










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.15 no.2 Chapingo jul./dic. 2009
Estimación de factores de expansión de carbono en comunidades forestales templadas del norte de Durango, México
Estimating carbon expansion factors in temperate forest communities of northern Durango, Mexico
F. M. Silva–Arredondo1; J. de J. Návar–Cháidez2*
1 Consultora Forestal Privada. Preparatoria Núm. 4. Linares, Nuevo León. MÉXICO.
2 Profesor de Manejo de Recursos Naturales. CIIDIR–IPN Unidad Durango. Calle Sigma s/n. Fraccionamiento 20 de Noviembre 2. Durango, Dgo. C. P. 34220. MÉXICO. Tel. 618–8142091. Correo–e: jnavar@ipn.mx. *Autor para correspondencia.
Recibido: 27 de marzo, 2009
Aceptado: 25 de agosto, 2009
RESUMEN
Este estudio tuvo por objetivo la cuantificación del almacén de carbono en la biomasa aérea de comunidades forestales templadas del norte de Durango, México. Se estimaron los valores de la concentración de carbono de diversos componentes de biomasa de 25 árboles provenientes de bosques de coníferas. Datos dasométricos de rodales inventariados, junto con ecuaciones alométricas y concentraciones de carbono sirvieron para estimar los factores de expansión de carbono, utilizando los volúmenes por hectárea. Se modeló la dependencia de los factores de expansión de carbono en función de las características dasométricas de los árboles y se determinó la distribución probabilística de estos factores. Los resultados mostraron que los factores de expansión de carbono son dependientes de la altura promedio del rodal y se distribuyeron normalmente. Esta fuente de información es clave en el desarrollo de planes de manejo de servicios ambientales de los ecosistemas forestales y permitirá el uso más amplio de datos de inventarios convencionales.
Palabras clave: existencias reales, factores de expansión de carbono, densidad de carbono, servicios ambientales.
ABSTRACT
This research set as objective to quantify carbon stocks in aerial biomass of temperate communities of northern Durango, Mexico. The carbon concentration of several biomass components of 25 trees were quantified in the laboratory. Dasometric data of inventoried trees, together with alometric equations to estimate biomass and carbon concentrations were employed to estimate carbon expansion factors using stand volume. The carbon expansion factors were modeled as a function of the dasometric features of trees at the stand scale and the normal probabilistic density function was fitted to obtain information on the skewness of this parameter. The results showed that the carbon expansion factors are dependent of the mean tree height and they distributed normally. This information is important for the development of environmental management plans of forest ecosystems and will allow the broader use of forest inventory data.
Keywords: standing volume, carbon expansion factors, carbon density, environmental services.
INTRODUCCIÓN
Los factores de expansión son parámetros que transforman el volumen fustal por árbol o por unidad de superficie en densidad de carbono por árbol o por la misma unidad superficie por cada tipo de vegetación (Gracia et al., 2004). Estos facilitan los cálculos de la densidad de carbono a partir de las existencias reales en un bosque. Se han desarrollado con la finalidad de someter a las comunidades forestales eventualmente al pago de servicios ambientales emergentes de las diferentes iniciativas nacionales e internacionales.
Existe preocupación por el calentamiento de la biosfera terrestre como consecuencia del aumento de la concentración de los gases de efecto invernadero de origen antropogénico, de los cuales el CO2 es el más importante producido por las actividades humanas. Mediante un adecuado manejo de los recursos forestales se podría mitigar el incremento del CO2 a la atmósfera y reducir así su efecto en el calentamiento global. El ciclo del CO2 en la tierra y en la atmósfera fue constante hasta antes de la era industrial; sin embargo, a partir del uso de los combustibles fósiles, este gas se ha incrementado considerablemente, lo que ha traído como consecuencia un aumento en la temperatura, situación que se podría agravar de no tomarse las medidas conducentes para reducir emisiones o secuestrar el CO2 atmósferico en sistemas terrestres estables. El manejo forestal sustentable podría mitigar el aumento de la emisión del CO2 a la atmósfera ya que este gas es utilizado por las plantas para realizar la fotosíntesis y por lo tanto el C es capturado, almacenado o secuestrado para mantener las estructuras leñosas de los organismos vegetales (Brown y Schroeder, 2000).
