










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.15 no.2 Chapingo jul./dic. 2009
Caracterización estructural del matorral submontano de la Sierra Madre Oriental, Nuevo León, México
Structural characterization of submontane scrub of the Sierra Madre Oriental, Nuevo Leon, Mexico
P. A. Canizales–Velázquez1; E. Alanís–Rodríguez1; R. Aranda–Ramos2; J. M. Mata–Balderas2; J. Jiménez–Pérez1; G. Alanís–Flores3; J. I. Uvalle–Sauceda1; M. G. Ruiz–Bautista2
1 Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León, MÉXICO. Correo–e: pamcanizales@hotmail.com.
2 Sistemas de Innovación y Desarrollo Ambiental, S. C.
3 Facultad de Ciencias Biológicas, Universidad Autónoma de Nuevo León. MÉXICO.
Recibido: 5 de junio, 2009
Aceptado: 28 de julio, 2009
RESUMEN
Las investigaciones concernientes al matorral submontano son escasas y se desconoce la riqueza y diversidad de sus recursos naturales. El estudio se desarrolló en siete zonas de la Sierra Madre Oriental (SMO) con historial de extracción selectiva de especies leñosas, con el objetivo de realizar la caracterización vertical y horizontal del matorral submontano, así como determinar la diversidad a. Para el análisis se establecieron 73 sitios de muestreo de 100 m2 sobre un gradiente altitudinal de 600 a 800 m, en los cuales se evaluaron todas las especies leñosas recabando información dasométrica como altura (h), diámetro (d0.10m) y cobertura de copa (N–S, E–O). Se determinaron los parámetros ecológicos de abundancia (Ar), dominancia (Dr), frecuencia (Fr) e índice de valor de importancia (IVI), así como la estimación de los índices de Shannon (H') y Margalef (Da). Se registraron 23 familias, distribuidas en 52 especies y 42 géneros; la familia más representada fue Fabaceae con 15 especies. El ecosistema se conforma principalmente por las especies Acacia rigidula, Bernardia myricaefolia y Acacia berlandieri. Presenta un índice de diversidad de H'=3, 0, lo cual indica que es un ecosistema muy diverso en relación a otros tipos de ecosistemas del noreste de México.
Palabras clave: Acacia berlandieri, Acacia rigidula, Bernardia myricaefolia, Fabaceae, noreste de México.
ABSTRACT
Investigations about the submontane scrub ecosystem are insufficient and the richness and diversity of its natural resources are still poorly known. This study was conducted in seven areas of the Sierra Madre Oriental (SMO) with a history of selective removal of woody species, in order to assess the characterization of the vertical and horizontal submontane scrub ecosystem, and determine diversity aa. Sampling was carried out in 73 sites of 100 m2 in an altitudinal range of 600–800 m, in which all woody species were assessed by obtaining dasometric information height (h), diameter (d0.10m) and cover (NS, EO). The ecological parameters of abundance (Ar), dominance (Dr), frequency (Fr), importance value index (IVI) were determined, as well as, the Shannon (H') and Margalef (Da) indices. A total 23 families distributed in 52 species and 42 genera were found, Fabaceae being represented with 15 species. The most representative species were Acacia rigidula, Acacia berlandieri and Bernardia myricaefolia. A diversity index H'= 3, 0, indicates that is submontane scrub is very diverse in relation to other types of ecosystems of northeastern Mexico.
Keywords: Acacia berlandieri, Acacia rigidula, Bernardia myricaefolia, Fabaceae, northeastern Mexico.
INTRODUCCIÓN
México es uno de los cinco países que posee mayor diversidad biológica en el mundo (Mittermeier et al., 1997; SEMARNAT, 2006). Las regiones áridas y semiáridas de México son altamente diversas desde el punto de vista fisonómico (Rzedowski, 2006). El matorral submontano (Alanís et al., 1996) considerando como una variante del matorral xerófilo (Rzedowski, 2006) es una formación arbustiva y subarbórea rica en sus formas de vida, característica de estas regiones (Alanís et al., 1996).
