Rafael Rodríguez-Martínez1; Miguel Mellado-Bosque1; Francisco Gerardo Véliz-Deras1¶
1 Universidad Autónoma Agraria Antonio Narro, Periférico Raúl López Sánchez y Carretera a Santa Fe, Torreón, Coahuila. C. P. 27054, MÉXICO. Correo- e: velizderas@yahoo.com (¶Autor para correspondencia).
2 Centro de Bachillerato Tecnológico Agropecuario Núm. 1, Carretera Torreón — La Partida km 14.5, Torreón, Coahuila, MÉXICO.
3 Instituto Tecnológico de Torreón, Carretera Torreón-San Pedro km 7.5, Torreón, Coahuila, México.
Recibido: 22 de septiembre, 2010
Aceptado: 07 de marzo, 2011
RESUMEN
El objetivo del presente estudio fue determinar si la presencia de hembras estrogenizadas y machos sexualmente inactivos estimula la actividad estral de las cabras anovulatorias del norte de México, y si el porcentaje de hembras estrogenizadas afecta esta respuesta a mitad del anestro estacional. Se utilizaron tres grupos (n=31, c/u) de hembras anovulatorias expuestas a dos machos/grupo. Un grupo fue expuesto además a seis hembras estrogenizadas (G20 %), un segundo grupo fue expuesto a tres hembras estrogenizadas (G10 %), y el tercer grupo fue expuesto a seis hembras no estrogenizadas (G0 %). El porcentaje total de cabras que presentaron actividad estral durante el estudio fue similar entre los grupos G10 y G20 % (93 y 90 %, respectivamente, P>0.05), pero diferente en el grupo G0 % donde ninguna hembra presentó estro (P<0.01). El inicio de la actividad estral fue más tardía en el G10 % (167.8±17 h) que en el G20 % (142±20 h; P<0.001). Estos resultados indican que la presencia de hembras estrogenizadas al momento del efecto macho, estimula la actividad estral de las hembras anovulatorias locales del semidesierto mexicano, y además, que la presencia de un 20 % de hembras estrogenizadas reduce el lapso entre la introducción de machos + hembras estrogenizadas, y el inicio de la actividad estral o latencia al primer estro.
Palabras clave: Efecto hembra, anestro estacional, actividad sexual.
ABSTRACT
The objective of this study was to determine whether the presence of estrogenized females and sexually inactive males stimulates the estrous activity of anovulatory goats, and if the percentage of females estrogenized affects this response. Three groups (n = 31 each) of anovulatory females exposed to two males/group were used. The first group was exposed to six estrogenized females (G20 %), a second group was exposed to tree estrogenized females (G10 %), and the third group was exposed to six non estrogenized females (GO %). The total percentage of goats in estrus during the study was similar between the groups G20 and G20 % ( 93 and 90 %, respectively, P>0.05), but different in the group G0 %, where any female was in estrus (P<0.01). The onest of estrous activity was slower in the G10 % (167.8±17 h) than in the G20 % (142 ± 20 h, P<0.001). Our results suggest that at the time of the male effect, the presence of estrogenized females stimulates the estrus activity of the native anovulatory females from the Mexican semi-desert, and that the presence of 20 % of estrogenized females reduces the period between male introduction + estrogenizzed females regarding the onset of estral activity or the latency to the first estrus.
Key words: Famale effect, seasonal anestrous, sexual activity.