La importancia de los bosques como sumideros de carbono ha tomado mucha importancia en las últimas décadas debido a que en años anteriores se han deforestado grandes áreas de vegetación. SEMARNAT (2001) reporta una tasa de deforestación de 3.93 % para México. Para varios bosques tropicales y subtropicales de México se han reportado valores de deforestación de 4.3 % en las últimas dos décadas (Dirzo y García, 1991; Cairns et al., 1995; 2000; De Jong et al., 1999; 2000; Návar–Cháidez, 2008). De esa manera se considera que los bosques podrían ser considerados como sumideros de C, y los dueños podrían recibir un pago por la venta de bonos de C en base al acuerdo de los diferentes protocolos internacionales e iniciativas nacionales (Masera et al., 2000; Návar et al., 2005).
Para recibir el pago de los servicios ambientales por el secuestro de C, es necesario comprobar la cantidad que se incrementa al ecosistema; por lo tanto, lo primero que se debe hacer es medir la cantidad de carbono almacenado en el bosque, para lo cual se requiere que inicialmente se calcule el peso seco de la biomasa de los diferentes compartimientos. Una vez estimada la biomasa, se puede hacer una estimación generalizada de que el contenido de carbono varía alrededor de 50 % del peso seco de la biomasa (Husch et al., 2003).
Sin embargo, la precisión de la estimación del carbono para proyectos ambientales hace importante evaluar las concentraciones de carbono en componentes de biomasa, sobre todo en especies o regiones en donde aún no existe esta información. También es importante evaluar las existencias de carbono y sus variaciones espacio–temporales en comunidades forestales, tareas en la que no se dispone todavía de una gran experiencia en varias regiones del mundo. Un método simple utiliza el volumen de madera calculado en los inventarios forestales y con los denominados factores de expansión de la biomasa se estima la biomasa arbórea, y con otro factor de expansión se transforma en densidad de carbono.
En el interés de reducir costos y tiempo, así como hacer aproximaciones, normalmente se aplican factores de expansión desarrollados en otros estudios (Husch et al., 2003). Los factores de expansión utilizan los volúmenes o existencias reales provenientes de los inventarios forestales para estimar biomasa y carbono (Schroeder et al., 1997; Gracia et al., 2004; Návar, 2009a). Los factores de expansión de biomasa difieren en función de muchas variables como las especies que componen el bosque, la edad, la estructura del rodal y la estación, y son aplicables factores diferentes a las existencias, el crecimiento y la extracción. La existencia de un conjunto centralizado de factores de expansión de biomasa y carbono de fácil acceso, con parámetros afines sobre los árboles, los rodales y las estaciones, podría favorecer la evaluación del carbono (FAO, 2009).
En México, existen grandes extensiones de áreas forestales con gran potencial para la conservación y la captura de carbono y que esta capacidad podría ser aprovechada y entendida (Masera et al., 2000; Návar et al., 2005). Sin embargo, hacen falta estos parámetros o índices que transformen las existencias reales, estimadas convencionalmente en planes de manejo forestal, en almacenes de carbono para poder entender y ajustar adecuadamente los planes y programas ambientales. Los objetivos de este trabajo de investigación fueron: a) determinar la concentración de carbono en los compartimientos de biomasa de árboles individuales y determinar si existen diferencias estadísticamente significativas entre los diferentes componentes de la madera, b) estimar el contenido de carbono almacenado en la biomasa aérea de bosques templados y c) estimar factores de expansión de carbono, d) modelar la dependencia de los factores en función de las características dasométricas de los árboles y definir su distribución probabilística. Las hipótesis intrínsicamente planteadas fueron: a) el contenido de carbono es similar en la albura y el duramen, b ) el contenido de carbono de la albura y el duramen es constante con la altura relativa del fuste en las especies de Pinus y del género Quercus, c) los factores de expansión de carbono son independientes de las características dasométricas del árbol y d) los factores de expansión de carbono se distribuyen normalmente en las especies de Pinus (P. arizonica Engelm., Pinus engelmannii, Pinus teocote Schiede ex Schlectendal et Chamisso y Pinus cooperi BlancoJ y del género Quercus (Quercus sideroxylla Humb & Bonpl. y Quercus rugosa Neé) estudiadas del norte de Durango, México.