El matorral es el ecosistema más abundante e históricamente más utilizado en las zonas áridas y semiáridas de México (García y Jurado, 2008). A lo largo de la historia se ha visto afectado por actividades antropogénicas, como extracción de especies vegetales para diferentes usos (seto, combustible, textil, medicinal y alimenticio) (Estrada et al., 2004; Rzedowski, 2006; García y Jurado, 2008) de la misma manera ha sufrido de continua deforestación para establecer zonas de uso agrícola, industrial y urbano (Alanís et al., 2008; Arriaga, 2009). Por otra parte, el uso ganadero es la práctica más frecuente, siendo el efecto más notable del pastoreo, la substitución paulatina de las plantas nativas (Rzedowski, 2006; García y Jurado, 2008). Tan sólo en los años de 1993 al 2002 el matorral sufrió una pérdida de 953 mil ha por cambio de uso de suelo, siendo el segundo ecosistema más afectado en México después de las selvas (SEMARNAT, 2006), y actualmente se ha observado un incremento en el cambio de uso suelo para el establecimiento de zonas urbanas.
Sin embargo los matorrales juegan un papel muy importante al ser uno de los principales tipos de vegetación del país, ya que cubre alrededor del 28,55 % de la superficie del territorio mexicano (Palacio et al., 2000). Por su parte el matorral submontano se encuentra en el noreste de México distribuido a lo largo de la Sierra Madre Oriental (SMO), desde Nuevo León hasta Hidalgo (Rzedowski, 2006), ocupando aproximadamente 11 % de la superficie del estado de Nuevo León (Palacio et al., 2000) albergando gran diversidad de organismos; algunas especies que lo conforman fungen como agentes en el control de la erosión (Foroughbakhch et al., 2003) y efectúa una función primordial como área de transición entre las zonas de matorral desértico en las partes bajas y los bosques de encino–pino existentes en los taludes superiores de la SMO (Alanís et al., 1995; Alanís et al., 1996). Para el estado de Nuevo León existen listados florísticos de matorral submontano realizados por diversos autores (INEGI, 1986; Alanís et al., 1995; Alanís et al., 1996; Estrada et al., 2004) pero todavía existe poca información en relación a la caracterización estructural de dicho ecosistema (García y Jurado, 2008), por lo cual es necesario generar más información, así como sobre su diversidad y riqueza de especies (Arriaga, 2009). Los objetivos planteados en la presente investigación fueron los siguientes: 1) estimar la diversidad del matorral submontano en la SMO dentro del estado de Nuevo León, 2) evaluar la estructura vertical y 3) estimar los parámetros ecológicos de abundancia (Ai), dominancia (Di), frecuencia (Fi) e índice de valor de importancia (IVI).
MATERIALES Y MÉTODOS
El área de estudio se encuentra dentro de la cuenca hidrológica Río Bravo–San Juan, posee elevaciones de 6001,400 m, el clima predominante es el semicálido, semiárido con una temperatura media anual mayor a 18 °C (BS1hw). En su mayoría los suelos son tipo rendiza y litosol, y en menor proporción se encuentran castañozem lúvico y feozem calcárico (INEGI, 1986).
La comunidad vegetal representativa es el matorral submontano conformado principalmente por las especies Helietta parvifolia, Cordia boissieri, Havardia pallens y Acacia rigidula (Alanís et al., 1996; García y Jurado, 2008). (cuadro 1).
El muestreo consistió en el establecimiento de 73 sitios de 100 m2 cada uno; con la finalidad de obtener mayor representatividad del área se distribuyeron en siete zonas de la SMO donde no se tiene registro de actividades agrícolas o pecuarias, sólo la extracción selectiva de sus componentes leñosos. Los sitios de muestreo se localizan en los municipios de Monterrey, San Pedro Garza García, Guadalupe y General Escobedo, a una altitud de 600 a 800 m (Figura 1). Se incluyeron todas las especies leñosas > 1 cm de diámetro (d0.10m), las especies herbáceas se descartaron dado que la época de muestreo no era adecuada para monitorear este grupo. Se obtuvo información dasométrica como: especie (basado en Alanís et al. (1996)), altura (h), diámetro (d0.10m) y cobertura de copa (norte–sur, este–oeste). Mediante el análisis de la información se derivaron parámetros estructurales de la vegetación como: abundancia relativa (Ai), frecuencia relativa (Fi), área basal (G), dominancia relativa (Di) e índice de valor de importancia (IVI) (Curtis y McIntosh, 1950; Mueller y Ellenberg, 1974; Magurran, 2004), los cuales se estimaron mediante las siguientes ecuaciones matemáticas:

La abundancia relativa se obtuvo mediante la fórmula (1).