INTRODUCCIÓN
La estacionalidad reproductiva es una característica de la mayoría de las razas de ovejas y cabras de latitudes templadas y subtropicales (Chemineau et al., 1992; Aréchiga et al., 2008; Delgadillo, 2010; Carrillo et al., 2010). Durante una temporada del año, las cabras manifiestan celos con ovulaciones fértiles (Shelton, 1978), esta condición provoca que la producción (leche y cabritos) sea también estacional, lo que representa un serio problema de comercialización para el productor (Álvarez et al., 1999; González-Bulnes et al., 2011). Con el objetivo de contrarrestar esta estacionalidad se han desarrollado diversos métodos, entre los que destacan los tratamientos fotoperiódicos, los hormonales, los socio-sexuales y los nutricionales (Meza-Herrera et al., 2004, 2007, 2008; Leboeuf et al., 2008; Guerra-García et al., 2009; De Santiago-Miramontes et al., 2008, 2009; Urrutia-Morales et al., 2009). Un método socio-sexual probado en el semidesierto mexicano es el denominado "efecto macho" (Véliz et al., 2002, 2006, 2009). Este método consiste en la introducción repentina de un macho en un grupo de hembras anovulatorias, promoviendo su actividad estral y ovárica en los días subsiguientes (Urrutia-Morales et al., 2000; Martínez et al., 2005; Véliz et al., 2009; Delgadillo, 2010; Flores-Najera et al., 2010; Rosales-Nieto et al., 2011). Sin embargo, un factor muy importante que influye en la respuesta de las hembras al efecto macho es el comportamiento sexual de los machos (Rivas-Muñoz et al., 2010). En efecto, las cabras anovulatorias del semidesierto del norte de México reinician su actividad estral y ovulatoria si son expuestas a machos cabríos inducidos a una intensa actividad sexual (olor, comportamiento sexual y vocalizaciones) mediante un tratamiento de 2.5 meses de días largos artificiales, mientras que cuando son expuestas a machos en reposo sexual (bajo las variaciones naturales del fotoperiodo de la región), la respuesta estral y ovulatoria es menor al 10 % (Véliz et al., 2002; Rivas-Muñoz et al., 2010). Otro factor que puede influir en la respuesta al efecto macho es la presencia de hembras en celo al momento de la introducción de los machos. La exposición de carneros (Rosa et al., 2000) o machos cabríos (Walkden-Brown et al., 1993) a hembras en celo antes o en el momento de la introducción de estos machos a un grupo de hembras anovulatorias, incrementa la proporción de hembras inducidas a la actividad sexual. En efecto, el 87 % de las cabras Cashmere australianas respondieron al efecto macho cuando al momento de la introducción de los machos estaban presentes hembras en estro, en contraste con un 72 % cuando existió ausencia de éstas (Walkden-Brown et al., 1993). Probablemente esto se deba a que las hembras en estro pueden inducir la actividad sexual (mediante su comportamiento sexual y feromonas) de los machos, lo que a su vez provoca una mayor estimulación sexual de las hembras en anestro (Walkden-Brown et al., 1993). Sin embargo, no se ha probado si la introducción de hembras en estro, al momento del empadre, puede estimular la actividad estral y ovulatoria de las cabras locales a la mitad del anestro en el semidesierto del norte de México, dada su marcada estacionalidad (Véliz et al., 2006). De tener éxito, se habrá encontrado un método efectivo y sencillo para controlar la actividad reproductiva de los caprinos del semidesierto del norte de México. El objetivo del presente estudio fue determinar si el estímulo ejercido por la presencia de machos + hembras estrogenizadas, estimula la actividad estral de las cabras anovulatorias explotadas extensivamente en el norte de México, y si el porcentaje de hembras estrogenizadas afecta esta respuesta a mitad del anestro estacional.
MATERIALES Y MÉTODOS
Localización del estudio
El estudio se llevó a cabo del 20 de mayo al 6 de junio del 2010, en el semidesierto del norte de México (26° N), y se utilizaron animales caprinos locales adultos. Los animales locales son descendientes de la cruza de razas españolas como Granadina, Murciana y Malagueña con otras razas europeas como Alpina, Saanen y Anglo-Nubia (Delgadillo et al., 1999). El clima es semi-desértico, la temperatura máxima promedio es de 37 °C en mayo-junio y la mínima de 6 °C en diciembre-enero. El fotoperiodo de la región es de 13 h 41 min luz durante el solsticio de verano y de 10 h 19 min en el solsticio de invierno (Delgadillo et al., 1999). Los machos cabríos locales (criollos) presentan un periodo de reposo sexual de enero a abril, mientras que en las hembras criollas el periodo de anestro sucede de marzo a agosto (Delgadillo, 2010).
Animales experimentales
Machos. Se utilizaron seis machos adultos locales del norte de México, los cuales fueron expuestos a las variaciones naturales del fotoperiodo de la región, y dos meses antes de iniciar el estudio se estabularon y fueron puestos en un corral de 5 x 5 m, y se alimentaron con una dieta que cubría sus necesidades fisiológicas (2 kg de heno de alfalfa, 1 kg de ensilado de maíz, 500 g de maíz rolado, por animal por día, sales minerales y agua a libre acceso).
