










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Chapingo serie ciencias forestales y del ambiente
versión On-line ISSN 2007-4018versión impresa ISSN 2007-3828
Rev. Chapingo ser. cienc. for. ambient vol.15 no.1 Chapingo ene./jun. 2009
Plantas nodriza en la reforestación con Pinus hartwegii Lindl.
Nurse plants in the reforestation with Pinus hartwegii Lindl.
A. Ramírez–Contreras; D.A. Rodríguez–Trejo
División de Ciencias Forestales, Universidad Autónoma Chapingo. Km 38.5 carretera México–Texcoco. Chapingo, Estado de México. C. P. 56230. Correo–e: dantearturo@yahoo.com.
Recibido: 15 de febrero, 2008
Aceptado:6 de febrero, 2009
RESUMEN
En el Ajusco, Distrito Federal, fueron plantados Pinus hartwegii en cinco tratamientos de plantas nodriza: al noreste (NE) de Lupinus montanus, al NE de Penstemon gentianoides, entre Festuca tolucensis, entre F. tolucensis con chaponeo, y en pequeños claros naturales. Se evaluó supervivencia, diámetro, altura, biomasa y concentración de N, P y K a seis (temporada de sequía) y doce meses (lluvias) de establecida la plantación. Se midió la radiación fotosintéticamente activa (RFA) en los distintos tratamientos mediante cámara con lente hemisférica y programa Hemiview. La mortalidad fue analizada con regresión logística, las variables morfológicas y fisiológicas fueron evaluadas con análisis de varianza multivariado y con análisis de varianza. A un año de la plantación, no hubo diferencias en mortalidad entre tratamientos (promedio=15 %). Los brinzales fueron más altos en los tratamientos con Lupinus, Penstemon y chaponeo, que entre zacates y pequeños claros naturales (P<0.10). Hubo mayores concentraciones de N y K en el follaje de los brinzales junto a Lupinus al año de establecida la plantación, en la temporada de lluvias, que en el tratamiento de zacates y superaron al chaponeo en N (P<0.10). La RFA fue menor en los tratamientos con nodrizas que en brinzales, sin limitar el crecimiento de los últimos. Se recomienda el uso de Lupinus como nodriza de P. hartwegii.
Palabras clave: aporte de potasio, Lupinus montanus, Penstemon gentianoides, plantación, restauración.
ABSTRACT
On the Ajusco volcano, Mexico City, Mexico, Pinus hartwegii seedlings were planted in five treatments: NE side of Lupinus montanus plants, NE side of Penstemon gentianoides plants, between Festuca tolucensis grasses, in small openings where grasses were cleared away, and in small natural gaps among the grasses. Mortality, diameter, height, biomass and foliar nutrient concentration (N, P, K) were evaluated six (dry season) and 12 months (rainy season) after tree–planting. Photosintetically active radiation (PAR) was measured with hemispheric photographs and the program Hemiview. Mortality was analyzed using logistic regression, while morphological and physiological variables were analyzed with multivariate variance analysis. Also was employed analysis of variance. There were no differences in mortality (average of 15 %) among treatments. The ANOVA showed higher trees in the Lupinus, Penstemon and cleared treatments, in comparison to grass and small natural gaps treatments (P<0.10). The MANOVA exhibited higher tree leaf concentrations of N and K in the seedlings associated to Lupinus one year after planting date in the rainy season, in comparison to the grass treatment, and higher N concentrations than the clearing treatment (P<0.10). The PAR was lower when the seedlings were associated to plants, but did not limit the growth of seedlings. Is recommended the use of Lupinus as nurse plant associated to Pinus hartwegii.
Keywords: potassium supply, Lupinus montanus, Penstemon gentianoides, plantation, restoration.
INTRODUCCIÓN
Las estimaciones de deforestación en México van de 75,000 hasta casi 2,000,000 ha·año–1 (Lund et al., 2002). La FAO refiere 314,000 ha anuales para el país (Hoare, 2005). La reforestación es una parte de los esfuerzos que se han realizado para contribuir con la restauración en el Volcán Ajusco del Distrito Federal. La Universidad Autónoma Metropolitana (2003) señala una supervivencia promedio de 46.5 % para las plantaciones forestales del sur del D. F. Con la finalidad de aumentar tal supervivencia en las plantaciones que se llevan a cabo en las elevadas altitudes del lugar, donde prevalece Pinus hartwegii Lindl. a más de 3,500 m, es necesario estudiar diferentes técnicas que ayuden a dicho propósito, una de ellas puede ser la utilización de plantas nodriza.
