










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.17 no.3 Chapingo sep./dic. 2011
Efecto del aprovechamiento forestal sobre la variación morfológica de Juglans pyriformis Liebm
Effect of forest harvesting on morphological variation of Juglans pyriformis Liebm
Celia Cecilia Acosta-Hernández1; Mauricio Luna-Rodríguez2; Pablo Octavio-Aguilar1; Zoylo Morales-Romero3; Jorge Galindo-González1; Juan Carlos Noa-Carrazana1; Santiago Mario Vázquez-Torres4; Lourdes Georgina Iglesias-Andreu1*
1 Instituto de Biotecnología y Ecología Aplicada (INBIOTECA), Universidad Veracruzana. Av. de las Culturas Veracruzanas Núm. 101, Col. Emiliano Zapata, C.P. 91090, Xalapa, Veracruz, MÉXICO. (*Autor para correspondencia). Correo-e: xliglesias@gmail.com
2 Laboratorio de Alta Tecnología de Xalapa, S. C. (LATEX), Universidad Veracruzana. Calle Médicos Núm. 5, Col. Unidad del Bosque, C.P. 91010, Xalapa, Veracruz, MÉXICO.
3 Facultad de Estadística e Informática. Universidad Veracruzana. Av. Xalapa Esq. Manuel Ávila Camacho s/n. C.P. 91020. Xalapa, Veracruz, MÉXICO.
4 Instituto de Investigaciones Biológicas (IIB), Universidad Veracruzana. Av. Luis Castelazo Ayala s/n, Col. Industrial Ánimas C.P. 91190, Apartado Postal 294, Xalapa, Veracruz, MÉXICO.
Recibido: 23 de abril 2011
Aceptado: 30 de agosto 2011
RESUMEN
El aprovechamiento forestal basado en la extracción de individuos fenotípicamente superiores, al igual que otros factores, podría hacer degenerar el bosque al dejar árboles de menor calidad y disminuir la diversidad genética. La población de Juglans pyriformis en San José Buenavista, Veracruz (SJB), ha sido reducida por actividades agropecuarias y aprovechamiento forestal con fines maderables bajo el método silvícola de Selección Individual, mientras que la población de Coacoatzintla, Veracruz (COA), se ha reducido por cambios de uso de suelo y explotación clandestina. El objetivo fue determinar los niveles de variación fenotípica de J. pyriformis en SJB y contrastarla con la variación encontrada en la población de COA en un estudio previo, utilizando la misma metodología. En SJB el análisis de componentes principales identificó tres indicadores, FUSTE, COPA y ALT-DIAM, con un coeficiente de variación alto 26, 30 y 42 % respectivamente. El análisis de conglomerados mostró dos grupos de árboles, y el estadístico de Levene fue significativo (P<0.05) para el indicador COPA (0.030) y ALT-DIAM (0.161). Los resultados muestran una menor variación morfológica en la población de San José Buenavista en contraste con la población de Coacoatzintla.
Palabras clave: Juglans pyriformis, manejo forestal, método silvícola de selección, indicadores morfológicos.
ABSTRACT
The forest harvesting based on the extraction of individuals phenotypically better, as well as other factors, could degrade the forest by leaving trees of lesser quality and reducing genetic diversity. The population of Juglans pyriformis in San José Buenavista, Veracruz (SJB) has been reduced by farming and logging for timber production purposes under the Silvicultural Individual Selection Method. The population of Coacoatzintla, Veracruz (COA) has been reduced by change of land use and illegal logging. The objective was to determine the levels of phenotypic variation of J. pyriformis in SJB population and contrast to COA populations found in a previously study, using the same method. In SJB principal component analysis identified three main indicators, SHAFT, CUP and HIGH-DIAM with a high coefficient of variation (26%, 30% and 42% respectively). Cluster analysis showed two groups of trees and the Levene statistic was significant (p <0.05) for the indicator CUP (0030) and HIGH-DIAM (0161). The results show less morphological variation in the population of San José Buenavista in contrast to the population of Coacoatzintla.
Key words: Juglans pyriformis, forest management, silvicultural selection method, morphological indicators.