Hembras. Se utilizaron en total 93 cabras multíparas anovulatorias de la raza local. Las hembras fueron aisladas de los machos dos meses antes del inicio del experimento (20 de mayo; día 0). Los días -21, -14 y -7 todas las hembras fueron sometidas a revisión de actividad ovulatoria mediante ecografía transrectal (Aloka SSD-500). Todas las hembras fueron diagnosticadas anovulatorias. Las hembras antes del estudio eran explotadas en un sistema de libre pastoreo de 10:00 a 18:00 h donde se alimentaban solamente de la flora disponible en la región, consistente predominantemente en arbustos como Prosopis glandulosa, Acacia farneciana, Atriplex acantocarpa, Agave scabra, Mimosa biuncife, herbáceas como Heliantus ciliaris, Salsola kali, Solanum elaeagnilolium y pastos como Sorghum halepense, Chloris virgata, Setaria verticillata, Eragrostis pectinacea, Bouteloua curtipendula, Aristida purpurea y Bouteloua barbata. En ocasiones aprovechaban esquilmos o rastrojos de cultivos agrícolas tales como sorgo (sorghum vulgare) y maíz (zea maiz), entre otros. Dos días antes del inicio del estudio, las cabras fueron divididas en tres grupos (n=31 c/u) homogéneos en cuanto a peso y condición corporal.
Empadre. El 20 de mayo (día 0) cada grupo de hembras fue expuesto a dos machos inactivos sexualmente; además, un grupo fue expuesto a seis hembras estrogenizadas (G20 %), un segundo grupo fue expuesto a tres hembras estrogenizadas (G10 %), y el tercer grupo fue expuesto a seis hembras testigo (no estrogenizadas; G0 %). Los machos y las hembras estrogenizadas estuvieron en contacto con los grupos de hembras anovulatorias durante 17 días. Los grupos estuvieron a una distancia aproximada de 100 m entre ellos. Durante todo el periodo de estudio todos los animales se alojaron en corrales provistos de sombra, y a partir del día 0 se les proporcionó alfalfa henificada, ensilaje de sorgo, sales minerales y agua ad libitum.
Tratamiento de las hembras estrogenizadas. Cada hembra estrogenizada fue tratada con 20 mg de progesterona como dosis única el día -8; esto se realizó con la finalidad de que las hembras sean más receptivas a estradiol y manifiesten el comportamiento estral a corto plazo. Además se les aplicaron 2 mg de cipionato de estradiol por vía intramuscular cada tercer día, iniciando el día -6 y hasta el Anal del estudio.
Variables determinadas. Durante todo el estudio se detectó el estro dos veces al día, de 8:00 a 10:00 h y de 18:00 a 20:00 h. Para ello, dos horas antes, los machos se separaron 50 m de los corrales de las cabras y se introdujeron alternadamente entre grupos. Se consideró una cabra en estro cuando ésta permitió la cópula. Se registró el intervalo entre la introducción de los machos + las cabras estrogenizadas y el inicio del estro de las cabras anéstricas, así como la proporción de cabras que presentaron estro en cada grupo. Se diagnosticó la gestación aproximadamente a los 65 días después de la última monta mediante ecografía transrectal (Aloka SSD-500).
Análisis estadísticos
El intervalo al inicio de la actividad estral fue comparado con la prueba de two-sample t-test. La proporción diaria y acumulada de cabras que presentaron estro, así como la proporción de hembras gestantes, se comparó con una prueba de Chi-squared test usando el programa MYSTAT 12.
RESULTADOS Y DISCUSIÓN
El porcentaje total de cabras en estro durante los 17 días del estudio fue similar en los grupos G10 y G20 % (93 y 90 % respectivamente; P>0.05; Cuadro 1), pero diferente en el grupo G0 % donde ninguna hembra presentó estro (P<0.001). Las proporciones de cabras en estro tanto diariamente como a lo largo del estudio no fueron diferentes entre el G10 y G20 %; sin embargo, fueron diferentes (P<0.001) en el G0 % (Figura 1).

