










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.17 no.1 Chapingo ene./abr. 2011
https://doi.org/10.5154/r.rchscfa.2010.03.015
Caracterización morfológica y dasométrica de la especie amenazada Juglans pyriformis Liebm.
Morphological and dasometric characterization of Juglans pyriformis Liebm ., a threatened species
Celia Cecilia Acosta–Hernández1; Mauricio Luna–Rodríguez2; Juan Carlos Noa–Carrazana1; Jorge Galindo–González1; Santiago Mario Vázquez–Torres3; Zoylo Morales–Romero4; Lourdes Georgina Iglesias–Andreu1
1 Instituto de Biotecnología y Ecología Aplicada (INBIOTECA), Universidad Veracruzana. Av. de las Culturas Veracruzanas Núm.101, Col. E. Zapata, C. P. 91090, Xalapa, Veracruz, MÉXICO.(xliglesias@gmail.com)
2 Laboratorio de Alta Tecnología de Xalapa, S. C. (LATEX), Universidad Veracruzana. Calle Médicos Núm. 5, Col. Unidad del Bosque, C. P. 91010, Xalapa, Veracruz, MÉXICO.
3 Centro de Investigaciones Biológicas (CIB), Universidad Veracruzana. Av. Luis Castelazo Ayala s/n, Col. Industrial Ánimas C. P. 91190, Apartado Postal 294, Xalapa, Veracruz, MÉXICO.
4 Facultad de Estadística e Informática. Universidad Veracruzana. MÉXICO.
Recibido: 29 de marzo, 2010
Aceptado: 21 de septiembre, 2010
RESUMEN
Juglans pyriformis es una especie endémica del bosque mesófilo de montaña, que se encuentra amenazada. Con el objetivo de determinar la variación morfológica dentro de la población de J. pyriformis de Coacoatzintla, Veracruz. Se evaluaron seis variables morfológicas y dos dasométricas, mediante un muestreo no probabilístico, en 1 a 5 individuos por sitio, ubicados a una distancia igual o mayor a 50 m o hasta localizar el siguiente individuo. Los datos fueron analizados mediante pruebas de independencia entre las variables, Análisis de Componentes Principales, análisis de conglomerado, coeficiente de variación y prueba de homogeneidad de varianza de Levene. Los resultados obtenidos permitieron identificar dos indicadores para las variables morfológicas COPA y FUSTE, y uno para las dasométricas ALT–DIAM. El indicador ALT–DIAM aportó mayor variación dentro de la población (42 %). Se encontraron diferencias significativas dentro de la población para los indicadores de COPA y ALT–DIAM.
Palabras clave: Variación morfológica, indicadores morfológicos, población natural vegetal, bosque mesófilo de montaña, componentes principales.
ABSTRACT
Juglans pyriformis is a threatened species endemic to a cloud forest environment. In order to determine the morphological variation within the J. pyriformis population in Coacoatzintla, Veracruz. Six morphological and two dasometric variables were evaluated, through non–probability sampling, in 1 to 5 individuals per site, located at a distance equal to or greater than 50 m or until locating the next individual. The data were analyzed using tests of independence between variables, Principal Components Analysis, cluster analysis, coefficient of variation and Levene's test of homogeneity of variance. The results obtained allowed identifying two indicators for the CROWN and TRUNK morphological variables, and one for the HEIGHT–DIAMETER dasometric relationship. The HEIGHT–DIAMETER indicator provided more variation within the population (42 %). Significant differences were found within the population for the CROWN and HEIGHT–DIAMETER indicators.
Key words: Morphological variation, morphological indicators, natural plant population, cloud forest, principal components analysis.
