










Serviços Personalizados
Journal
Artigo
 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Acessos
Acessos
Links relacionados
-
 Similares em
SciELO
Similares em
SciELO
Compartilhar

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versão On-line ISSN 2007-4018versão impressa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.17 no.2 Chapingo Ago. 2011
https://doi.org/10.5154/r.rchscfa.2010.06.039
Estructura espacial de Pachycereus weberi (Coult.) Backeb. En la cañada del Zopilote, Guerrero
Spatial structure of Pachycereus weberi (Coult.) Backeb. In cañada del Zopilote, Guerrero
Blanca Estela Linzaga–Román1; Gregorio Ángeles–Pérez*1; Cesario Catalán–Heverástico2; Patricia Hernández de la Rosa1
1 Posgrado Forestal. Colegio de Postgraduados. Montecillo, Estado de México. C. P. 56230. MÉXICO. Correo–e: gangeles@colpos.mx (*Autor para correspondencia)
2 Unidad Académica de Ciencias Agropecuarias y Ambientales, Universidad Autónoma de Guerrero. Periférico Poniente s/n, Col. Guadalupe. Iguala, Guerrero. C. P. 40030. MÉXICO.
Recibido: 15 de junio, 2010
Aceptado: 23 de agosto, 2010
RESUMEN
El propósito del presente estudio fue evaluar la estructura poblacional de Pachycereus weberi en la Cañada del Zopilote, Guerrero, México. Se estableció una parcela de observación de 50 x 50 m (2,500 m2), dividida en cuadros de 10 x 10 m para facilitar el levantamiento de la información de campo. Todos los individuos de P. weberi con altura ≥ 0.10 m se marcaron, se les midió la altura total y Diámetro Normal (DN), y se ubicaron por sus coordenadas X y Y. Además, todos los individuos de las especies leñosas asociadas con DN ≥ 5 cm, se marcaron, se identificaron y se les registró su diámetro, y coordenadas X y Y. Se determinó el índice de distribución y correlación espacial de individuos de P. weberi, divididos en dos estadios de desarrollo, juveniles y adultos, a través del índice de Ripley. Se registraron un total de 18 especies, asociadas a P. weberi. El mayor índice de Valor de Importancia Relativa lo registró P. weberi con 41.1 %, seguido de Acacia tenciifolia con 22.6 %. Se encontraron individuos de P. weberi con más de 8.5 m de altura y diámetros desde 5 a 43.8 cm. El patrón de distribución espacial fue de forma agregada tanto para individuos en periodo de desarrollo juvenil como adulto. La correlación espacial de ambos estadios de desarrollo fue de repulsión en distancias de 2 a 13 m. La distribución diamétrica encontrada sugiere un reclutamiento de nuevos individuos a la población de tipo episódico. Es decir, existen pocas incorporaciones en tamaños intermedios entre juveniles y adultos.
Palabras clave: Pachycereus weberi, Índice de Ripley, índice de valor de importancia relativa, patrón de distribución espacial, patrón de correlación espacial.
ABSTRACT
The aim of this study was to determine the population structure of Pachycereus weberi in Cañada del Zopilote (Buzzard Canyon), Guerrero, Mexico. An observation plot of 50 x 50 m (2,500 rrP) was established and divided into quadrats of 10 x 10 m to facilitate the field survey. All P. weberi individuals with height ≥ 0.10 m were tagged, and their total height, diameter at breast height (dbh), and location by X and Y coordinates were recorded. In addition, all individuals of associated woody species with dbh ≥ 5 cm were tagged, identified and their diameter and coordinates within the plot recorded. We calculated the spatial distribution and correlation patterns of the P. weberi population divided into two developmental stages, juveniles and adults, using Ripley's index. A total of 18 species associated with P. weberi were recorded. P. weberiattained the highest Index of Relative Importance Value with 41.1 %, followed by Acacia tenciifolia with 22.6 %. P. weberi individuals with total height greater than 8.5 m and diameters from 5 to 43.8 cm were recorded. The spatial distribution pattern was clumped for individuals in both juvenile and adult development stages. The spatial correlation showed a repulsion pattern at distances of 2 to 13 m between juvenile and adult P. weberi. The diameter distribution suggests an episodic recruitment behavior, with low recruitment rates in intermediate diameter classes between juveniles and adults.