MATERIALES Y MÉTODOS
Área de estudio. El presente trabajo se realizó en la comunidad El Tarahumar y en los ejidos El Tule y La Soledad, los cuales se ubican en la Sierra Madre Occidental en el municipio de Tepehuanes, Durango. Las coordenadas donde se localizan los predios son 25° 33' 48" a 25° 49' 07" latitud norte y de 106° 11' 04" a 106° 37' 22" de longitud oeste con respecto al meridiano de Greenwich (Figura 1). Los tipos de vegetación presentes en la zona corresponden a bosque de pino, bosque de pino–encino, bosque de encino y otras asociaciones de pino–otras coníferas y pino–otras hojosas, donde los bosques más importantes en el área son del género Pinus con una amplia distribución, encontrándose la mayor presencia del Pinus arizonica Engelmann, Pinus teocote Schiede ex Schlectendal et Chamisso y Pinus durangensis Martínez, siendo posible encontrar manchones puros de encino cuyas especies representativas son Quercus sideroxylla Humb. & Bonpl y Q. rugosa Neé. La región se encuentra incluida en la provincia fisiográfica denominada Sierra Madre Occidental en la subprovincia Sierra y Cañones Duranguenses. El clima que prevalece en la región que comprende el predio en estudio es el C(w0) y el C(w2), con una temperatura media anual de 10.5 °C y una precipitación media anual de 932 mm, con lluvias en verano. Los tipos de suelos en general que se encuentran en la región es un litosol podzolico.

Muestreo. Para la estimación de volumen y biomasa se eligieron los árboles que reunían características dasométricas representativas de los sitios, se derribaron 88 árboles en total, de los cuales 14 fueron de Pinus durangensis, 15 de Pinus arizonica, 16 de Pinus leiophylla, ocho de Pinus teocote, cinco de Pinus engelmannii, 19 de Pinus cooperi y 11 de Quercus spp. Para la estimación del contenido de carbono se analizaron cinco árboles muestra representativos del total de árboles derribados de cada una de las siguientes especies: P. arizonica, P. engemannii, P teocote, P. cooperi y Quercus spp.
A cada árbol se le midió en pie la variable diámetro normal. Posteriormente en árboles derribados se midió el diámetro a: 1) la base del fuste, 2) a 0,5 m, 3) a 1,3 m, 4) a 1,5 m y 5) a cada metro adicional, 2,5 m, 3,5 m, etc., hasta la punta del fuste principal. Aquí se midió la altura total del árbol. Posteriormente cada árbol fue separado en los componentes fuste y ramas, de los cuales se tomaron muestras, en el caso del fuste este fue cortado en trozas, a las cuales se les extrajeron rodajas. Se pesaron los componentes con sus muestras en campo por separado en estado verde. Las muestras fueron llevadas a laboratorio para ser analizadas por peso seco además de la densidad básica y la concentración de carbono.
Obtención de probetas. En cada árbol, el fuste se cortó en secciones de 2.5 m y se tomaron rodajas en cada sección a partir del tocón hasta la punta del árbol, registrando el nombre de la especie, número de árbol y altura de la sección. Posteriormente se trasladaron al laboratorio donde se obtuvo de cada rodaja una sección transversal de 6 cm de ancho y el largo en función del diámetro de la muestra. De esta muestra se tomaron una probeta de 2.5 x 2.5 x 2.5 cm de la albura y del duramen. Cada muestra por componente (albura y duramen) fue secada en la estufa hasta obtener un peso constante a una temperatura de 105 °C, ya secas se pesaron y se volvieron a medir. De cada muestra seca, se tomó una submuestra y se pulverizó en un molino. Cada muestra se colocó en una bolsa de plástico, la cual fue identificada con un registro.
Determinación del contenido de carbono. Para la determinación del contenido de carbono se utilizó el equipo anatítico Solids TOC Analizer, el cual permite identificar niveles de 0.05 mg – 30 mg de carbono en muestras sólidas. Como primer paso se realizó la calibración del equipo con un estándar de sacarosa al 30 % con cuatro volúmenes de la solución, obteniendo un r2 en la recta de calibración de 0.999. Una vez calibrado el equipo se analizó cada uno de los componentes utilizando una cantidad de 30 mg de la submuestra molida.
Conversión de volúmenes verdes a peso seco.
Para la conversión de volumen a biomasa se utilizaron dos procedimientos. El primero se realizó por medio de las ecuaciones de biomasa descritas en el Cuadro 1. La biomasa total por unidad de superficie se determinó con la suma de la biomasa del fuste y de las ramas de cada árbol dentro del sitio de muestreo en el inventario forestal. Para el cálculo de la biomasa del fuste y de las ramas de cada árbol se utilizó el diámetro normal de los árboles, consistente con la aplicación de las ecuaciones alométricas de biomasa. Este procedimiento resulta en toneladas de biomasa arbórea por unidad de superficie (Mg ha–1).