INTRODUCCIÓN
Juglans pyriformis Liebm. (Juglandaceae) es una especie endémica de México característica del bosque mesófilo de montaña, apreciada por la calidad de su madera, que es utilizada en la elaboración de muebles Anos e instrumentos musicales. Por ello ha sido sobreexplotada y hoy se encuentra en la categoría de amenazada (SEMARNAT, 2010, NOM-059-SEMARNAT-2010). De acuerdo a Hammer y Khoshbakht (2005), esta especie podría pasar a la categoría de "En Peligro".
En el centro del estado de Veracruz, las poblaciones de Juglans pyriformis Liebm., localizadas en San José Buenavista (SJB) y Coacoatzintla (COA), se encuentran fragmentadas y reducidas por diversos factores. Estas poblaciones están separadas por barreras geográficas (barrancas y cerros), y en ambas se recolectan semillas para su comercialización sin un método de selección (Bandala1, 2008).
La población de SJB se ha reducido por actividades agropecuarias y aprovechamiento forestal con fines maderables. Esta población estuvo bajo manejo forestal por tres años (2000-2003) mediante el Método Mexicano de Ordenación de Bosques Irregulares (MMOBI), con cortas selectivas individuales como tratamiento principal para el aprovechamiento de Juglans pyriformis (Cedro-nogal). Se removieron 80 árboles, en su mayoría sanos de interés forestal con diámetro igual o mayor a 45 cm, así como dañados, huecos, secos y sobremaduros con diámetros entre 10 y 60 cm y alturas entre 10 y 15 m, que en conjunto representaron 69,637 m3 rollo total árbol (Servicio Forestal Oriente, 2000). La reducción de la población de COA se debe principalmente al crecimiento urbano, actividades agropecuarias y explotación clandestina. En esta población la extracción de individuos de J. pyriformis ha sido menor y sin un plan de manejo (Acosta-Hernández et al., 2011).
El MMOBI fue diseñado para bosques de pino y se basa en cortas selectivas individuales o por grupos del arbolado más grande y viejo (Torres, 2000). Este método recibió duras críticas, ya que al no aplicarse correctamente, sólo se extrajo el arbolado con las mejores características fenotípicas, ocasionando la degeneración del bosque al dejar árboles de menor calidad y disminuir la diversidad genética (Jardel-Pelaez, 1985; Putz, 1994; Finkeldey y Ziehe, 2004). No obstante, en muchos programas de aprovechamiento forestal de especies comerciales se sigue implementando este sistema silvícola; tal es el caso de J. pyriformis en San José Buenavista.
Por lo anterior se propuso determinar los niveles de variación fenotípica en población de J. pyriformis bajo manejo en la localidad de SJB, y contrastarla con la variación encontrada en un estudio previo para COA, sin manejo (Acosta-Hernández et al., 2011).
MATERIALES Y MÉTODOS
Se estudiaron dos poblaciones de J. pyriformis. La primera localizada en San José Buenavista (SJB), municipio de Altotonga, Veracruz (19° 48' N, 97° 03' O; altitud 1600 m). El área de estudio comprendió una superficie de 32.36 ha, compuesta por áreas remanentes de bosque mesófilo de montaña y áreas de potrero (Figura 1a). En esta área el suelo es pobre en materia orgánica y contenido de nitrógeno, de textura franco arcillo arenosa (Cra) y pH de fuertemente a moderadamente ácido (SEMARNAT 2000, NOM-021-RECNAT-2000). El clima de la región es semicálido húmedo con precipitación media anual de 1,438 mm y temperatura media anual de 14.3 °C (Servicio Meteorológico Nacional. Normales Climatológicas, 1971-2000a).