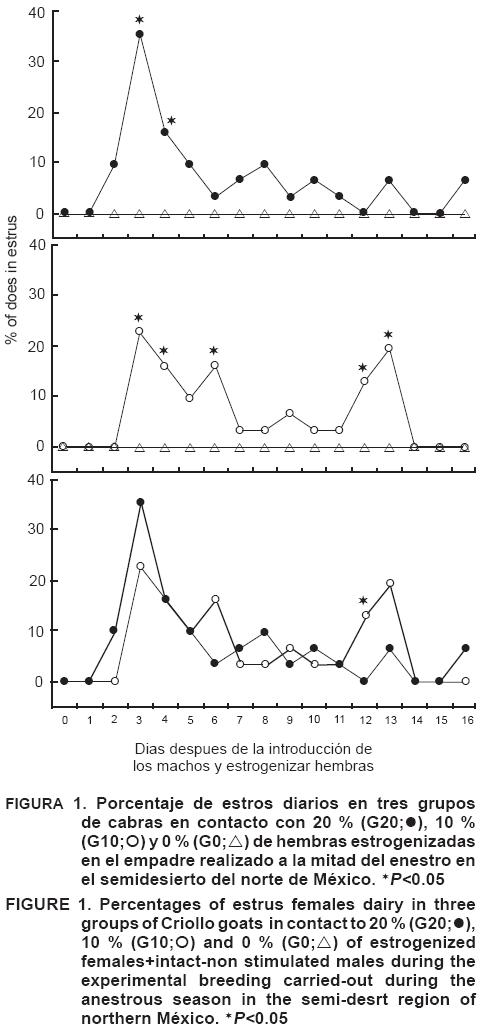
El intervalo entre la introducción de machos + hembras estrogenizadas y el inicio de la actividad estral de los grupos en estudio fue más tardío en el G10 % (167±17 h) que en el G20 % (142±20 h; P<0.001), lo cual indica que el porcentaje de hembras estrogenizadas tiene una influencia directa sobre el inicio de la actividad estral. La proporción de hembras gestantes no fue diferente entre los grupos G10 y G20 % (45 y 48 %, respectivamente; P>0.05), pero sí entre estos dos grupos y el G0 % (0 %; P<0.001). Los resultados del presente estudio demuestran que las hembras estrogenizadas ejercen un efecto de "facilitación social" para inducir la actividad estral de hembras anéstricas en el norte de México. La mayoría de las hembras anovulatorias reiniciaron su actividad estral al ser expuestas a machos + hembras estrogenizadas, mientras que ninguna hembra reinició su actividad estral al ser expuesta solamente a los machos en el periodo de reposo sexual (Figura 1). Además, el 20 % de hembras estrogenizadas acortó la latencia al primer estro, probablemente porque el contacto establecido con las hembras anovulatorias y los machos, provocó una mayor y más rápida respuesta sexual (Álvarez et al. 1999). Por otra parte, la falta de respuesta en las hembras expuestas solamente a machos se debió probablemente a que éstos estaban en reposo sexual, pues se ha reportado que las hembras anovulatorias no reinician su actividad estral al ser expuestas a machos que muestran comportamiento sexual, olor y vocalizaciones bajos (Véliz et al., 2006; Rivas-Muñoz et al., 2010). La alta respuesta estral de los grupos G10 y G20 % (Cuadro 1), contrasta con lo reportado por Véliz et al. (2002), donde la introducción de machos + 20 % de hembras inducidas al estro no estimuló la actividad estral de las hembras anovulatorias. Esta diferencia se debió probablemente a dos factores: 1) La época del año en el que fue realizado el efecto macho; el presente estudio en mayo, y el de Véliz et al. (2002) en marzo; 2) En el presente estudio se utilizaron hembras en estro permanente, mientras que en lo reportado por Véliz et al. (2002) las hembras inducidas al estro mostraron esta conducta solamente 24 h, ya que fueron tratadas con eCG. Probablemente en este estudio la constante presencia de las hembras es-trogenizadas estimuló la actividad sexual y la secreción de feromonas de los machos, provocando que éstos ejercieran una mayor estimulación sexual de las hembras en anestro (Walkden-Brown et al., 1993). Además, la alta respuesta estral de las hembras anovulatorias a la introducción de las hembras estrogenizadas pudo ser una estimulación de forma directa. Se ha reportado que las hembras en estro inducen la actividad estral en sus compañeras anéstricas (Zarco et al., 1995). Por otra parte, solamente la mitad de las hembras fueron diagnosticadas gestantes; esto pudo deberse a la baja calidad y duración del cuerpo lúteo generado después de la inducción de la ovulación (Chemineau et al., 2006). También es probable que la baja condición corporal de las hembras del presente estudio