El segundo procedimiento se realizó por medio del uso de la densidad básica de la madera de la siguiente forma:
B = V * p
Donde: B= Biomasa (Mg·ha–1), V= Volumen (m3·ha–1), p = Densidad básica de la madera (g·cm–3 o Mg·m–3).
La densidad básica de la madera es una variable aleatoria a lo largo del árbol y del centro del fuste hacia la periferia. Para determinar la densidad básica promedio de la madera se utilizaron las ecuaciones descritas en el Cuadro 2.

Para determinar el volumen de ramas y fuste se utilizaron las ecuaciones descritas en el Cuadro 3.

Estimación de la biomasa y cantidad de carbono por hectárea. Para la determinación de la densidad de carbono por hectárea la biomasa estimada al nivel del sitio se multiplicó por la concentración promedio de carbono y fue expresada en Mg ha–1.
Estimación de los factores de expansión de carbono. Para la estimación de los factores de expansión de carbono por unidad de superficie se utilizó la siguiente fórmula:

Donde: FEC= Factor de expansión de carbono (expresado en Mg·m–3), CC = Contenido de carbono total (expresado en Mg·ha–1), V= Existencia reales por hectárea o Volumen por hectárea para el rodal i (estimado mediante la ecuaciones de volumen aplicada a cada uno de los árboles inventariados del sitio o rodal).
De esta manera existe un factor de expansión de carbono por cada sitio o rodal inventariado. Se utilizaron datos provenientes de 98 rodales inventariados y por consiguiente se estimaron factores de expansión para cada uno de los 98 rodales. Los factores de expansión se relacionaron con los datos dasométricos al nivel del rodal de la siguiente forma: FEC = f ( , Densidad, Diámetro). Se utilizaron programas gráficos de excell y Sigma Plot para observar la tendencia de los datos y posteriormente se utilizó el programa de cómputo SAS para determinar las ecuaciones correspondientes en donde las ecuaciones que se ajustaron fueron del tipo lineal y no lineal. Adicionalmente, los factores de expansión para los rodales estudiados se ajustaron a la distribución probabilística Normal.
, Densidad, Diámetro). Se utilizaron programas gráficos de excell y Sigma Plot para observar la tendencia de los datos y posteriormente se utilizó el programa de cómputo SAS para determinar las ecuaciones correspondientes en donde las ecuaciones que se ajustaron fueron del tipo lineal y no lineal. Adicionalmente, los factores de expansión para los rodales estudiados se ajustaron a la distribución probabilística Normal.
RESULTADOS
Contenido de carbono en probetas. La concentración del contenido de carbono promedio obtenido para los géneros Pinus spp y Quercus spp fue de 0.56 y 0.54; con desviaciones estándares e intervalos de confianza (p=0.05) de 0.0373 y 0.0256 y de 0.038 y 0.052, respectivamente. El contenido de carbono promedio de la biomasa obtenido en este estudio se encuentra dentro de los límites reportados por Gutiérrez y Lópera (2001) 51.20 %; Ramírez et al. (1997) 50 – 53 %; Green et al. (2005) 52 % y Husch et al. (2003) 45 – 55 %. La diferencia en los valores obtenidos del contenido de carbono promedio para los géneros Pinus spp y Quercus spp concuerda con lo descrito en la literatura científica donde se reporta que el contenido de carbono en coníferas es más alto que en latifoliadas (Gracia et al., 2004).
Los resultados indican que no existen diferencias estadísticamente significativas en la concentración de carbono entre los componentes duramen y albura para los géneros Pinus y Quercus (P>F = 0.1945) y la altura relativa del árbol (P>F = 0.9297; P>F = 0.9119). Sin embargo, la concentración de carbono resultó ser estadísticamente diferente entre las especies estudiadas de Pinos (P. arizonica, P. cooperi, P. engelmanii y P. teocote) y encinos (Quercus spp) (P>F = 0.0001). Esto concuerda con lo reportado por Pearson y Williams (1992), quienes señalan que las variaciones entre los árboles son más significativas que la variación dentro del mismo árbol.