Key words: Pachycereus weberi, Ripley's index, Index of Relative Importance Value, spatial distribution pattern, spatial correlation pattern.
INTRODUCCIÓN
La estructura de una población, es el resultado de la acción de fuerzas bióticas y abióticas a que los miembros de dicha población, y de sus ancestros, han estado sujetas en el pasado (Hutchings, 1997). Por consecuencia la estructura de los bosques, es producto de la dinámica de los ecosistemas y se ve afectada por una variedad de factores del sitio y de las especies que ahí coexisten. Por lo tanto, el estudio de la estructura proporciona información relevante respecto a la composición de las comunidades, y poblaciones dentro de las comunidades (Álvarez et al., 2004). Asimismo, la estructura de una población se puede ver como el arreglo tridimensional de los individuos, así como la proporción de tamaños y su arreglo espacial (horizontal y vertical) (Godínez–Álvarez et al., 2008).
Se han reconocido tres patrones generales de distribución de los individuos en el espacio, distribución aleatoria, distribución uniforme y distribución agregada (Nanami et al., 1999). Una distribución aleatoria, indica que la probabilidad de encontrar a un individuo es la misma para todos los puntos del espacio, o que todos los individuos tienen la misma probabilidad de ser hallados en cada punto del espacio. Una distribución uniforme significa que las distancias entre individuos son aproximadamente las mismas dentro de la población. Una distribución agregada, implica que los individuos se agrupan en parches, dejando porciones del espacio relativamente desocupadas (Pielou, 1977). Este tipo de análisis representan la base para conocer el estado que guardan las poblaciones dentro de comunidades y que puede revelar los mecanismos a que han estado expuestos sus componentes.
Por tanto, con el presente trabajo sobre Pachycereus weberi se analizó la estructura, distribución espacial, y la composición de las especies de las comunidades donde se presenta en la Cañada del Zopilote, en el estado de Guerrero. Asimismo, se analizaron los patrones de distribución y correlación espacial entre estadios de desarrollo juvenil y adulto de la especie. La meta, es generar información que sea útil para sugerir algunas alternativas de uso y conservación de la especie, con el propósito de reducir el riesgo de sobre–explotación, ya que la madera de estos árboles es utilizada por los artesanos y es una fuente de ingresos económicos para los habitantes de la región. Los objetivos del presente estudio fueron: 1) Determinar el patrón de distribución y correlación espacial de dos estadios de desarrollo de Pachycereus weberi, 2) Determinar la composición de especies de la comunidad y 3) Analizar la estructura poblacional de Pachycereus weberi en una parcela de observación localizada en su distribución natural en la Cañada del Zopilote, Guerrero.
MATERIALES Y MÉTODOS
Área de estudio
Se ubica en la Cañada del Zopilote, en el estado de Guerrero, la cual se localiza en la cuenca del Río Balsas a lo largo de la carretera federal 95, México–Acapulco; entre las coordenadas geográficas aproximadas de 17° 39' 41" y 18° 03' 59" de latitud norte, y 99° 31' 01" y 99° 46' 35" de longitud oeste. La Cañada del Zopilote cubre una superficie aproximada de 738 km2. El tipo de vegetación predominante en la región es el bosque tropical caducifolio (sensu Rzedowski, 2006). Presenta una alta diversidad de especies del género Bursera spp. y es una comunidad muy rica en endemismos (Arriaga et al., 2000).