disminuyera la fertilidad. Se ha reportado que cuando se realiza el efecto macho en hembras con baja condición corporal, solamente el 50 % de las hembras queda gestante (Fitz-Rodríguez et al., 2009). Tanto la desnutrición como desbalances en la relación energía-proteína pueden afectar la pérdida embrionaria, probablemente debido a cambios en el ambiente uterino (Abecia et al., 2006; Martin y Kadokawa, 2006; Meza-Herrera et al., 2006, 2010). Otra posibilidad es que estos machos tuvieran una inadecuada calidad seminal, ya que antes del empadre estaban en reposo sexual, y por lo tanto su calidad seminal probablemente era baja (motilidad, viabilidad, número de espermatozoides, etc; Delgadillo et al., 1999; Carrillo et al., 2010). La calidad seminal no se puede mejorar en pocos días; sin embargo, los niveles de LH y testosterona plasmáticos, responsables del nivel de olor y del comportamiento sexual, se elevan inmediatamente después del contacto con hembras en estro (Walkden-Brown et al., 1994). Efectivamente, el comportamiento sexual del macho y su olor son decisivos en la estimulación de la actividad estral y ovulatoria de las hembras (Walkden-Brown et al., 1993); se ha reportado que las hembras durante el anestro estacional solamente responden a la introducción de machos sexualmente activos (Rivas-Muñoz et al., 2010; Delgadillo y Vélez, 2010).
El tiempo de preparación de las hembras estro-genizadas y los machos para el empadre es muy corto (una semana antes y durante el empadre), en contraste con tecnologías que requieren de por lo menos cinco meses de preparación y de una infraestructura eléctrica (Delgadillo y Vélez, 2010) que muchos productores no poseen en sus rústicas explotaciones. Paralelo a ello, los animales estimulados no reciben tratamiento hormonal, lo que convierte a este método en una estrategia limpia, verde y ética (Martin y Kadokawa, 2006). Para incrementar el porcentaje de hembras gestantes al utilizar esta técnica, sería útil complementar este método con uno que permita una mejor calidad seminal en los machos, e investigar si una complementación alimenticia en las hembras incrementa su tasa ovulatoria y fertilidad (De Santiago-Miramontes et al., 2008; Fitz-Rodríguez et al., 2009).
CONCLUSIONES
Estos resultados demuestran que la presencia de cabras estrogenizadas al momento del efecto macho utilizando machos inactivos sexualmente, estimula la actividad estral de las cabras anovulatorias bajo un sistema de producción marginal del semidesierto mexicano. Además, la presencia de 20 % de hembras en estro reduce el lapso entre la introducción de machos + hembras estrogenizadas, y el inicio de la actividad estral. Como esta metodología sólo requiere la presencia de 10 % de hembras estrogenizadas en el rebaño, además de ser muy efectiva desde un punto de vista reproductivo, es de muy bajo costo desde una óptica económica.
AGRADECIMIENTOS
Los autores agradecen a los señores Daniel y Rogelio Alvarado el haber facilitado las instalaciones y los animales experimentales. Asimismo, a Leticia Gaytán Alemán, Giovanni Huerta y Luis Fernando López por su asistencia técnica.