Los resultados gráficos con el promedio de concentración de carbono para los componentes de albura y duramen en relación con las especies estudiadas de pino (P. arizonica, P. cooperi, P. engelmanii y P. teocote) y encino (Quercus spp) del norte de Durango, México se presentan en la Figura 2. La concentración de carbono cambia en relación con las especies de pino y encino (Quercus spp) estudiadas (P>F= 0.0001). La especie que mayor concentración de carbono tuvo fue el P. engelmannii, seguido del P. teocote, P. cooperi, Quercus spp y P. arizonica, cuyos valores se presentan gráficamente a continuación.

Los factores de expansión de carbono. El factor de expansión de carbono promedio estimado mediante la ecuación de biomasa para el género Pinus spp fue de 0.42 Mg C·m–3 donde el factor mínimo y máximo fue de 0.31 y 0.50 Mg C·m–3. Para el género Quercus este valor fue de 0.57 Mg C·m–3 y sus valores mínimo y máximo fueron de 0.32 y 0.82 Mg C·m–3. Los valores de volumen, biomasa, carbono y factor de expansión de carbono promedios para los géneros Pinus y Quercus se muestran en el Cuadro 4.
El factor de expansión de carbono promedio estimado utilizando la densidad de la madera y el volumen fustal fue de 0.36 para el género Pinus spp donde el factor mínimo y máximo fue de 0.28 y 0.48 y para el género Quercus fue de 0.99 mientras que sus valores mínimo y máximo son 0.55 a 1.48. Este procedimiento sobreestima, en contraste con el primer procedimiento que utiliza las ecuaciones alométricas, el valor en el género Quercus con 0.42 Mg·m–3 (73.68 %) y lo subestima en el género Pinus con 0.06 Mg·m–3 (14.28 %). Los valores de volumen, biomasa, carbono y factor de expansión de carbono promedios para los géneros Pinus spp y Quercus spp se muestran en el Cuadro 5.
A partir de las estimaciones de biomasa realizadas mediante los dos procedimientos se concluye que es más recomendable utilizar las ecuaciones de biomasa porque el uso de un solo valor de densidad de la madera es demasiado simple para estimar la biomasa de un árbol.
Con el uso de las ecuaciones alométricas, el contenido de carbono promedio presente en la biomasa aérea en los ecosistemas templados del norte de Durango, México en el género Pinus spp fue de 41.29 Mg C·ha–1 con valores mínimo y máximo de 19.92 a 95.62 Mg C·ha–1, y el carbono total promedio contenido en la biomasa en el género Quercus fue de 14.07 Mg C·ha–1 con valores mínimo y máximo de 1.63 y 44.93 Mg C·ha–1. Utilizando la ecuación que incluye el uso de la densidad de la madera, el contenido de carbono promedio presente en la biomasa aérea en los ecosistemas templados del norte de Durango, México en el género Pinus spp fue de 35.85 Mg C·ha–1 con valores mínimo y máximo de 17.36 a 69.39 Mg C·ha–1, y el carbono total promedio contenido en la biomasa en el género Quercus fue de 23.62 con valores mínimo y máximo de 6.49 y 68.64 Mg C·ha–1.
Los factores de expansión de carbono estimados mediante el primer procedimiento se relacionaron con la densidad básica de la madera y con la altura del árbol en el género Pinus spp. Aunque todas las relaciones tuvieron coeficientes de determinación bajos, éstas fueron estadísticamente significativas (P<0.05). La relación del factor con la altura promedio del rodal se muestra en la Figura 3. Los factores de expansión de carbono cambian con la altura en las especies de pino estudiadas (P. arizonica, P. cooperi, P. engelmanniiy P. teocote) (P>F= 0.0429) y se ajusta la tendencia a una línea recta con pendiente negativa (r2=0.22 y Sx=0.10). Conforme aumenta la altura el factor de expansión de carbono disminuye, indicando que por cada unidad de altura del árbol el factor de expansión de carbono en el género Pinus spp cambia por –0.00816438 unidades.

La relación entre los factores de expansión de carbono con la altura del árbol en el género Quercus se muestra en la Figura 4. Los factores de expansión de carbono cambian con la altura en las especies de encino estudiadas (Quercus sideroxylla y Quercus rugosa) (P>F= 0.0025) y se ajusta la tendencia a una línea recta con pendiente negativa (r2=0.01 y Sx=0.21). Conforme aumenta la altura el factor de expansión de carbono disminuye, indicando que por cada unidad de altura del árbol el factor de expansión de carbono en el género Quercus cambia por –0.00256832 unidades.