Trabajo de campo
Se realizó un recorrido de campo para localizar los sitios donde se distribuye Pachycereus weberi en la Cañada del Zopilote. Con base en la presencia y abundancia de individuos de P. weberi, se estableció una parcela de observación de 50 x 50 m (2,500 m2), en el paraje denominado La Cañada. Esta parcela se dividió en cuadros de 10 x 10 m para facilitar la toma de datos de campo (Mueller–Dombois y Ellenberg, 1974). En la delimitación de la parcela se consideró la inclinación del terreno para compensar la distancia horizontal por la pendiente.
Todos los individuos de Pachycereus weberi dentro de la parcela se marcaron y se diferenciaron en dos estadios de desarrollo: juveniles (individuos con 0.1 < altura < 4m altura) y adultos (individuos con altura > 4 m). La diferenciación entre juveniles y adultos se realizó con base en la observación de estructuras reproductivas, las cuales ocurren en individuos con altura mínima de 4 m. Se registraron los datos de altura total de la especie desde el nivel del suelo hasta el ápice de cada árbol; se registró el diámetro normal (1.30 m sobre el nivel del suelo), y el número de ramas por árbol. Los individuos se ubicaron por sus coordenadas X y Y dentro de la parcela de observación.
Análisis de la información
Para analizar la estructura de la comunidad, se determinó el Índice de Valor de Importancia Relativa (IVIR) a través de la densidad relativa, dominancia relativa y frecuencia relativa (Curtis y Mclntosh, 1951). Asimismo se determinó la composición de especies y se construyeron histogramas de frecuencias con base en el DN.
Distribución espacial
Con los datos de las coordenadas X, Y, de los individuos, se realizó el análisis de distribución espacial de la especie por estadio de desarrollo. El patrón espacial univariado de individuos de P. weberi se analizó a través de la función K (t) de Ripley (Ripley, 1977) que está dada por:
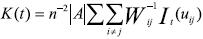
Donde: n es el número de individuos en la parcela A; |A| es el área de la parcela; uij.. es la distancia entre i–ésimo y j–ésimo individuo en A; lt(u) = 1 si u ≤ t y 0 si no lo es; mientras que Wij es la proporción de la circunferencia de un circulo con centro en el /–ésimo individuo y radio uij que está dentro de A; y la sumatoria es de todos los pares de individuos no mayor que t (Ripley, 1977; Diggle, 1983). Se utilizó la siguiente transformación de la raíz cuadrada de K (t) sugerida por Besag (1977).
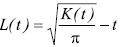
Donde: K(t) es la función de Ripley; π = 3.1416; y t es la distancia.
La significancia estadística de la función L (t) se determinó con simulaciones de Monte Carlo (Besag, 1977; Besag y Diggle, 1977; Marriott, 1979). Los límites máximos y mínimos de L(t) se definieron con un 99 % de confiabilidad.
Correlación espacial
Se analizó la correlación entre los estadios de desarrollo de Pachycereus weberi y la de las especies asociadas con diámetro normal ≥ 5 cm y con una abundancia mayor a 10 individuos. La interacción espacial entre los árboles adultos (con floración) y juveniles (sin floración). Se analizó usando la función bivariada L12(t,), misma que es una transformación de K12(t) de Ripley (Lotwick y Silverman, 1982):
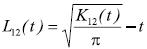
La significancia estadística de la función K12(t) se determinó con simulaciones de Monte Carlo (Besag, 1977; Besag y Diggle, 1977; Marriott, 1979). Los límites máximos y mínimos de L12(t) se definieron con un 99 % de confiabilidad.
RESULTADOS Y DISCUSIÓN
Densidad
Se registró una densidad total de 1,680 individuos·ha–1 de los cuales P. weberi tuvo el 39.3 % (660 individuos·ha1) y Acacia tenciifolia el 34.5 % (580 individuos·ha1) (Cuadro 1). Asimismo, se registró un área basal total de 17.5 m2·ha–1, y nuevamente P. weberi tuvo la mayor dominancia relativa con 63.5 %. Pachycereus weberi presentó el mayor IVIR con 41.1 %, seguida de Acacia tenciifolia con 22.6 %. Esta especie está estrechamente relacionada con Pachycereus weberi en este sitio de estudio y dominan fisonómicamente el paisaje donde co–ocurren. La tercera especie con el mayor IVIR fue Euphorbia schlechtendalii con 6.77 %, valor muy inferior que el registrado por las dos especies anteriores (Cuadro 1).