LITERATURA CITADA
ABECIA, J. A.; SOSA, C.; FORCADA, F.; MEIKLE, A. 2006. The effect of undernutrition on the establishment of pregnancy in the ewe. Reproduction Nutrition and Development 46: 367-378. [ Links ]
ÁLVAREZ, L.; DUCOING, A. E.; ZARCO, L. A.; TRUJILLO, A. M. 1999. Conducta estral, concentraciones de LH y función Lútea en cabras en enestro estacional inducidas a ciclar mediante el contacto con cabras en estro. Veterinaria México 30: 25-31. [ Links ]
ARÉCHIGA, C. F.; AGUILERA, J. I.; RINCÓN, R. M.; MENDEZ DE LARA, S.; BAÑUELOS, V. R.; MEZA-HERRERA, C. A. 2008. Role and perspectives of goat production in a global world. Tropical and Subtropical Agroecosystems. 9: 1-14. [ Links ]
CARRILLO, E.; MEZA-HERRERA, C. A.; VÉLIZ, F. G. 2010. Estacionalidad reproductiva de los machos cabríos de la raza Alpino-Francés adaptados al subtrópico mexicano. Revista Mexicana de Ciencias Pecuarias 1: 169-178. [ Links ]
CHEMINEAU, P; PELLICER-RUBIO, M. T.; LASSOUED, N.; KHALDI, G.; MONNIAUX, D. 2006. Male-induced short oestrous and ovarian cycles in sheep and goats: a working hypothesis. Reproduction Nutrition Develop 46(4): 417-429. doi:10.1051/rnd:2006022 [ Links ]
CHEMINEAU, P.; DAVEAU, A.; MAURICE, F.; DELGADILLO, J. A. 1992. Seasonality of estrus and ovulation is not modified by subjecting female Alpine goats to a tropical photoperiod. Small Ruminant Research 8: 299-312. doi:10.1016/0921-4488(92)90211-L [ Links ]
DE SANTIAGO-MIRAMONTES, M. A.; RIVAS-MUÑOZ, R.; MUÑOZ-GUTIÉRREZ, M.; MALPAUX, B.; SCARAMUZZI, R. J.; DELGADILLO, J. A. 2008. The ovulation rate in anoestrous female goats managed under grazing conditions and exposed to the male effect is increased by nutritional supplementation. Animal Reproduction Science 105: 409-416. doi:10.1016/j.anireprosci.2007.11.008 [ Links ]
DE SANTIAGO-MIRAMONTES, M. A.; MALPAUX, B.; DELGADILLO, J. A. 2009. Body condition is associated with a shorter breeding season and reduced ovulation rate in subtropical goats. Animal Reproduction Science 114: 175-182. 2008: doi:10.1016/j.anireprosci.2008.09.001 [ Links ]
DELGADILLO, J. A.; CANEDO, G. A.; CHEMINEAU, P.; GUILLAUME, D.; MALPAUX, B. 1999. Evidence for an annual reproductive rhythm independent of food availability in male Creole goats in subtropical northern Mexico. Theriogenology 52: 727-737. doi:10.1016/S0093-691-X(99)00166-1 [ Links ]
DELGADILLO J. A. 2010. Environmental and social cues can be used in combination to develop sustainable breeding techniques for goat reproduction in the subtropics. Animal. doi:10.1017/S175173111000140. [ Links ]
DELGADILLO, J. A.; VÉLEZ, L. I. 2010: Stimulation of reproductive activity in anovulatory French Alpine goats exposed to bucks treated only with artificially long days. Animal. doi:10.1017/S1751731110001345. [ Links ]
FITZ-RODRÍGUEZ, G.; DE SANTIAGO-MIRAMONTES, M. A.; SCARAMUZZI, R. J.; MALPAUX, B.; DELGADILLO, J. A. 2009. Nutritional supplementation improves ovulation and pregnancy rates in female goats managed under natural grazing conditions and exposed to the male effect. Animal Reproduction Science 116: 85-94. doi:10.1016/j. anireprosci.2009.01.004 [ Links ]