INTRODUCCIÓN
El bosque mesófilo de montaña mexicano tiene la más alta diversidad de especies y endemismos (30 %). Su cobertura original era del 1 % del territorio nacional (Rzedowski, 1978), aunque actualmente se estima la pérdida de más del 50 % de estos bosques a causa de la deforestación y fragmentación, quedando aproximadamente unas 800,000 ha en condiciones relativamente conservadas (Toledo et al., 1989; Challenger, 1998). A nivel global, el bosque mesófilo de montaña es considerado uno de los ambientes más amenazados (Hamilton et al., 1995). En la parte central de Veracruz los fragmentos remanentes de este bosque se encuentran aislados por potreros, cultivos y asentamientos humanos, con la consecuente disminución de la biodiversidad y servicios ambientales que proporcionan (Williams–Linera et al., 2002), lo que coloca a sus especies endémicas en riesgo de desaparecer.
Juglans pyriformis Liebm. (Juglandaceae), conocida comúnmente como cedro–nogal, es una especie característica del bosque mesófilo, endémica y amenazada (SEMARNAT, 2002). Sus poblaciones se distribuyen de forma discontinua y fragmentada a través de Hidalgo, Oaxaca y Veracruz, entre los 1,200 y 1,600 msnm. Las poblaciones se encuentran aisladas y reducidas como consecuencia de las actividades antropogénicas, incluyendo la sobreexplotación. Esto ocasiona que en algunas localidades de difícil acceso se observen sólo individuos aislados (Rzedowski, 1978; Narave, 1983; Luna–Vega et al., 2006).
Los trabajos realizados con J. pyriformis (Maning, 1957; Rzedowski, 1978; Narave 1983; Niembro et al., 2004; Benítez et al., 2004; Luna–Vega et al., 2006; Martínez et al., 2009) abordan aspectos generales de su taxonomía, distribución, fenología, producción, biogeografía y el uso potencial de su semilla como biocombustible. En Europa, las especies J. regia y J. nigra han sido ampliamente estudiadas, tanto por su importancia económica como por el valor de su madera y frutos (Balci et al., 2001; Aleta et al., 2003; Fady et al., 2003; Colaric et al., 2006; Pollegioni et al., 2006; Oguz et al., 2008).
En México, el valor ecológico y la importancia económica de J. pyriformis apenas se reconoce. Particularmente en el estado de Veracruz, en el periodo 2002–2008, se establecieron aproximadamente 400 ha de plantaciones de J. pyriformis en el marco de los programas PRODEPLAN y PROÁRBOL (CONAFOR, 2008). Los viveros encargados de producir las plántulas para dichas plantaciones, fueron abastecidos con semilla proveniente de colectas masales realizadas por particulares en poblaciones naturales de fácil acceso, sin conocimiento previo de la variabilidad de las mismas, ni clasificación de los árboles de colecta (Bandala1, 2008). Una de ellas es la población de J. pyriformis, localizada en Coacoatzintla, municipio de Coacoatzintla, Veracruz, cuyo estado actual se desconoce, no obstante su notable reducción por actividades antropogénicas.
Por lo anterior, nos hemos propuesto determinar los niveles de variación y la estructura de las poblaciones de J. pyriformis mediante un estudio morfológico y molecular de las mismas. El presente trabajo evalúa la variación morfológica dentro de la población natural de J. pyriformis, de la localidad de Coacoatzintla, Veracruz.
MATERIALES Y MÉTODOS
La localidad de Coacoatzintla se localiza en el centro del estado de Veracruz (19° 39' N, 96° 59' O), a una altitud de 1,487 m (Figura 1). El clima es templado–húmedo regular con lluvias abundantes en verano y principios de otoño; la temperatura media anual es de 17.4 °C (máx. 25.3 °C, mín. 8.3 °C), con una precipitación media anual de 1,437.7 mm (Servicio Meteorológico Nacional).

Se seleccionaron todos los árboles que crecen en los remanentes del bosque, áreas de potrero y áreas de traspatio aledañas al bosque. Se cubrió una superficie aproximada de 73 ha, estableciéndose 30 sitios (Figura 1). Se seleccionaron 112 individuos por un método no probabilístico incidental o casual. Para ello se realizó un recorrido previo y se establecieron circunferencias de 400 m2 donde se localizaron los individuos. La selección se basó en dos criterios: que cada sitio estuviese representado por 1 a 5 individuos y que la distancia entre sitios fuese igual o mayor a 50 m o hasta localizar el siguiente individuo, para evitar el efecto de coancestría.