Distribución de tamaños
La distribución diamétrica de Pachycereus weberi se presenta en la Figura 1. La mayor concentración de individuos ocurrió en la categoría diamétrica de 0.1 a 5 cm. Una segunda moda se detectó en individuos de 10 a 20 cm de diámetro. El diámetro máximo registrado para esta especie fue de 48.9 cm. La forma de la distribución diamétrica sugiere la existencia de reclutamiento de nuevos individuos a la población, que puede ser de tipo episódica, es decir, no constante.
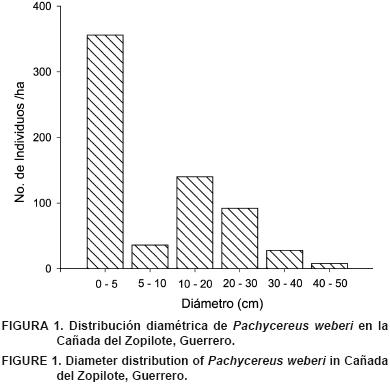
Los individuos de Pachycereus weberi son plantas arborescentes y ampliamente ramificadas, y es una de las especies de Cactaceae que alcanzan mayor altura en Norteamérica (Backeberg, 1960; Bravo, 1978; Britton y Rose, 1920). Arias y Terrazas (2009) mencionan que algunos ejemplares pueden alcanzar hasta 60 cm de diámetro y 15 m de altura. Estas características hacen que la especie sea conspicua en las comunidades donde se presenta.
A pesar de que en la parcela se registró un mayor número de individuos en la categoría de juveniles, no quiere decir que el reclutamiento de nuevos individuos a la población se esté presentando de manera constante. Lo anterior es más evidente si se analiza la distribución de alturas (Figura 2). Se observó un número casi nulo de individuos en categoría intermedias entre juveniles y adultos reproductivos. El tamaño mínimo promedio de individuos en que se observó floración fue de 6.0 m de altura. Este hallazgo indica posibles problemas para la regeneración de la especie, debido a la alta mortalidad en las primeras etapas de desarrollo (Silvertown y Charlesworth, 2001); sin embargo, este proceso debe ser evaluado a profundidad para determinar las causales de este patrón en la estructura poblacional.

Patrón de distribución espacial
La distribución espacial y el índice de distribución espacial de Ripley se presentan en la Figura 3 para juveniles y adultos de P. weberi. Se puede observar que tanto los juveniles como los adultos presentan un patrón agregado en el terreno prácticamente en todas las distancias analizadas (0–25 m), aunque el grado de intensidad de agregación es ligeramente mayor en los juveniles (Figura 3). La mayoría de las especies de Cactaceae muestran una distribución espacial agregada, lo cual es un reflejo de la alta variación en la distribución de los recursos ambientales y del sitio a lo largo de su distribución (Valiente–Banuet y Ezcurra, 1991; Larrea–Alcázar y Soriano, 2008; Zenteno–Ruiz et al., 2009).
Por otra parte, la agrupación de los individuos puede deberse a muchos factores, entre estos se pueden encontrar las condiciones topográficas específicas. En la Cañada del Zopilote Pachycereus weberi crece en sitios con pendientes pronunciadas, afloramientos rocosos y en cañadas. Sin embargo, este es un aspecto que aún se necesita explorar a mayor detalle. Las especies incluidas en Pachycereus muestran, en términos generales, preferencias por substratos calizos, aunque no en forma exclusiva y se asocian con diferentes comunidades vegetales, principalmente los bosques tropicales caducifolios, los bosques espinosos y los matorrales xerófilos (Backeberg, 1960; Bravo, 1978; Rzedoski, 1978).