FLORES-NAJERA M. J.; MEZA-HERRERA, C. A.; ECHAVA-RRIA, F. G.; VILLAGOMEZ, E.; IÑIGUEZ, L.; SALINAS, H.; GONZÁLEZ-BULNES, A. 2010. Influence of nutritional and socio-sexual cues upon reproductive efficiency of goats exposed to the male effect under extensive conditions. Animal Production Science 50(9): 897-901. dx.doi.org/10.1071/AN10030 [ Links ]
GONZÁLEZ-BULNES, A.; MEZA-HERRERA, C. A.; REKIK,M,; BEN SALEM, H.; KRIDLI, R. T. 2011. Limiting factors and strategies for improving reproductive outputs of small ruminants reared in semi-arid environments. In: Semi-arid environments: Agriculture, water supply and vegetation. Ed: K.M. Degenovine. Nova Science Publishers Inc. Hauppauge, NY, USA, 41-60. [ Links ]
GUERRA-GARCÍA, M.; MEZA-HERRERA, C. A.; SÁNCHEZ-TORRES-ESQUEDA, M. T.; GALLEGOS-SÁNCHEZ, J.; TORRES-HERNÁNDEZ, G; PRO-MARTÍNEZ, A. 2009. IGF-1 and ovarian activity of goats in divergent body condition and supplemented with non-degradable ruminai protein. Agrociencia 43(3): 241-247. [ Links ]
LEBOEUF, B.; DELGADILLO, J. A.; MANFREDI, E.; PIACÈRE, A.; CLÉMENT, V.; MARTIN, P.; PELLICER, M.; BOUÉ, P.; DE CREMOUX, R. 2008. Management of goat reproduction and insemination for genetic improvement in France. Reproduction Domestic Animal 43(2): 379-85. doi: 10.1111/j.1439-0531.2008.01188.x [ Links ]
MARTIN, G. B.; KADOKAWA, H. 2006. "Clean, green and ethical" animal production. Case study: Reproductive efficiency in small ruminants. Journal Reproduction and Development 52: 145-152. doi:10.1262/jrd.17086-2 [ Links ]
MARTÍNEZ, G.; URRUTIA, J.; MARTÍNEZ, L.; MEZA-HERRERA, C. A. 2005. Reproductive performance of Merino Rambouillet prepubertal ewes under highland sub-tropical conditions. II. Male stimulation of seasonal anestrous prepubertal ewes. Journal of Applied Animal Research 27(1):25-28. [ Links ]
MEZA-HERRERA, C. A.; SÁNCHEZ, J. M.; CHÁVEZ-PER-CHES, J. G.; SALINAS, H.; MELLADO, M. 2004. Protein supplementation, body condition and ovarian activity in goats. Preovulatory serum profile of insulin. South Africa Journal Animal Science 34: 223-226. [ Links ]
MEZA-HERRERA, C. A.; ROSS, T.; HAWKINS, D.; HALLFORD, D. 2006. Interactions between metabolic status, pre-breeding protein supplementation, uterine pH, and embryonic mortality in ewes: Preliminary observations. Tropical Animal Health and Production 38(5): 407-413. doi: 10.1007/s11250-006-4331-6 [ Links ]
MEZA-HERRERA, C. A.; ROSS, T.; HALLFORD, D. M.; HAWKINS, D.; GONZÁLEZ-BULNES, A. 2007. Effects of body condition and protein supplementation on LH secretion and luteal function in sheep. Reproduction Domestic Animals 42(5): 461-465. doi: 10.1111/j.1439-0531.2006.00807.x [ Links ]
MEZA-HERRERA, C. A; HALLFORD, D. M.; ORTIZ, J. A.; CUEVAS, R. A.; SÁNCHEZ, J. M.; SALINAS, H; MELLADO, M.; GONZÁLEZ-BULNES, A. 2008. Body condition and protein supplementation positively affect perio-vulatory ovarian activity by non-LH mediated pathways in goats. Animal Reproduction Science 106: 412-420. doi:10.1016/j.anireprosci.2007.06.004 [ Links ]
MEZA-HERRERA, C. A.; ROSS, T.; HALLFORD, D.; HAWKINS, D.; GONZÁLEZ-BULNES, A. 2010. High periconceptional protein intake modifies uterine and embryonic relationships increasing early pregnancy losses and embryo growth retardation in sheep. Reproduction in Domestic Animals 45: 723-728. doi: 10.1111/j.1439-0531-.2009.01341.x [ Links ]