Por cada individuo se registraron seis variables morfológicas, siguiendo los criterios de Samaniego–Rojas (2002) en la identificación y selección de fuentes semilleras que garanticen la colecta de germoplasma de calidad para el establecimiento de plantaciones comerciales; por tanto, los mayores puntajes están asignados a las mejores características (Cuadro 1). Asimismo, se midió el diámetro a la altura del pecho (DAP) con cinta métrica y altura total del árbol (ALT) con clinómetro (marca Sunnto 15/20). Estas dos variables fueron estimadas posteriormente, aplicando las fórmulas correspondientes.
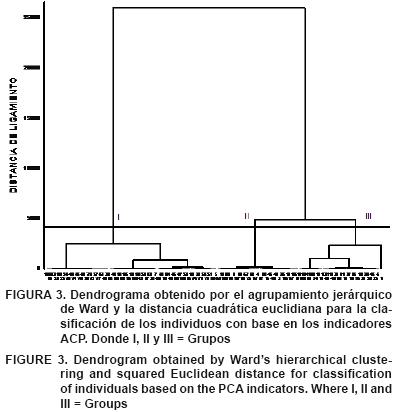
Análisis estadísticos. Los datos obtenidos fueron incluidos en un Análisis de Componentes Principales (ACP). Éste se realizó de forma independiente a las variables morfológicas y dasométricas, a fin de reducir su dimensionalidad y transformarlas a un conjunto menor de variables independientes denominadas indicadores (Hidalgo, 2003). La pertinencia del ACP se determinó mediante las pruebas de adecuación muestral de Kaiser–Meyer–Olkin (KMO) y de esfericidad de Bartlett, aplicadas a la matriz de correlación resultante. A partir de las desviaciones típicas de los indicadores, se obtuvo el Coeficiente de Variación (CV), a fin de describir el comportamiento de la población. Con base en los indicadores se determinó la variación morfológica intrapoblacional mediante la prueba de homogeneidad de varianza de Levene. La clasificación de los individuos se realizó mediante un Análisis de Conglomerado (AC), empleando como medida de disimilitud la distancia cuadrática euclidiana y el método de Ward como algoritmo de ligamiento y distancia de 4 como umbral (Hidalgo, 2003). La confiabilidad de la clasificación se comprobó aplicando un análisis discriminante. Los análisis estadísticos fueron realizados con los programas SPSS y STATISTICA (versión 7.0).
RESULTADOS Y DISCUSIÓN
La matriz de correlación pareada entre variables reveló que existen correlaciones bajas pero significativas (Kaiser–Meyer–Olkin 0.56, P<0.05; Bartlett P<0.001). Las relaciones positivas fueron: forma de copa (FC) con forma de fuste (FF) (0.233); ángulo de inserción de las ramas (AIR) con FC (0.260) y con diámetro de copa (DC) (0.239); finalmente DC con FC (0.562). Una asociación negativa resultó significativa: altura de bifurcación (AB) con DC (–0.217). Estos resultados sugieren que a formas circulares mejor definidas de la copa (circular y circular irregular) hay mayor DC (>10m), las ramas se insertan en mayor ángulo (60 a 90°) y el fuste es más recto, pero a mayor AB (1/3 superior) el DC es menor. Respecto a las variables dasométricas existe una correlación positiva (0.89). Según Husch et al., 1982 existe una correlación entre la altura de los árboles (h) y su diámetro a la altura del pecho (DAP) lo que se comprueba en ésta especie.