Los factores de expansión de carbono cambian con la densidad del árbol en las especies de pino estudiadas (P>F=0.0054) y se ajusta la tendencia a una línea recta con pendiente negativa (r2=0.19 y Sx=0.10) (Figura 5). Esto indica que por cada unidad de densidad de la madera el factor de expansión de carbono en el género Pinus spp cambia 14.3875. Estos resultados concuerdan con los obtenidos en los factores de expansión de biomasa, que a mayor densidad el factor de expansión de carbono disminuye.

La relación de los factores de expansión de carbono con la densidad del árbol del género Quercus spp del norte de Durango, México se presenta en la Figura 6. Los factores de expansión de carbono cambian con la densidad del árbol en el género Quercus (P>F= 0.0119) y se ajusta la tendencia a una línea recta con pendiente positiva (r2=0.007 y Sx=0.20). Esto indica que por cada unidad de densidad de la madera el factor de expansión de carbono en el género Quercus cambia 46.7568. Estos resultados concuerdan con los obtenidos en los factores de expansión de biomasa, en donde el factor de expansión de carbono aumenta conforme aumenta la densidad.

La distribución de los factores de expansión de carbono en el género Pinus spp se muestra en la Figura 7. Los factores de expansión de carbono en el género Pinus spp se distribuyen normalmente (P ≥ χ2 = 0.5114).

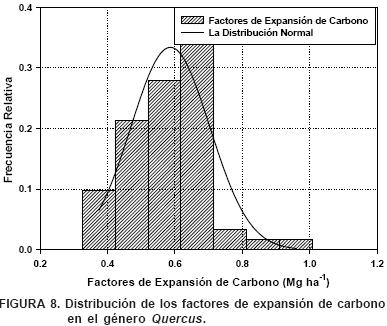
La distribución de los factores de expansión de carbono en el género Quercus spp se muestra en la Figura 8. Los factores de expansión de carbono en el género Quercus spp también se distribuyen normalmente (P ≥ χ2 = 0.5142).
DISCUSIÓN
Návar–Cháidez (2009) reportó que los factores de expansión para bosques provenientes de la selva baja caducifolia de Sinaloa, México se distribuyen Weibull y no normalmente en contraste con las observaciones de este estudio. Este investigador registró densidades de carbono en bosques tropicales secos de Sinaloa, México valores promedio de 36.5 Mg C·ha–1 (3.5 Mg C·ha–1). Los resultados de las estimaciones de este estudio se encuentran por encima de los reportados Zamora (2003) en bosques de pino en Michoacán, con valores de 35.6 Mg C·ha–1 en pino y 6.56 Mg C·ha–1 en encino. Este investigador cita que la diferencia en los valores del contenido de carbono entre un rodal y otro pueden deberse al volumen por hectárea, superficie y densidad de la biomasa. Para bosques templados de Durango, México, Návar (2009a) reportó en promedio (intervalo de confianza) la densidad de carbono al nivel del rodal de 41.17 (0.54), 19.6 (0.25), y 67.16 (0.87) Mg C·ha–1 para fustes, ramas y biomasa aérea total sobre el suelo, respectivamente. Las ramas explican aproximadamente el 47 y el 29 % de la biomasa fustal y total aérea, respectivamente. Los porcentajes son un poco diferentes que aquellos valores reportados para plantaciones de Gmelina arbórea por Swami et al. (2003), quienes registraron valores de 55, 19, 19, y 7 % para fustes, ramas, raíces y follaje, respectivamente. La densidad de carbono en promedio para los bosques del estado de Nuevo León es de 37.1 Mg·ha–1 (±6 Mg·ha–1), con una desviación estándar de 30 Mg·ha–1 (Návar, 2010). Para bosques tropicales se registran valores de 112 Mg·ha–1 para la Península de Yucatán (Cairns et al., 2000) y 133 Mg ha–1 para Veracruz (Cairns et al., 2000). Para la selva tropical seca de Jalisco, México se reporta un valor de 37 Mg·ha–1 (Castellanos et al., 1991) o de 47–62 Mg ha–1 para un bosque tropical del centro de México (Jaramillo et al., 2003).