Donde Ai es la abundancia relativa de la especie i con respecto a la abundancia total, n es el número de individuos de la especie i y N el número total de individuos.
La dominancia relativa se obtuvo mediante la ecuación (2).


Donde Di es la dominancia relativa de la especie i respecto a la dominancia total, g es el área basal de la especie i, siendo G el área basal total, ecuación (3).
La frecuencia relativa se obtuvo mediante la fórmula (4).

Donde Fi es la frecuencia relativa de la especie i con respecto a la frecuencia total, m la frecuencia de la especies i en los sitios de muestreo y M el número total de sitios de muestreo.
El índice de valor de importancia se obtuvo para cada especie, mediante la ecuación (5).

Donde Ai es la abundancia relativa, Di es la dominancia relativa y Fi la frecuencia relativa.
Para estimar la diversidad se utilizó el índice de Shannon (Shannon, 1948) y el índice de Margalef (Clifford y Stephenson, 1975). El índice de Shannon se estimó mediante la ecuación (6).

Donde S es el número de especies presentes, ln es logaritmo natural y pi es la proporción de las especies.
pi=ni/N; donde ni es el número de individuos de la especie i y N es el número total de individuos.
El índice de diversidad de Margalef se estimó con la ecuación (7).

De acuerdo a Pretzsch (1996) se interpretó la distribución de las especies en tres diferentes zonas de altura: zona I: 80–100 % de la altura máxima de la población, zona II: 50–80 %, zona III: 0–50%. Posteriormente se calculó el índice de distribución vertical de especies (A), el cual cuantifica la diversidad de especies y su ocupación en el espacio vertical en la población (Estrada et al., 2005). El índice de distribución vertical se calculó de acuerdo a la fórmula (8).

Donde S es el número de especies presentes; Z el número de zonas de altura y pij (9) la proporción de especies en cada zona de altura, n es el número de individuos de la misma especie (i) en la zona (j) y N el número total de individuos.
RESULTADOS Y DISCUSIÓN
Parámetros ecológicos
Se registraron un total de 23 familias distribuidas en 52 especies y 42 géneros, la familia Fabaceae fue la mejor representada con 15 especies, cantidad menor a la registrada por Estrada et al. (2004) para la región centro de Nuevo León y cantidad menor para la región norte del estado de Nuevo León por Estrada et al. (2005), pero mayor para la registrada por García y Jurado (2008) en el matorral submontano con condiciones prístinas en Linares. Los elementos registrados son típicos de matorral submontano particularmente en el estado de Nuevo León (Alanís et al., 1996; Estrada et al., 2004; García y Jurado, 2008). Las especies más abundantes son Acacia rigidula (713 n/ha), Acacia berlandieri (320 n/ha), típicas del matorral submontano (Alanís et al., 1996; Estrada et al., 2005; García y Jurado, 2008) y Bernardia myricaefolia (441 n/ha), la cual no destaca como especie predominante de este ecosistema para el estado de Nuevo León (Alanís et al., 1996; Estrada et al., 2005; Rzedowski, 2006; García y Jurado, 2008), a diferencia de lo reportado por García y Jurado (2008) para el matorral submontano en condiciones prístinas donde Helietta parvifolia, Randia rhagocarpa y Acacia rigidula son las más abundantes.