La agrupación espacial de los árboles de Pachycereus weberi, parece ocurrir en toda el área de la Cañada, esto se puede observar a gran distancia, por la altura, y cobertura que tienen estos cactus, que los hace distinguir entre toda la vegetación presente en el área. Un patrón agregado indica la presencia de interacciones entre los individuos, o entre los individuos y el medio (Leirana–Alcocer y Parra–Tabla, 1999). Existen muchas causas probables para la formación de un patrón agregado. Pueden ser por factores intrínsecos o extrínsecos, estas dos clases de factores pueden igualmente interactuar de muchas formas, y afectar la trayectoria evolutiva de la población o especie a todos los niveles de organización (Larrea–Alcázar y Soriano, 2008).
Patrón de correlación espacial
El Índice de correlación espacial de Ripley entre juveniles y adultos de P.weberi mostró un patrón de repulsión entre distancias de 2 a 13 m; mientras que se presentó un patrón independiente a distancias mayores de 13 m. No es común encontrar estudios donde se analice la correlación espacial entre estadios de desarrollo en poblaciones de cactáceas. Sin embargo, esta información es muy importante porque puede definir la relación espacial entre estadios de desarrollo y los posibles sitios donde ambos pueden prosperar. Se ha establecido que en poblaciones de especies longevas, como en especies arbóreas, las condiciones que son favorables para el establecimiento y crecimiento de individuos en etapa de plántula, pueden no ser las más favorables para estadios juveniles y adultos (Grubb, 1977). Por tanto, los sitios de colonización de los juveniles pueden no ser los sitios donde un individuo tiene la mayor probabilidad de alcanzar su estado adulto.
La relación de repulsión entre juveniles y adultos podría ser explicada de acuerdo a lo reportado sobre la alta mortalidad de semillas y juveniles cerca de los árboles progenitores. Las enfermedades específicas y la depredación se concentran cerca de árboles progenitores causando mortalidad denso–dependiente, ya que existe mayor probabilidad de supervivencia a medida que aumenta la distancia entre la descendencia y los adultos productores de semillas (Janzen, 1970; Augspurger y Kelly, 1984; Clark y Clark, 1984; Silvertown, 1987; Condit etal., 1994; Silvertown y Charlesworth, 2001). Sin embargo, es necesario realizar futuras evaluaciones de la demografía de juveniles para determinar los sitios específicos donde un individuo tiene mayor oportunidad de supervivencia.
Se realizó un intento por determinar si especies arbóreas asociadas a Pachycereus.weberi , presentaron algún patrón de asociación con juveniles y adultos. En la Figura 4 se presenta la correlación espacial entre juveniles y adultos de P. weberiy Acacia tenciifolia. Se pudo observar un patrón de atracción entre juveniles y la especie de Acac a en distancias menores a 3 m. Lo anterior indica que los juveniles de P. weberise distribuyen bajo el dosel de Acac a.tenc fol a. Sin embargo, la correlación espacial fue independiente en distancias mayores a 3 m, por lo que la relación espacial entre adultos de P. weberiy elementos de Acac a.tenc fol a fue de independencia espacial.