Esta revisión de literatura indica que las densidades de carbono son muy variables entre diferentes comunidades forestales y entre la misma comunidad forestal en diferentes localidades. Por esta razón se deben de extrapolar las densidades de carbono de bosques similares de localidades lo más adyacentes posibles al lugar del estudio. En la ausencia de trabajos de esta naturaleza para las comunidades forestales deseadas, se recomienda utilizar los factores de expansión de biomasa, utilizando las ecuaciones de biomasa reportadas para bosques Norteamericanos (Ter–Mikaelian y Korzukhin, 1997) o para bosques mundiales (Zianis y Mencuccini, 2004) o para bosques Latino Americanos (Návar, 2009b),
CONCLUSIONES
Los resultados de este estudio indican que en promedio las concentraciones de carbono varían de 54 y 56 % del peso seco de la biomasa para los componentes albura y duramen. Las concentraciones de carbono son estadísticamente diferentes entre las especies estudiadas de Pinos (Pinus arizonica, Pinus cooperi, Pinus engelmannii y Pinus teocote) y encinos (Quercus sideroxylla y Quercus rugosa). Se debe parcialmente a los cambios de densidad de cada una de las especies. La densidad de carbono promedio (intervalo de confianza) en la biomasa aérea de los bosques templados del norte de Durango, México es de 57.12 (0.24) Mg C·ha–1, pero este valor varía en función de la mezcla de pinos y encinos. A pesar de que los factores de expansión de carbono en los géneros Pinus spp y Quercus spp se distribuyen normalmente, se debe tener precaución con el uso de estos valores porque cambian en función de la altura promedio del rodal y de la densidad de la madera del árbol. También cambian en función de las diferentes ecuaciones utilizadas, con ecuaciones alométricas o con la función de volumen por la densidad. En general se recomienda el procedimiento que utiliza las ecuaciones de biomasa en el cálculo de las densidades de carbono en rodales forestales.
LITERATURA CITADA
BROWN, S. L.; SCHROEDER, P. E. 2000. Spatial patterns of aboveground production and mortality of woody biomass for eastern U.S. forests: Erratum. Ecological Applications 10:937–945. [ Links ]
CAIRNS, M. A.; DIRZO R. R.; ZADROGA F. 1995. Forests of Mexico: a diminishing resource? Journal of Forestry 93:21–24. [ Links ]
CAIRNS, M. A.; HAGGERTY, P. K.; ALVAREZ, R.; DE JONG B.H.J.; OLMSTED, E. I. 2000. Tropical Mexico's Recent Land–Use Change: A Region's Contribution to the Global Carbon Cycle. Ecological Applications 10: 1426–1441. [ Links ]
CASTELLANOS, J.; MASS M.; KUMMEROW, J. 1991. Root biomass of a dry deciduous tropical forest in Mexico. Plant and Soil 131: 18–33. [ Links ]
DE JONG, B. H. J.; CAIRNS, M. A.; HAGGERTY, P. K.; RAMIREZ–MARCIAL, N.; OCHOA–GAONA, J.; MENDOZA–VEGA, J.; GONZÁLEZ–ESPINOSA, M.; MARCH–MIFSUT, I. 1999. Land–use change carbon flux between 1970's and 1990's in central highlands of Chiapas, Mexico. Environmental Management 23: 373–385. [ Links ]
DE JONG, B.H.J.; OCHOA–GAONA, S.; CASTILLO–SANTIAGO, M. A.; RAMIREZ–MARCIAL N.; CAIRNS M. A. 2000. Carbon flux and patterns of land–use/land–cover change in the Selva Lacandona, Mexico. Ambio 29: 504–511. [ Links ]
DIRZO R.; GARCÍA, C. 1991. Rates of deforestation in Los Tuxtlas, a netropical area in southeast Mexico. Conservation Biology 6: 84–90. [ Links ]
FAO. Food and Agriculture Organization. 2009. Estado de los Bosques del Mundo. Rome, Italy. 215 p. [ Links ]
GRACIA, C.; VAYREDA, J.; SABATÉ S.; IBAÑEZ J. 2004. Main components of the aboveground biomass expansion factors. CREAF, Centre de Recerca Ecological Aplicacions Forestals. University of Barcelona, Spain. 24 p. [ Links ]