El ACP de las variables morfológicas mostró que los dos primeros componentes tienen valores superiores a uno, y en conjunto explican el 49.7 % de la varianza total. En cuanto a las variables dasométricas, el primer componente explica el 94.5 % de la varianza total (Cuadro 2). Las variables que presentan mayor correlación con el primer componente son: AIR, FC y DC (Cuadro 3). Estas tres variables calificaron a la copa y presentaron una correlación positiva, por lo que en conjunto se les denominó como el indicador "COPA". Las variables con mayor peso en la formación del segundo componente, que califica al fuste, fueron: AB y Dominancia del Eje Principal (DEP). Para este segundo componente, la variable FF tiene un valor menor a FC, pero dada su condición de calificar el fuste se agrupó con las dos variables anteriores (AB y DEP) para formar el indicador designado "FUSTE". Con relación a las variables dasométricas h y DAP, ambas conforman al componente 1 con un valor de correlación alto (0.972), por lo cual se agruparon en el indicador designado como "ALT–DIAM" (Cuadro 3).


El coeficiente de variación para los indicadores ALT–DIAM, COPA y FUSTE (50, 26 y 18 %, respectivamente) demuestra una amplia variación entre los individuos en altura, diámetro y copa, pero una menor variación en las características del fuste (Figura 2). La prueba de homogeneidad de varianzas de Levene (Cuadro 4) resultó significativa (P<0.05) para los indicadores de COPA (0.036) y ALT–DIAM (0.024). Estos resultados coinciden con los obtenidos en el CV, corroborando que los caracteres evaluados para copa, altura y diámetro son los que determinan principalmente las diferencias dentro de la población de J. pyriformis.


El AC diferenció 3 grupos (Figura 3). El grupo I fue el más heterogéneo y agrupó 50 individuos que representaron el 45 % de la población. Los árboles de este grupo presentaron alturas extremas entre 4 y 25 m, y diámetros menores a 30 cm; asimismo, presentaron fustes ligeramente torcidos y bifurcados en los tres tercios, con dominancia completa y parcial del eje principal sobre las ramas laterales, ángulo de inserción de ramas de 30° a 90°, con un solo individuo de 0° a 30°, diámetro de copa de <5 a >10 m, forma de copa menor a medio círculo y árboles con pocas ramas. El grupo II, conformado por 38 individuos, agrupó al 34 % de los árboles evaluados. Éstos se caracterizaron por presentar alturas > 22 m y diámetros entre los 16 y 57 cm, fustes ligeramente torcidos, bifurcados a partir del tercio medio e inferior, con dominancia completa y parcial del eje principal sobre las ramas laterales, ángulo de inserción de ramas de 30° a 90°, diámetro de copa entre 5 y 10 m y forma de copa de medio círculo a menos de medio círculo. El grupo III agrupó 24 individuos que correspondieron al 21 % del total de los árboles evaluados. Todos los individuos de este grupo presentaron las mejores características, con alturas >26 m y diámetros de 38 a 69 cm, fustes rectos en la mayoría de los árboles y el resto ligeramente torcidos, bifurcados a partir de los tres tercios. El 62 % de los árboles del grupo presentan dominancia parcial del eje principal sobre las ramas laterales y el resto dominancia completa sobre las ramas laterales, ángulo de inserción de las ramas de 30° a 90°, copa vigorosa (>10 m) y forma de copa de circular irregular. El valor de Lambda de Wilks fue bajo (0.160), lo cual demuestra una buena clasificación de los individuos dentro de los grupos formados (P≤0.001).
Los valores medios de las estadísticas descriptivas del indicador ALT–DIAM, resultaron altamente divergentes para los tres grupos (Cuadro 5). Para el indicador de FUSTE se constató cierta semejanza en los valores medios de los tres grupos (Cuadro 5). En cambio, las medias del indicador de COPA de los grupos 1 y 2 respecto al valor medio del grupo 3 presentaron valores distantes (Cuadro 5). Estos datos confirman que los indicadores de ALT–DIAM y COPA fueron los que más contribuyeron a la clasificación de los individuos en la población estudiada.