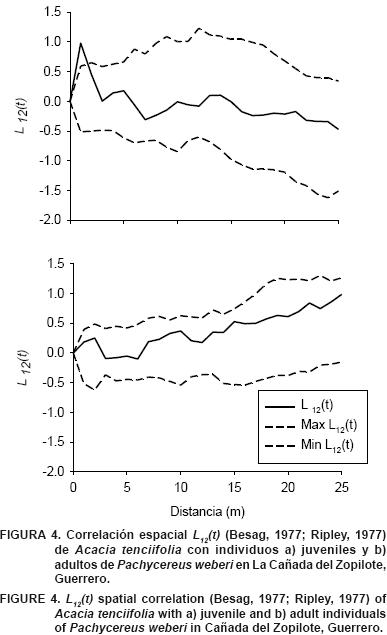
En especies de cactáceas está ampliamente documentada la relación con especies arbustivas y arbóreas que funcionan como facilitadoras a través del síndrome de nodricismo (Reyes–Olivas etal., 2002; López etal., 2007; Larrea–Alcázar etal., 2008; Larrea–Alcázar y Soriano, 2008; Zenteno–Ruiz etal., 2009). El hecho de que se presente una correlación espacial positiva a distancias cortas (0–3 m) entre juveniles de P..weberi e individuos de Acac a.tenc fol a, puede ser un indicio de la existencia de dicho síndrome. Sin embargo, la falta de contundencia de la relación de nodricismo entre Acacia tenciifolia y P. %, respectivamente, lo que demuestra su importancia estructural y fisonómica en las comunidades donde se presenta. Asimismo, el patrón de distribución espacial tanto de individuos en estadio juvenil como adulto, es agrupado, lo que sugiere sitios específicos donde la especie es capaz de establecerse y alcanzar su estatura adulta. La correlación espacial intraespecífica fue de repulsión, mientras que la interespecífica con Acacia tenciifolia fue de atracción con juveniles en distancias de 0 a 3 m.
LITERATURA CITADA
ÁLVAREZ, R.; GODÍNEZ–ÁLVAREZ, H.; GUZMÁN, U.; DÁVILA, P. 2004. Aspectos ecológicos de dos cactáceas mexicanas amenazadas: implicaciones para su conservación. Boletín de la Sociedad Botánica de México 75: 7–16. [ Links ]
ARIAS, S.; TERRAZAS, T. 2009. Taxonomic revision of Pachycereus (Cactaceae). Systematic Botany 34(1): 68–83. [ Links ]
ARRIAGA, L.; ESPINOZA, J. M.; AGUILAR, C.; MARTÍNEZ E.; GÓMEZ L.; LOA, E. (Coords.). 2000. Regiones terrestres prioritarias de México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. [ Links ]
AUGSPURGER, C. K.; KELLY, C. K. 1984. Pathogen mortality of tropical tree seedling: experimental studies of the effects of dispersal distance, seedling density, and light conditions. Oecologia 61: 211–217. [ Links ]
BACKEBERG, C. 1960. Die Cactaceae, Vol. 3. Gustav Fisher–Verlag, Jena, Germany. [ Links ]
BESAG, J. 1977. Contribution to the discussion on Dr. Ripley's paper. Journal of the Royal Statistical Society B (39): 193–195. [ Links ]
BESAG, J.; DIGGLE, P. J. 1977. Simple Monte Carlo tests for spatial pattern. Applied Statistics. 26: 327–333. [ Links ]
BRAVO. H. 1978. Las cactáceas de México. Vol. l. Universidad Nacional Autónoma de México. México, D. F. México. 743 p. [ Links ]
BRITTON, N. L.; ROSE, J. N. 1920. The Cactaceae. Vol, ll, 2a ed. Dover Publications, Inc. New York. USA. 241 p. [ Links ]
CLARK, D. A.; CLARK, D .B. 1984. Spacing dynamics of a tropical rain forest tree: evaluation of the Janzen–Connell model. The American Naturalist 124 (6): 769–788. [ Links ]
CONDIT, R.; HUBBELL, S. P.; FOSTER, R .B. 1994. Density–dependence in two understory tree species in a Neotropical forest. Ecology 75: 671–680. [ Links ]
CURTIS, J. T.; McINTOSH, R. P. 1951. An upland forest continuum in the pariré–forest border region of Wisconsin. Ecology 32:476–496. [ Links ]
DIGGLE, P. J. 1983. Statistical analysis of spatial point patterns. Academic Press, London, UK. 148 p. [ Links ]
GODÍNEZ–ÁLVAREZ, H. T.; VALVERDE, T.; ORTEGA–BAES, P. 2008. Demographic trends in the Cactaceae. Botanical Review 69(2): 173–203. [ Links ]