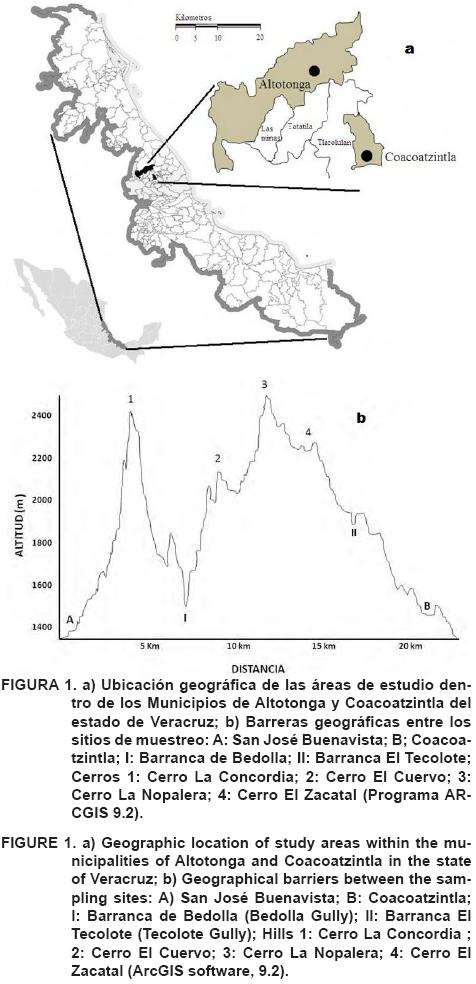
La segunda población se ubica en Coacoatzintla (COA), municipio de Coacoatzintla, Veracruz (19° 39' N, 96° 59' O; altitud 1,487m). El área de estudio comprendió una superficie aproximada de 73 ha, compuesta por áreas remanentes de bosque mesófilo, áreas de potrero y traspatio. El suelo es de rico a muy alto en materia orgánica y nitrógeno, textura es franco limosa (Cl) y pH moderadamente ácido (SEMARNAT 2000, NOM-021-RECNAT-2000). El clima en la región es templado-húmedo regular con precipitación media anual de 1,462 mm y temperatura media anual de 17.4 °C (Servicio Meteorológico Nacional. Normales Climatológicas, 1971-2000b). La distancia geográfica en línea recta entre las poblaciones de COA y SJB es de aproximadamente 20 km. Las barreras geográficas entre ambas localidades son barrancas y cerros (Figura 1b).
La variación morfológica de J. pyriformis en SJB se evaluó con la metodología reportada para COA (Acosta-Hernández et al., 2011). Se establecieron 46 sitios de 400 m2 de circunferencia. La distancia entre sitios fue igual o mayor a 50 m o hasta localizar el siguiente individuo para evitar el efecto de coancestría. En cada sitio se evaluaron de uno a cinco individuos de J. pyriformis (148 en total) mediante seis variables morfológicas utilizadas por Samaniego-Rojas (2002): forma de fuste (FF), forma de copa (FC), diámetro de copa (DC), altura de bifurcación (AB), dominancia del eje principal (DEP) y ángulo de inserción de las ramas (AIR); además de dos variables dasométricas: diámetro a la altura de 1.3 m (DN), medido con cinta métrica y altura total del árbol (ALT), estimado con clinómetro (Sunnto® 15/20).
Análisis estadísticos. Los datos morfológicos y dasométricos fueron sometidos a un Análisis de Componentes Principales (ACP). La pertinencia del ACP se determinó mediante un análisis muestral de Kaiser-Meyer-Olkin y una prueba de esfericidad de Bartlett aplicadas a la matriz de correlación resultante (Álvarez, 1994). Para cada indicador (morfológico y dasométrico) se calcularon los coeficientes de variación con base en sus desviaciones típicas. La variación morfológica intrapoblacional e interpoblacional se determinó mediante la prueba de Levene (Correa et al., 2006) aplicada a los indicadores obtenidos. La clasificación de los árboles se realizó con un Análisis de Conglomerado Jerárquico, utilizando como medida de disimilitud la distancia cuadrática euclidiana y como algoritmo de ligamiento el método de Ward con una distancia de 4 como umbral (Hidalgo, 2003). Para determinar la confiabilidad de la clasificación se utilizó un Análisis Factorial Discriminante. Los análisis fueron realizados con el programa SPSS v. 15 (SPSS, 2006). La media y desviación estándar de los indicadores por conglomerado se calculó con el programa STATISTICA v. 7.0 (Statsoft, 2004).