La variación intrapoblacional detectada por los dos anteriores indicadores puede atribuirse a las características del sitio donde se desarrolla la especie, ya que el 21 % de los árboles con mayores alturas, diámetros y mejor conformación de copa (grupo III) crecen en las áreas abiertas (potreros y traspatio), a diferencia de los árboles del grupo I, con características dasométricas y morfológicas menores, que crecen bajo el dosel. Los árboles del grupo II, con características intermedias, crecen en los bordes de los remanentes de bosque. Esto confirma a la altura y el diámetro como caracteres fuertemente influidos por las características físicas y climáticas de los sitios en donde se desarrollan los árboles (Zobel y Talbert, 1988), y coincide con lo reportado para las poblaciones de P. hartewii del Pico de Orizaba y Cofre de Perote (Iglesias–Andreu y Tivo–Fernández, 2006). Asimismo, los trabajos del comportamiento agroforestal de 12 genotipos de J. regia (Aletà et al., 2003) y de 24 progenies de la misma especie (Aletà et al., 2009) refieren que las variaciones en altura, diámetro y conformación de los árboles se deben a las características de los sitios de plantación.
Adicionalmente, Aletà et al. (2004) determinaron la heredabilidad de los caracteres de altura, diámetro y conformación de copa (ramificación, grosor de ramas y ángulo de inserción de las ramas) en J. regia. En particular, encontraron que la variación en altura y diámetro está asociada a la heterogeneidad de la familia, tanto a nivel genético como ambiental. En J. nigra, Beineke (1983) encontró valores de heredabilidad entre 0.33 a 0.45 para la altura y 0.25 a 0.40 para el diámetro. En nuestro estudio, si consideramos que la edad de los árboles evaluados varía entre 25 y 150 años de edad y que es una especie anemófila (Narave, 1983), podemos suponer la existencia histórica de un intercambio genético entre los mismos. Por ello, es posible que la variabilidad fenotípica detectada sea el resultado de procesos de selección dirigida debido a la extracción selectiva de los árboles mejor conformados, hecho que, conjuntamente con la reducción y fragmentación que ha sufrido esta población, podría estar llevándola a un cuello de botella reciente. Por lo anterior, se recomienda llevar a cabo estudios genéticos en las poblaciones de J. pyriformis para evaluar el impacto del manejo sobre la variabilidad genética de la especie.
Por otra parte, con base en los criterios de Samaniego–Rojas (2002), los árboles de los grupos II y III pertenecen a la Clase 1 y pueden ser usados principalmente para la construcción, fabricación de muebles y artesanías. Los árboles del grupo I corresponden a las Clases 1 y 2. Los árboles de la clase 2 pueden ser fuente de germoplasma para el establecimiento de futuras plantaciones, y el diseño de estrategias para el mantenimiento de la variabilidad local y la conservación de la especie en esta zona.
CONCLUSIONES
El método utilizado para la caracterización morfológica de los árboles, es eficiente y rápido para la identificación de fuentes semilleras con buenas características, en el establecimiento de plantaciones.
La variación morfológica dentro de la población de J. pyriformis se atribuye principalmente al indicador ALT–DIAM, y en menor medida al indicador de COPA. Con base en la variación detectada, los individuos fueron clasificados en tres grupos, lo cual indica una variación importante.
El nivel de variabilidad local detectado es atribuible a las características físicas y climáticas de los sitios, no obstante lo cual ésta población puede ser considerada un recurso genético con potencial en la conservación del nivel de variabilidad local.
AGRADECIMIENTOS
El autor principal, C. C. Acosta–Hernández, agradece al CONACYT la beca otorgada para la realización de los estudios de doctorado en Ecología y Biotecnología, del INBIOTECA–UV. Especialmente agradecemos al Doctor Pablo Octavio–Aguilar por la corrección y comentarios de este manuscrito y el invaluable apoyo en campo del señor J. Javier Juárez de La Fuente, Jesús Baillet E., Santiago Ángeles T. y señor Carmelo.