GREEN, C.; BRIAN, T.; O'SHEA, M.; FARELL, E.; BYRNE, A. 2005.Above–and belowground biomass measurements in an unthinned stand of Sitke spruce (Picea stchensis (Bong) Carr.). Forest Ecology and Management 10: 937–945. [ Links ]
GUTIÉRREZ, V.; LÓPERA G. 2001. Metodología para la cuantificación de existencias y flujo de carbono en plantaciones forestales. Simposio Internacional Medición y Monitoreo de la captura de Carbono en Ecosistemas Forestales. Valdivia, Chile. 12 p. [ Links ]
HUSCH, B.; BEERS, T. W.; KERSHAW, J. A. 2003. Forest Mensuration. John Wiley & Sons. Hoboken, New Jersey. USA. 433 p. [ Links ]
JARAMILLO, V. J.; KAUFFMAN, J. B.; RENTERÍA–RODRÍGUEZ, L.; CUMMINGS, D. L.; ELLINGSON, L. J. 2003. Biomass, Carbon, and Nitrogen Pools in Mexican Tropical Dry Forest Landscapes. Ecosystems 6: 609–629. [ Links ]
MASERA, O.; DE JONG, B.; RICALDE, I. 2000. Consolidación de la Oficina Mexicana para la Mitigación de Gases de Efecto Invernadero. Sector Forestal. Estudio Elaborado para el Instituto Nacional de Ecología con fondos del Banco Mundial. ECOSUR. México. 197 p. [ Links ]
NÁVAR, J.; GONZÁLEZ, N.; GRACIANO, J. 2005. Carbon sequestration by forest plantations of Durango, México. Madera y Bosques 11: 15–34. [ Links ]
NÁVAR–CHÁIDEZ, J. J. 2008. Carbon fluxes resulting from land–use changes in the Tamaulipan thornscrub of northeastern Mexico. Carbon Balance and Management 2008 3:6 doi10.1186/1750–0680–3–6. [ Links ]
NÁVAR–CHÁIDEZ, J. J. 2009. Allometric equations and expansion factors for tropical dry trees of eastern Sinaloa, México. Tropical and Subtropical Agroecosystems 10: 45–52. [ Links ]
NÁVAR, J. 2009a. Allometric equations for tree species and carbon stocks for forests of northwestern Mexico. Forest Ecology and Management 257: 427–434. doi:10.1016/ j.foreco.2008.09.028. [ Links ]
NAVAR, J. 2009b. Biomass component equations for Latin American species and groups of species. Annals of Forest Science 66:1–8. doi:10.1051/forest/2009001. [ Links ]
NAVAR, J. 2010. Los bosques templados del estado de Nuevo León: el manejo sustentable para bienes y servicios ambientales. En Prensa en Madera y Bosques. [ Links ]
PEARSON, R.; WILLIAMS, J. 1992. Review of Methods for Sampling of Timber. Forest Products Journal 8:263–268. [ Links ]
RAMÍREZ, O.; GÓMEZ, M.; SHULTZ, S. 1997. Valuing the contribution of plantation forestry to the national accounts of Costa Rica from the ecological economics perspective. Beijer Research Seminar. Costa Rica. 28 p. [ Links ]
SCHROEDER, P.; BROWN, S.; BIRDSEY, R.; CIESZEWSKI, C. 1997. Biomass estimation for temperate broadleaf forests of the United States using inventory data. Forest Science 43: 424–434. [ Links ]
SECRETARÍA DEL MEDIO AMBIENTE Y RECURSOS NATURALES (SEMARNAT). 2001. Estadísticas de Deforestación en México: 1993–2000. México, D. F. [ Links ]
SILVA–ARREDONDO, F. M.; PRIETO–RUIZ, J. A.; NÁVAR–CHÁIDEZ, J. J. 2009. Estimación de la densidad de la madera en comunidades forestales templadas del norte de Durango, México. En Prensa en Madera y Bosques. [ Links ]
SWAMI, S. L.; KUSHWAJA, S. K.; PURI, S. 2003. Tree growth biomass, allometry and nutrient distribution in Gmelina arborea stands grown in red lateritic soils of Central India. Biomass and Bioenergy 26: 305–317. [ Links ]
TER–MIKAELIAN, M. T.; KORZUKHIN, M. D. 1997. Biomass equations for 65 North American tree species. Forest Ecology and Management 97: 1–24. [ Links ]
ZIANIS, D.; MENCUCCINI, M. 2004. On simplifying allometric analyses of forest biomass. For. Ecol. Manage. 187: 311–332. [ Links ]