RESULTADOS Y DISCUSIÓN
En la población de SJB, la matriz de correlaciones pareadas reveló varias correlaciones significativas (Kaiser-Meyer-Olkin = 0.59, P<0.05; Bartlett p<0.001), lo que indica la pertinencia del análisis de componentes principales para las variables evaluadas. Se encontraron varias correlaciones positivas: FF vs FC (r = 0.201), FF vs DC (r = 0.209), AB vs DEP (r = 0.499), AIR vs FC (r = 0.366), AIR vs DC (r = 0.488); y FC vs DC (r = 0.362). Además de tres correlaciones negativas AB vs AIR (r = -0.164), AB vs DC (r = -0.289) y DEP vs AIR (r = -0.173).
Las correlaciones positivas sugieren que los árboles con fuste recto no bifurcado, presentan dominancia del eje principal con ramas insertas en un ángulo de 30° a 60° con copas circulares y vigorosas con diámetros mayores a 10 m. Las correlaciones negativas sugieren que los árboles bifurcados en el tercio inferior, tienen dominancia completa del eje principal sobre las ramas laterales, ramas insertas en un ángulo de 0° a 30° y copas pequeñas menores a 5 m de diámetro. Por otro lado, las variables dasométricas mostraron una correlación positiva alta (r = 0.836), lo cual confirma que a mayor altura mayor diámetro, situación que también sucede en Coacoatzintla (Acosta-Hernández et al., 2011).
Los resultados del ACP mostraron que los dos primeros componentes explicaron el 58 % de la varianza total. Las variables con mayor contribución en la formación del primer componente (34.6 %), y que presentaron correlaciones positivas, fueron AIR, FC y DC; estas variables califican a la copa, por lo que se les denominó como el indicador "COPA" (Cuadros 1 y 2). El segundo componente explicó el 23.1 % de la varianza total; las variables con mayor peso fueron AB y DEP. La variable FF (0.37) tuvo un valor de contribución menor a FC (0.50) y no presentó una correlación significativa con ninguna de las dos variables anteriores; no obstante, dado que permitió calificar al fuste, se agrupó con las variables AB y DEP en el indicador que se denominó "FUSTE" (Cuadros 1 y 2). Las variables dasométricas, altura (H) y diámetro a la altura de 1.3m (DN), se agruparon en el componente 1 con una varianza total de 91.8 %, por lo que se les denominó como indicador "ALT-DIAM" (Cuadros 1 y 2).
El coeficiente de variación para los tres indicadores obtenidos en SJB fue mayor al 25 % (ALT-DIAM = 42 %, COPA = 30 %, FUSTE = 26 %). En cambio en COA, sólo dos indicadores (ALT-DIAM = 50 % y COPA = 26 %) fueron los que contribuyeron a la variación dentro de la población (Acosta-Hernández et al., 2011). De acuerdo a Iglesias-Andreu y Tivo (2006), un coeficiente de variación mayor de veinte por ciento para el diámetro a la altura del pecho evidencia una alta heterogeneidad morfológica en la población.
El análisis de conglomerado efectuado en la población de SJB, clasificó a los individuos evaluados en dos grupos (Figura 2a). La población de COA reveló una mayor variación al formarse tres grupos con características consistentemente distintas (Figura 2b); en esta población el grupo III concentró a los árboles con las mejores características silvícolas, alturas >26 m, diámetros de 38-69 cm, y fustes rectos en la mayoría de los árboles, que correspondieron a 21 % de la población (Acosta-Hernández et al., 2011).
El agrupamiento en SJB fue determinado por la altura total, diámetro del tronco, forma de la copa y diámetro de la copa. El grupo I estuvo conformado por 54 árboles que correspondieron a 37.2 % de la población; los individuos de este grupo se caracterizaron por tener un fuste recto, no bifurcado con dominancia completa del eje principal, ángulo de inserción de ramas de 30° a 60°, copa de forma circular a circular irregular vigorosa, con un diámetro mayor a 10 m presentando las mejores características en cuanto a altura (18-37 m) y diámetro (2174 cm). El grupo II estuvo conformado por 94 árboles, que correspondieron al 64.2 % de la población; de éstos, trece presentaron diámetros entre 2 y 10 cm y alturas entre 3.5 y 10 m, mientras que el resto (81 árboles) presentaron diámetros entre 10 y 24 cm, alturas entre 10 y 18.9 m, fuste ligeramente torcido con dominancia completa del eje inicial, pocos individuos bifurcados, ángulo de inserción de las ramas de 30° a 60°, diámetro de copa entre 5 y 10 m, de forma circular a menos de un círculo.