GRUBB, P. J. 1977. The maintenance of species–richness in plant communities: the importance of the regeneration Niche. Biological Review 52:107–145. [ Links ]
HUTCHINGS, M. J. 1997. The Structure of plant populations. En: Crawley MJ (ed) Plant Ecology: Blackwell Science. Oxford. UK. pp. 325–358. [ Links ]
JANZEN, D. H. 1970. Herbivores and the number of tree species in tropical forests. American Naturalist 104 (940):201–528. [ Links ]
LARREA–ALCAZAR, D. M.; MURILLO, J. J.; FIGUEREDO, C. J.; SORIANO, P. J. 2008. Spatial associations between two globose cacti and two dominant mimosoid bushes in a tropical semiarid enclave. Ecotropicos 21(2): 97–105. [ Links ]
LARREA–ALCÁZAR, D. M.; SORIANO, P. J. 2008. Columnar cacti–shrub relationships in an Andean semiarid valley in western Venezuela. Plant Ecology 196: 153–161. [ Links ]
LEIRANA–ALCOCER, J.; PARRA–TABLA, P. 1999. Factor affecting the distribution, abundance and seedling survival of Mammillaria gaumer, and endemic cactus of coastal Yucatán, México. Journal of Arid Environments 41: 421–428. [ Links ]
LÓPEZ, R.; VALDIVIA, S.; SANJINÉS, N.; QUINTANA, de la, D. 2007. The role of nurse plants in the establishment of shrub seedlings in the semi–arid subtropical Andes. Oecologia 52: 779–790. [ Links ]
LOTWICK, H. W.; SILVERMAN, B. W. 1982. Methods for analyzing spatial processes of several types of points. Journal of the Royal Statistical Society, London, Series B 44: 403–413. [ Links ]
MARRIOTT, F. H. C. 1979. Barnard's Monte Carlo tests: how many simulations? Applied Statistics 28: 75–77. [ Links ]
MUELLER–DOMBOIS, D; ELLENBERG, H. 1974. Aims and methods of vegetation ecology. John Willey and Sons. New York. USA. 547 p. [ Links ]
NANAMI, S.; KAWAGUCHI, H.; YAMAKURA, T. 1999. Dioecy–induced spatial patterns of two codominant tree species, Podocarpus nagi and Neolitsea aciculata. Journal of Ecology 87: 678–687. [ Links ]
PIELOU, E. C. 1977. Mathematical ecology. John Wiley and Sons. New York. USA. 385 p. [ Links ]
REYES–OLIVAS, A.; GARCÍA–MOYA, E.; LÓPEZ–MATA, L. 2002. Cacti–shrub interactions in the coastal desert of northern Sinaloa, Mexico. Journal of Arid Environments (2002) 52: 431–445. [ Links ]
RIPLEY, B. D. 1977. Modelling spatial patterns (with discussion). Journal of the Royal Statistical Society B39: 172–212. [ Links ]
RZEDOWSKI, J. 1978. Vegetación de Mexico. Edit. Limusa. México, D.F. México. 432 p. [ Links ]
SILVERTON, J. 1987. Introduction to plant Population Ecology. Second edition. Longman Scientific and Technical. 229 p. [ Links ]
SILVERTOWN, J.; CHARLESWORTH, D. 2001. Introduction to plant population biology. Fourth edition. Blackwell Science. London, UK. 347 p. [ Links ]
VALIENTE–BANUET, A.; EZCURRA, E. 1991. Shade as a cause of association between the cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisana in the Tehuacán Valley, México. Journal of Ecology 79: 961–971. [ Links ]
ZENTENO–RUIZ, F. R.; LÓPEZ, R. P.; LARREA–ALCÁZAR, D. M. 2009. Patrones de distribución espacial de Parodia maassii (Heese) A. Berger (Cactaceae) en un semidesierto de los Andes subtropicales, la prepuna. Ecología en Bolivia 44(2): 17–26. [ Links ]