Las especies dominantes son Cordia boissieri (Di=15,52 %) lo cual concuerda con lo mencionado por Rzedowski (2006); así mismo se reportan a Acacia rigidula (D =10,42 %) y Zanthoxylum fagara (D=8,17 %) como dominantes, a diferencia por lo encontrado por García y Jurado (2008) que señalan a Helietta parvifolia, Gochnatia hypoleuca y Acacia greggii como dominantes. Así mismo, Alanís (1995), Estrada et al. (2005) y Rzedowski (2006) refieren a Helietta parvifolia como una de las especies dominantes del matorral submontano del noreste del país; sin embargo, en este estudio presentó dominancia baja (0,06 %). La disminución de las poblaciones de Helietta parvifolia puede deberse principalmente a la importancia forestal de ésta, ya que a través de los años ha sido extraída para su utilización en la elaboración de viviendas, cercas y corrales, debido a la alta durabilidad y resistencia de la madera, además de ser utilizada también como planta forrajera (Foroughbakhch et al., 2003), motivos por los cuales se ha visto reducida a un pequeño número de individuos aislados; caso contrario a lo mencionado por García y Jurado (2008) debido a que su estudio fue realizado en matorral submontano en condiciones prístinas, por lo cual Helietta parvifolia se presentó como la especie más abundante, predominante y de mayor valor ecológico. Para el presente estudio las especies con mayor índice de valor de importancia son Acacia rigidula (IVI=35,13 %) (similar a lo reportado por Alanís et al., 1995; García y Jurado, 2008), Cordia boissieri (IVI=30,45 %) (Alanís et al., 1995; Rzedowski, 2006) y Zanthoxylum fagara (IVI=21,59 %).
Diversidad α
El matorral submontano estudiado presentó una diversidad de H'=3.0 de acuerdo al índice de Shannon, lo cual indica que es un ecosistema muy diverso en relación a otros tipos de ecosistemas del noreste de México (Treviño et al., 2001; Corral et al., 2005; Alanís et al., 2008). El índice de riqueza de especies de Margalef (Da) indicó un valor de 6.34 y una riqueza específica (S) de 52, lo cual indica que es relativamente alta. Lo anterior debido a que la extracción selectiva de algunos de sus componentes leñosos ha ocasionado que el ecosistema evaluado presente diferentes estadíos sucesionales conformando un mosaico donde se registraron desde especies pioneras con valores dasométricos bajos, hasta individuos adultos característicos de zonas prístinas, contrario a lo encontrado por García y Jurado (2008) quienes registraron una riqueza de tan sólo 16 especies en condiciones prístinas, ya que en un ecosistema sin ningún tipo de influencia, el reemplazo de especies se da en condiciones naturales de manera lenta (Arriaga, 2009). La relativa alta riqueza se deriva del hecho de que algunos sitios estudiados incluyen elementos del ecosistema de bosque de encino debido que el matorral submontano es un ecosistema de transición por lo cual algunos sitios constituyen ecotonos con el bosque de encino, y se presentan especies del mismo como Quercus rysophylla, Q. polymorpha, Q. laeta, Q. canbyi. Así mismo se presentan especies del matorral xerófilo colindante ubicado a altitudes menores que el matorral submontano.
Estructura vertical
En el análisis de la estructura vertical mediante el índice vertical de especies (A) éstas se distribuyen en tres estratos. El estrato I (alto) se encuentra conformado por diversas especies del género Quercus (Q. canbyi, Q. laeta y Q. polymorpha, Celtis laevigata, Platanus occidentalis y Yucca filifera, especie con mayor número de individuos en el estrato y que obtuvo la altura máxima (hmax= 13 m) del ecosistema. El estrato II (medio) está conformado casi en su totalidad por Yucca filifera, Diospyros texana, Quercus rysophylla, Havardia pallens y Platanus occidentalis, siendo Yucca filifera la predominante, con una altura máxima de 9.7 m para el estrato. Por su parte el estrato III (bajo) que fue el mejor representado, se encuentra conformado principalmente por Acacia rigidula, Bernardia myricaefolia, Acacia berlandieri, Cordia boissieri y Zanthoxyllum fagara, alcanzando la altura máxima para el estrato Cordia boissieri con 6 m.