El análisis discriminante indica un 99 % de confiabilidad para la clasificación obtenida por el Análisis de Conglomerado Jerárquico para SJB. El valor de la Lambda de Wilks (0.308) fue significativo (P<0.05), lo cual demuestra que hubo una buena clasificación de los individuos dentro de los dos grupos formados. Asimismo, los estadísticos descriptivos para los indicadores ALT-DIAM y COPA mostraron diferencias entre conglomerados. Los valores detectados para el FUSTE resultaron similares para los dos conglomerados (Cuadro 3).
De acuerdo a la prueba de Levene, la variación intrapoblacional en SJB fue significativa (P<0.05) para los indicadores COPA (0.030) y ALT-DIAM (0.161) (Cuadro 4). En COA la prueba de Levene fue significativa (P<0.05) para estos mismos indicadores (COPA = 0.036; ALT-DIAM =0.024), no así para FUSTE (0.534). El indicador FUSTE no mostró diferencias significativas (P<0.05) en ninguna de las dos poblaciones, lo que muestra que los indicadores COPA y ALT-DIAM son los más importantes en la clasificación de los individuos para ambas poblaciones (Acosta-Hernández et al., 2011).
Respecto a la variación interpoblacional (SJB vs COA), la prueba de Levene fue significativa (p<0.05) para los tres indicadores, COPA, FUSTE y ALT-DIAM (Cuadro 4), lo cual revela que las poblaciones son morfológicamente diferentes. El promedio de estos indicadores es mayor en COA con respecto a SJB (Cuadro 3).
En el presente estudio la altura, el diámetro y la copa fueron los caracteres que aportaron mayor variación y determinaron el agrupamiento de los árboles dentro y entre poblaciones. En ambas, las características edáficas y climáticas son diferentes, por lo que es posible que la variación intra e interpoblacional observada se deba a la respuesta de cada individuo a las condiciones del sitio. Esta variación ambiental afecta en mayor medida a la altura y el diámetro (Zobel y Talbert, 1988). En dos regiones de Cataluña, España, se encontró que la variación en altura y diámetro de J. regia está significativamente ligada a las características edáficas y climáticas de las localidades de origen (Aletá et al., 2003).
La variación morfológica detectada en SJB (148 árboles clasificados en dos grupos) en relación con la observada en COA (112 árboles clasificados en tres grupos), puede explicarse si se tiene en cuenta la extracción selectiva que se llevó a cabo en la población de SJB, durante el periodo 2000-2003. Conforme a la evaluación dasométrica, en SJB se encontraron las siguientes categorías diamétricas: 1) 25 % entre 1.5 y 17cm; 2) 31.8 % entre 18 y 32 cm; 3) 35.8 % entre 33 y 47cm; 4) 7.4 % mayores a 48 cm. En la categoría diamétrica 3 no se encontraron árboles de 45 cm de DAP y solo dos presentaron DAP mayores a 45 cm y en la categoría 4 sólo se agruparon 11 árboles. Estos datos demuestran que la extracción selectiva; fue dirigida a árboles sanos de interés forestal con diámetro normal igual o mayor a 45 cm. En la población de COA, los árboles se distribuyeron en las categorías diamétricas de la siguiente manera: 1) 24 %; 2) 27 %; 3) 29 % y 4) 20 %. En esta población la eliminación del arbolado no ha sido selectiva, más bien ha estado determinada por el crecimiento urbano y explotación clandestina (Acosta-Hernández et al., 2011). Lo que explica la proporción más homogénea de los individuos en cada categoría diamétrica.
Considerando que no se evaluó a la población de SJB previamente al proceso de extracción selectiva, no podemos asegurar que antes del aprovechamiento la población tuviese igual o mayor variación que la de COA. Sin embargo, teniendo en cuenta que la única diferencia evidente en la historia de las poblaciones es la extracción selectiva, asumimos que tanto la variación intrapoblacional en SJB como la variación interpoblacional (SJB vs COA) podrían estar dadas por el aprovechamiento forestal llevado a cabo en SJB.
Asimismo, es posible que debido a la eliminación selectiva de ejemplares en SJB, algunas características de importancia silvícola se estén perdiendo, y con ello algunos alelos relacionados con estos fenotipos (Climent et al., 2008), lo que se presume también puede estar sucediendo en COA por la reducción de la población, aunque por causas diferentes.