LITERATURA CITADA
ALETA, N.; NINOT, A.; VOLTAS, J. 2003. Caracterización del comportamiento agroforestal de doce genotipos del nogal (Juglans sp) en dos localidades de Cataluña. Investigación Agraria. Sistema de Recursos Forestales. 12(1): 39–50. [ Links ]
ALETA, N.; NINOT, A.; VOLTAS, J. 2004. Retrospective evaluation of parental selection in nursery tests of Juglans regia L. Using a Mixed Model Analysis. Silvae Genetica 53, (1): 26–33. [ Links ]
ALETA, N.; VILANOVA, A; DÍAZ, R; VOLTAS, J. 2009. Comportamiento de 24 progenies de nogal común (Juglans regia L.) para su uso en la producción de madera. Resultados hasta el sexto año de crecimiento. 5° Congreso Forestal Español. 12p. http://www.recercat.net/bitstream/2072/43631/1/Comunicaci%C3%B3n+N.+Alet%C3%A0.pdf. [ Links ]
BALCI, I.; BALTA, F.; KAZANKAYA, A.; SEN, S. M. 2001. Promising native walnut genotypes (Juglans regia L.) of the east black sea region of Turkey. Journal American Pomological Society 55(4): 204–208. [ Links ]
BEINEKE W. F. 1983. Genetic improvement of black walnut for timber production. Plant Breeding Reviews 1: 236–266. [ Links ]
BENÍTEZ, B. G.; PULIDO–SALAS, T. P.; EQUIHUA–ZAMORA, M. 2004. Árboles multiusos nativos de Veracruz para reforestación, restauración y plantaciones. Instituto de Ecología A.C, SIGOLFO, CONAFOR. Xalapa, Veracruz, México. 288 p. [ Links ]
COLARIC, M. F.; STAMPAR, M. H.; SOLAR, A. 2006. Sensory evaluation of different walnut cultivars (Juglans regia L.). Acta Agriculturae Slovenica 87: 403–413. [ Links ]
CONAFOR. 2008. Expedientes de los proyectos autorizados por el PRO–DEPLAN (1997–2008). Gerencia Nacional de Plantaciones. PRODEPLAN. Comisión Nacional Forestal. México. [ Links ]
CHALLENGER, A. 1998. Utilización y conservación de los ecosistemas terrestres de México: pasado, presente y futuro. Comisión Nacional para el Uso y Conocimiento de la Biodiversidad, Instituto de Biología de la UNAM y Agrupación Sierra Madre, S. C., México. Instituto Nacional de Ecología. 847 p. [ Links ]
FADY, B.; DUCCI, F.; ALETA, N.; BECQUEY, J.; DÍAZ–VÁZQUEZ, R.; FERNÁNDEZ–LÓPEZ, F.; JAY–ALLEMAND, C.; LEFE'VRE, F.; NINOT, A.; PANETSOS, K.; PARIS, P.; PISANELLI, A.; RUMPF, H. 2003. Walnut demonstrates strong genetic variability for adaptive and wood quality traits in a network of juvenile field tests across Europe. Kluwer Academic Pu–blishers. New Forest 25: 211–225. [ Links ]
HAMILTON, L. S.; JUVIK J. L.; SCATENA, F. N. 1995. Tropical montane cloud forests: pp. 1–23. In: Introduction and workshop synthesis. The Puerto Rico tropical montane cloud forest symposium. L. S. HAMILTON, J. L. JUVIK y F. N. SCATENA (eds.). Serie: Ecological Studies No. 110. Springer Verlag, Nueva York. [ Links ]
HIDALGO, R. 2003. Variabilidad Genética y Caracterización de Especies Vegetales. In: Análisis Estadístico de Datos de Caracterización Morfológica de Recursos Fitogenéticos (2003). FRANCO T., L. e HIDALGO R. (eds.). Instituto Internacional de Recursos Fitogenéticos (IPGRI). Boletín Técnico. Cali, Colombia Núm. 8: 89 p. [ Links ]
HUSH, B.; MILLER, C. I.; BEERS, T. W. 1982. Forest Mensuration. John Wiley & Sons. New York. 347 p. [ Links ]