De acuerdo a la caracterización vertical del matorral submontano realizada por García y Jurado (2008), la altura máxima fue de 6 m para Helietta parvifolia, Cordia boissieri y Fraxinus greggii; en el presente estudio esta altura se alcanzó en el estrato III, debido a la presencia de especies altas como Yucca filifera y Quercus spp., no registradas en el estudio de García y Jurado (2008) y a que Helietta parvifolia en el presente estudio posee alturas inferiores a 6 m (1.7 a 2.4 m).
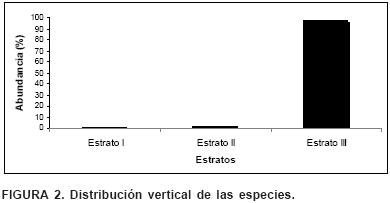
El estrato I posee una abundancia de 0.41 %, el estrato II de 1.07 % y el estrato III de 97.04 % (Figura 2), lo que indica que el ecosistema de matorral submontano con extracción selectiva de sus componentes leñosos en la SMO de Nuevo León es cohortal, es decir que predomina un sólo estrato (III).
CONCLUSIÓN
El matorral submontano en la Sierra Madre Oriental en el estado de Nuevo León con historial de extracción selectiva de sus componentes leñosos se encuentra conformado principalmente por Acacia rigidula, Bernardia myricaefolia y Acacia berlandieri, y las especies más importantes ecológicamente son Acacia rigidula, Cordia boissieri y Zanthoxylum fagara. Los elementos anteriores son claves para la identificación del matorral submontano en el estado de Nuevo León y que se caracteriza por presentar elementos de hasta 13 metros de altura. Se presenta como un ecosistema relativamente diverso y con gran riqueza de especies, lo cual es favorecido a su vez por la influencia de elementos de comunidades vegetales de bosque de encino y matorral xerófilo colindante.
La estructura vertical presenta un estrato principal, el estrato III, el cual se encuentra conformado por especies arbustivas de hasta 6 m de altura. Asimismo destacan en el estrato I elementos característicos de bosque de encino debido algunos sitios estudiados representan ecotonos con la comunidad de matorral submontano estudiada. La presente investigación aporta información cuantitativa de la composición horizontal y vertical de los elementos florísticos del matorral submontano de la SMO.
AGRADECIMIENTOS
Los autores agradecen la colaboración en campo de Raúl Pulido Pérez, Enrique Gómez Nacianceno, Raymundo Daniel Vega Aranda, José Cruz Ambriz Marín y Edgar Iván Aguirre Cuevas.
LITERATURA CITADA
ALANÍS, E.; JIMÉNEZ, J.; ESPINOZA, D.; JURADO, E.; AGUIRRE, O. A.; GONZÁLEZ, M. A. 2008. Evaluación del estrato arbóreo en un área restaurada post–incendio en el Parque Ecológico Chipinque, México. Revista Chapingo Serie Ciencias Forestales y del Ambiente 14(2): 81–87. [ Links ]
ALANÍS, E.; JIMÉNEZ, J.; AGUIRRE, O. A.; TREVIÑO, E.; JURADO, E.; GONZÁLEZ M. A. 2008. Efecto del uso del suelo en la fitodiversidad del matorral espinoso tamaulipeco. Revista Ciencia UANL 11(1): 56–62. [ Links ]
ALANÍS, G. J.; CANO, G.; ROVALO, M. 1996. Vegetación y Flora de Nuevo León. Una Guía Botánico–Ecológica. Impresora Monterrey, S. A. de C. V. México. 251 p. [ Links ]
ALANÍS, G. J.; GUZMÁN, M. A.; GONZÁLEZ, M.; CANO, G. 1995. Flora Representativa de Chipinque. Árboles y Arbustos 1era. Parte. Monterrey N. L., México. 40 p. [ Links ]