Diversos autores (Putz, 1994; Lee et al., 2002; Wickneswari et al., 2004) indican que las prácticas de aprovechamiento forestal basadas únicamente en el diámetro, pueden provocar la erosión genética y la declinación del bosque. Según estos autores, el aprovechamiento afecta la composición y diversidad vegetal, la edad, la distribución de la densidad y genera un alto riesgo de deriva genética en las poblaciones raras o reducidas.
Por otro lado, la variación observada entre las poblaciones no necesariamente está ligada a la disminución del potencial evolutivo (variabilidad genética) de J. pyriformis en SJB (Lande, 1999), sino que podría tener un componente ambiental fenotípico (Sultan, 2003; Gianoli, 2004; Chambel et al., 2005). Para corroborar esta información, actualmente se realiza un estudio genético para ambas poblaciones.
CONCLUSIONES
Los indicadores de COPA y ALT-DIAM, fueron los que determinaron la variación intrapoblacional y la clasificación de los individuos en las dos poblaciones estudiadas. La población de J. pyriformis de SJB presenta menor variación morfológica que la de COA. Debido a que el programa de aprovechamiento forestal sólo fue aplicado en SJB y que no existe otra diferencia en la historia de las poblaciones, podemos asumir que estas diferencias en la variación morfológica se deben a este factor.
AGRADECIMIENTOS
El primer autor agradece el apoyo del CONACYT, por la beca otorgada para la realización de los estudios de doctorado en Ecología y Biotecnología del INBIOTECA-UV. Especialmente agradecemos el invaluable apoyo en campo de J. Javier Juárez De La Fuente, Jesús Baillet E., Santiago Ángeles T. y al Sr. Carmelo Rivera Hernández.
LITERATURA CITADA
Acosta-Hernández, C. C., Luna-Rodríguez, M., Noa-Carrazana, J. C., Galindo-González, J., Vázquez-Torres, S. M., Morales-Romero, Z., & Iglesias-Andreu, L. G. (2011). Caracterización morfológica y dasométrica de la especie amenazada Juglans pyriformis Liebm. Revista Chapingo Serie Ciencias Forestales y del Ambiente, 17, 59-67. [ Links ]
Aletà, N., Ninot, A., & Voltas, J. (2003). Caracterización del comportamiento agroforestal de doce genotipos del nogal (Juglans sp) en dos localidades de Cataluña. Investigación Agraria. Sistema de Recursos Forestales, 12, 39-50. [ Links ]
Álvarez, C. R. (1994). Estadística multivariante y no paramétrica con SPSS: aplicación a las ciencias de la salud. Madrid, España. Ediciones Díaz Santos, S.A. [ Links ]
Correa, J. C., Iral, R., & Rojas, L. (2006). Estudio de potencia de pruebas de homogeneidad de varianza. Revista Colombiana de Estadística, 29, 57-76. [ Links ]
Climent, J., Chambel, R., Brabas, D., Voltas, J., & Alía, R. (2008). Evaluación genética de la población forestal: Concepto y resultados disponibles para rodales de pinos españoles. Boletín del CIDEU, 6-7,69:82. [ Links ]
Chambel, M. R., Climent, J., Alía, R., & Valladares, F. (2005). Phenotypic plasticity: a useful framework for understanding adaptation in forest species. Investigación Agraria, Sistemas y Recursos Forestales, 14, 334-344. [ Links ]
Finkeldey, R. & Ziehe, M. (2004). Genetic implications of silvicultural regimes. Forest Ecology and Management, 197, 231-244. [ Links ]
Gianoli, E. (2004). Plasticidad fenotípica adaptativa en plantas. pp. 13-25. In: Fisiología Ecológica en plantas. Mecanismos y Respuestas a Estrés en los ecosistemas. Valparaíso Chile. Marino C. H. [ Links ]
Hammer, K. & Khoshbakht, K. (2005). Towards a "red list" for crop plant species. Genetic Resources and Crop Evolution, 52, 249-265. [ Links ]