IGLESIAS, A. L.; TIVO, F. Y. 2006. Caracterización morfométrica de la población de Pinus hartwegii Lindl, del Cofre de Perote, Veracruz, México. Ra Ximhai 2(2): 449–468. [ Links ]
LUNA–VEGA, I.; ALCÁNTARA–AYALA, O.; CONTRERAS–MEDINA, R.; PONCE–VARGAS, A. 2006. Biogeography, current knowledge and conservation of threatened vascular plants characteristic of Mexican temperate forests. Biodiversity and Conservation, 15: 3773–3799. DOI 10.1007/s10531–005–5401–1. [ Links ]
MANNING, W. E. 1957. The genus Juglans in Mexico and Central America. Journal of Arnold Arboretum 38: 121–150. [ Links ]
MARTÍNEZ–PÉREZ, D.; PARTIDA–SEDAS, J. G.; PÉREZ–PORTILLA, E. 2009. Especies vegetales para biocombustibles en sistemas agrícolas diversificados en Veracruz, México. Revista Brasileira de Agroecología, (4): 2 3438–4342. [ Links ]
NARAVE F. H. 1983. Juglandaceae. Fascículo 31. INIREB. Xalapa, Veracruz. Flora de Veracruz. 30 p. [ Links ]
NIEMBRO–ROCAS, A.; MORATO, I.; CUEVAS–SÁNCHEZ, J. A. 2004. Catálogo de frutos y semillas de árboles y arbustos de valor actual y potencial para el desarrollo forestal de Veracruz y Puebla. Instituto de Ecología, A.C. Reporte Final del proyecto CONACYT–CONAFOR–2002–CO1–5741. Xalapa, Veracruz. México. 928 p. [ Links ]
OGUZ, H. I.; DOGAN, A.; GÜN, A.; KAZANKAYA, A.; MURADOGLU, F. 2008. Determination of Fruit Characteristics of Some Turkish Walnut (Juglans regia L.) Cultivars Grown in Bozdogan District (Aydin). Journal of Applied Biological Sciences 2(2): 89–91. [ Links ]
POLLEGIONI, P.; BARTOLI, S.; MALVOLTI, M. E.; MAPELLI, S.; BERTANI, A.; CANNATA, F. 2006. Identificación di ecotipi italiani di Juglans regia L. mediante marcadori molecolari, morfologici e biochimici. Forest@ 3(4): 598–60. [ Links ]
RZEDOWSKI, J. 1978. Vegetación de México. LIMUSA. México D.F. 432 p. [ Links ]
SAMANIEGO–ROJAS, C. 2002. Las fuentes semilleras y semillas forestales nativas en el austro ecuatoriano: participación social en el manejo. Fundación Ecológica Condesur, Loja, Ecuador. pp. 1–14. [ Links ]
SEMARNAT. 2002. "Norma Oficial Mexicana (NOM–059–ECOL–2001)". Diario Oficial. Instituto Nacional de Ecología. México, DF. 102 p. [ Links ]
SERVICIO METEOROLÓGICO NACIONAL. Estación Meteorológica 00030114 Naolinco de Victoria. CONAGUA. http://smn.conagua.gob.mx/climatologia/normales/estacion/ver/NORMAL30114.TXT [ Links ]
TOLEDO, V. M.; CARABIAS, J.; TOLEDO, C.; GONZÁLEZ–PACHECO, C. 1989. La producción rural en México: alternativas ecológicas. Colección Medio Ambiente, núm. 6. Fundación Universo Veintiuno, México. 392 p. [ Links ]
WILLIAMS–LINERA, G.; MANSON, R. H.; ISUNZA–VERA, E. 2002. La fragmentación del bosque mesófilo de montaña y patrones de uso del suelo en la región oeste de Xalapa, Veracruz, México. Madera y Bosques 8(1): 73–89. [ Links ]
ZOBEL, J. B.; TALBERT, J. T. 1988. Técnicas de mejoramiento genético en árboles forestales. Trad. por M. Guzmán Ortiz. LIMUSA. México, D.F. 545 p. [ Links ]
1 Bandala Santiago. 2008. Proveedor de semilla forestal a viveros oficiales y particulares. San José Buenavista, Mpio. de Altotonga s/n.