ARRIAGA, L. 2009. Implicaciones del cambio de uso de suelo en la biodiversidad de los matorrales xerófilos: un enfoque multiescalar. Investigación ambiental 1(1): 6–16. [ Links ]
CLIFFORD, H.; STEPHENSON, W. 1975. An Introduction to Numerical Classification. Academic Press. New York. 229 p. [ Links ]
CORRAL, J. J.; AGUIRRE, O. A.; JIMÉNEZ, J.; CORRAL, S. 2005. Un análisis del efecto del aprovechamiento forestal sobre la diversidad estructural en el bosque mesófilo de montaña <<El Cielo>>, Tamaulipas, México. Investigación Agraria: Sistemas y Recursos Forestales 14(2): 217–228. [ Links ]
CURTIS, J. T.; MCINTOSH, R. P. 1950. The interrelations of certain analytic and synthetic phytosociological characters. Ecology (31): 434–455. [ Links ]
ESTRADA, E.; VILLARREAL J.; JURADO, E. 2005. Leguminosas del norte del estado de Nuevo León, México. Acta Botanica Mexicana 73: 1–18. [ Links ]
ESTRADA, E.; YEN,A. D.; VILLARREAL, J. 2004. Leguminosas del centro del estado de Nuevo León, México. Anales del Instituto de Biología, Universidad Nacional Autónoma de México, Serie Botánica 75: 73–85. [ Links ]
FOROUGHBAKHCH, R.; ALVARADO, M. A.; NÚÑEZ, A.; HERNÁNDEZ, J.; ROCHA, A. 2003. Structural analysis and performance of Helietta parvifolia (Gray) Benth. in southeastern Nuevo Leon, Mexico. Interciencia 28(11): 651–655. [ Links ]
GARCÍA, J.; JURADO, E. 2008. Caracterización del matorral con condiciones prístinas en Linares N. L., México. Ra Ximhai 4(1): 1–21. [ Links ]
INEGI. 1986. Síntesis Geográfica de Nuevo León. Secretaría de Programación y Presupuesto. México, D. F. 170 p. [ Links ]
MAGURRAN, A. 2004. Measuring Biological Diversity. Blackwell Science Ldt. Blackwell Publishing Company. Oxford, UK. 256 p. [ Links ]
MITTERMEIER, R. A. y GOETSCH, C. 1997. Megadiversidad: Los países biológicamente más ricos del mundo. Cemex, Ciudad de México. [ Links ]
MUELLER–DOMBOIS, D.; ELLENBERG, H. 1974. Aims and Methods of Vegetation Ecology. John Wiley and Sons Ltd. New York. 547 p. [ Links ]
PALACIO, J. L.; BOCCO, G.; VELÁZQUEZ, A.; MAS, J. F.; TAKAKI, F.; VICTORIA, A.; LUNA, L.; GÓMEZ, G.; LÓPEZ, J.; PALMA, M.; TREJO, I.; PERALTA, A.; PRADO, J.; RODRÍGUEZ, A.; MAYORGA, R.; GONZÁLEZ, F. 2000. La condición actual de los recursos forestales en México: resultados del Inventario Forestal Nacional 2000. Investigaciones Geográficas, Boletín del Instituto de Geografía, UNAM (43): 183–203. [ Links ]
PRETZSCH, H. 1996. Strukturvielfalt als Ergerbnis Waldbaulichen Handels. Deutscher Verband Forstlicher Forschungsanstalten. Sekt. Ertragskunde. Jahrestagung, Nehresheim. pp. 134–154. [ Links ]
RZEDOWSKI, J. 2006. Vegetación de México. 1ª Edición digital, Comisión Nacional para el Conocimiento y Uso de la Biodiversidad. México. 504 p. [ Links ]
SECRETARÍA DE MEDIO AMBIENTE Y RECURSOS NATURALES. 2006. El Medio Ambiente en México 2005: en resumen. México. 91 p. [ Links ]
SHANNON, C. 1948. The Mathematical Theory of Communication. En Shannon, C.E.; Weaver, W. (Ed). Univ. of Illinois Press. pp. 134–154. [ Links ]
TREVIÑO, E. J.; CAVAZOS, C.; AGUIRRE, O. A. 2001. Distribución y estructura de los bosques de galería en dos ríos del centro sur de Nuevo León. Madera y Bosques 7(1): 13–25. [ Links ]