RIVAS-MUÑOZ, R.; CARRILLO, E; RODRÍGUEZ-MARTÍNEZ, R; LEYVA, C.; MELLADO, M.; VÉLIZ, F. G. 2010. Effect of body condition score of does and use of bucks subjected to added artificial light on estrus response of Alpine goats. Tropical Animal Health Production 42: 1285-1289. doi: 10.1007/s11250-010-9563-9 [ Links ]
ROSA, H. J. D.; JUNIPER, D. T.; BRYANT, M. J. 2000. The effect of exposure to oestrous ewes on rams' sexual behaviour, plasma testosterone concentration and ability to stimulate ovulation in seasonally anoestrous ewes. Applied Animal Behavior Science 67: 293-305. doi:10.1016/S0168-1591(00)00086-1 [ Links ]
ROSALES-NIETO, C. A.; GAMEZ-VAZQUEZ, H. G.; GUDI-NO-REYES, J.; REYES-RAMIREZ, E. A.; EATON, M.; STANKO, R. L.; MEZA-HERRERA, C. A.; GONZÁLEZ-BULNES, A. 2011. Nutritional and metabolic modulation of the male effect upon resumption of ovulatory activity of goats. Animal Production Science 51(2): 115-122. dx.doi. org/10.1071/AN10124 [ Links ]
SHELTON, M., 1978. Reproduction and breeding of goats. Journal of Dairy Science 61: 994-1010. doi:10.3168/jds. S0022-0302(78)83680-7 [ Links ]
URRUTIA-MORALES, J.; VILLALPANDO, F.; MEZA-HERRERA, C. A.; MANCILLA, C.; RAMIREZ. B. 2000. Effect of ram exposure on out-of-season reproductive performance of young maiden Merino Rambouillet ewes. Wool Technology & Sheep Breeding 48(2): 94-101. [ Links ]
URRUTIA-MORALES, J.; MEZA-HERRERA, C. A.; ESCOBAR-MEDINA, F. J.; GAMEZ-VAZQUEZ, H. G.; RAMIREZ-ANDRADE, B. M.; DIAZ-GOMEZ, M. O.; GONZÁLEZ-BULNES, A. 2009. Relative roles of photoperiodic and nutritional cues in modulating ovarian activity in goats. Reproductive Biology 9(3): 283-294. [ Links ]
VÉLIZ, F. G.; MORENO, S.; DUARTE, G.; VIELMA, J.; CHEMINEAU, P.; POINDRON, P.; MALPAUX, B.; DELGADILLO, J. A. 2002. Male effect in seasonally anovulatory lactating goats depends on the presence of sexually active bucks, but not estrous females. Animal Reproduction Science 72: 197-207. doi:10.1016/S0378-4320(02)00093-3 [ Links ]
VÉLIZ, F. G.; POINDRON, P.; MALPAUX, B.; DELGADILLO, J. A. 2006. Positive correlation between the liveweigh of anestrous goats and their response to the male effect with sexually active bucks. Reproduction. Nutrition and Development 6: 1-6. doi:10.1051/rnd:2006039 [ Links ]
VÉLIZ, F. G.; MEZA-HERRERA, C. A.; DE SANTIAGO-MI-RAMONTES, M. A.; ARELLANO-RODRIGUEZ, G., LEYVA, C.; RIVAS-MUÑOZ, R.; MELLADO, M. 2009. Effect of parity and progesterone priming on induction of reproductive function in Saanen goats by buck exposure. Livestock Science 125: 261-265. doi:10.1016/j. livsci.2009.05.008 [ Links ]
WALKDEN-BROWN, S. W.; RESTALL, B.; HENNIAWATI. 1993. The male effect in the Australian cashmere goat 3. Enhancement with buck nutrition and use of estrous females. Animal Reproduction Science 32: 69-84. doi:10.1016/0378-4320(93)90058-Y [ Links ]
WALKDEN-BROWN, S. W.; RESTALL, B. J.; NORTON, B. W.; SCARAMUZZI R. J. 1994: The "female effect" in Australian cashmere goats: effect of season and quality of diet on the LH and testosterone response of bucks to estrous does. Journal Reproduction Fertility 100: 521-531. doi:10.1530/jrf.0.1000521 [ Links ]
ZARCO, Q. L. A.; RODRÍGUEZ, E. F.; ANGULO, M. R. B.; VALENCIA, M. J. 1995. Female to female stimulation of ovarian activity in the ewe. Animal Reproduction Science 39: 251-258. doi:10.1016/0378-4320(95)01384-C [ Links ]