Hidalgo, R. (2003). Variabilidad genética y caracterización de especies vegetales. pp. 2-49. In: Análisis Estadístico de Datos de Caracterización Morfológica de Recursos Fitogenéticos (2003). Boletín Técnico 8, Tito L. F. y Rigoberto Hidalgo (eds). Cali, Colombia. Instituto Internacional de Recursos Fitogenéticos (IPGRI). [ Links ]
Iglesias-Andreu, L. G. & Tivo, F. Y. (2006). Caracterización morfométrica de las poblaciones de Pinus hartwegii Lindl del Cofre de Perote, Veracruz, México. Ra Ximhai, 2, 449-468. [ Links ]
Jardel-Pelaez, E. J. (1985). Una revisión crítica del método mexicano de ordenación de bosques, desde el punto de vista de la ecología de poblaciones. Ciencia Forestal, 58, 3-16. [ Links ]
Lande, R. (1999). Extinction risks from anthropogenic, ecological and genetic factors. pp. 1-22. In: Genetics and the extinction of species. Landweber, L. F. and Dobson, A. P. (eds). Princeton, NJ: Princeton University Press. [ Links ]
Lee, C. T., Wickneswari, R., Mahani, M. C., & Zakri, Y. A. H. (2002). Effect of selective logging on the genetic diversity of Scaphium macropodum. Biology Conservation, 104,107-118. [ Links ]
Putz, F. E. (1994). Approaches to sustainable forest management. Bogor, Indonesia, CIFOR. [ Links ]
Samaniego-Rojas, C. (2002). Las fuentes semilleras y semillas forestales nativas en el austro ecuatoriano: participación social en el manejo. Loja, Ecuador. Fundación Ecológica Condesur, [ Links ].
SEMARNAT. (2000). Norma Oficial Mexicana NOM-021-RECNAT-2000. México, D.F. Diario Oficial. [ Links ]
SEMARNAT. (2010). Norma Oficial Mexicana NOM-059-SEMARNAT-2010, Protección ambiental-Especies nativas de México de flora y fauna Silvestres-Categorías de riesgo y especificaciones para su inclusión, exclusión o cambio-Lista de especies en riesgo. México, D.F. Diario Oficial. [ Links ]
Servicio Forestal Oriente. (2000). Programa Forestal de aprovechamiento maderable persistente del predio rústico innominado de la Congregación de San José Buenavista, Municipio de Altotonga, Veracruz. Veracruz: Servicio Forestal Oriente. [ Links ]
Servicio Meteorológico Nacional. Normales Climatológicas 1971-2000a. Estado de Veracruz. Estación Meteorológica 00030008, Altotonga, Veracruz. CONAGUA. http://smn.conagua.gob.mx/climatologia/normales/estacion/catalogos/cat_ver.html [ Links ]
Servicio Meteorológico Nacional. Normales Climatológicas 1971-2000b. Estado de Veracruz. Estación Meteorológica 00030114, Naolinco de Victoria, Veracruz. CONAGUA. http://smn.conagua.gob.mx/climatologia/normales/estacion/catalogos/cat_ver.html [ Links ]
SPSS, Inc. (2006). SPSS: Statistical Package for the Social Sciences, versión 15 para Windows. [ Links ]
STATSOFT, Inc. (2004). STATISTICA (data analysis software system), version 7. [ Links ]
Sultan, E. S. (2003). Phenotypic plasticity in plants: a case study in ecological development. Evolution and Development, 5, 25-33. [ Links ]
Torres, R. J. M. (2000). Sostenibilidad del volumen de cosecha calculado con el Método Mexicano de Ordenación de Montes. Instituto de Ecología A. C. Madera y Bosques, 6, 57-72. [ Links ]
Wickneswari, R., Ho, W. S., Lee, K. S., & Lee, Y. C. T. (2004). Impact of disturbance on population and genetic structure of tropical forest trees. Forest Genetics, 11,193-201. [ Links ]
Zobel, J. B. & Talbert, J. T. (1988). Técnicas de Mejoramiento Genético en Árboles Forestales. Trad. por M. Guzmán Ortiz. LIMUSA. México, D.F. 545p. [ Links ]
1 Bandala Santiago. 2008. Proveedor de semilla forestal a viveros oficiales y particulares. San José Buenavista, municipio de Altotonga s/n.