Las plantas nodriza dan protección (o recursos) a plántulas de otras especies en un ambiente difícil, mientras estas últimas crecen lo suficiente para enfrentar las condiciones del medio (Cavieres et al., 2006). Bajo la sombra de una planta nodriza, incluyendo árboles, las temperaturas del aire y del suelo son menos extremas, y la humedad de las capas superficiales del suelo tiende a permanecer a menor profundidad (Callaway y Pugnaire, 1999).
Las plantas pueden ejercer fuertes efectos positivos directos o indirectos hacia otras especies. Por ejemplo, Callaway et al. (1996) encontraron que Pinus monophylla Torr. & Frém, fue ayudado por Artemisia tridentata Nutt.; no obstante, P. ponderosa Douglas ex. C. Lawson, una especie poco tolerante a la sequía, no pudo competir con A. tridentata (De Lucia et al. 1988).
Cuando algunas plantas son ayudadas por otras especies, su crecimiento es un indicador de los efectos de la especie nodriza y tal crecimiento puede deberse a una mayor concentración de nutrientes. El N forma parte de aminoácidos y proteínas, de la clorofila y participa en procesos enzimáticos (Barker y Bryson, 2006). Los árboles plantados junto a Lupinus arboreus Sims se benefician del N fijado de la atmósfera por esta especie, gracias a sus asociaciones simbióticas con la bacteria Rhizobium lupini. El mantenimiento de Lupinus implica una entrada continua de N en los ecosistemas forestales (Mattson, 2007). La descomposición de la materia orgánica y los brinzales de Lupinus exudan componentes de N que los árboles cercanos pueden usar para su desarrollo (Beets, 1996). No se hallaron antecedentes de Penstemon gentianoides como especie nodriza.
El P juega un papel muy importante en procesos como: generación de energía, síntesis de ácidos nucleicos, fotosíntesis, respiración, activación e inactivación de enzimas, metabolismo de carbohidratos y fijación de nitrógeno (Vance et al., 2003). Las raíces de las plantas pueden alterar la solución del suelo y hacer disponible o incrementar la disponibilidad de P inorgánico, por ejemplo mediante la acidificación de la rizósfera, la exudación de ácidos orgánicos y la secreción de fosfatos (Neumann y Römheld, 1999).
El K influye en la apertura y cierre estomatal, consecuentemente tiene que ver con la resistencia a ambientes secos y fríos (Mengel, 2006). Estos últimos como los del área de estudio. Las bajas temperaturas del suelo pueden afectar la adquisición de nutrientes, pues el agua estará viscosa, se afecta la membrana de las células de las raíces y se reduce la demanda de nutrientes en los brotes (Kozlowski y Pallardy, 1997, Lambers et al., 1998).
La radiación fotosintéticamente activa (RFA) se utiliza en el estudio de la ecofisología de las plantas, especialmente cuando se analizan procesos de intercepción de luz en fotosíntesis (Jones, 1992). Hay poca información relativa a potencial competencia por luz entre especies nodriza y las beneficiadas.
Considerando lo expresado en los párrafos anteriores, el objetivo de este trabajo fue determinar si Lupinus montanus H.B.K. y Penstemon gentianoides (Kunth) tienen influencia como plantas nodriza en la supervivencia, crecimiento y concentración foliar de N, P y K de Pinus hartwegii.
MATERIALES Y MÉTODOS
El estudio se realizó en el Parque Nacional Cumbres del Ajusco, a 19° 12' y 19° 12' 50" N y 99° 14' 35" y 99° 16' 20" O, al sur de la ciudad de México. El clima de la zona es C(W2)(w)(b')ig, que corresponde al clima templado subhúmedo con lluvias en verano, el más húmedo de los subhúmedos, con un porcentaje de lluvia invernal menor a 5% de la anual (García, 1981). Se identifican dos unidades de suelo que son: andosol y litosol. La vegetación que se desarrolla en el lugar está conformada por bosques de pino (Pinus hartwegii), asociado a pastizales de los géneros Festuca y Muhlenbergia y en el sitio abundan plantas como Lupinus montanus y Penstemon gentianoides, por haberse conducido ahí quemas prescritas hacia 2002.
Se estableció una plantación a 3,600 m, con 500 brinzales de Pinus hartwegii, proporcionados por la Comisión de Recursos Naturales y Desarrollo Rural (CORENADER) del gobierno de la ciudad de México, producidos en el vivero de San Luis Tlaxialtemalco. Estas plantas fueron producidas en contenedores V–93 (93 cm3 de capacidad) y tenían 1.5 años de edad al momento de ser plantadas.
Se utilizó un diseñó experimental completamente al azar, con cinco repeticiones y los siguientes tratamientos: a) plantación del brinzal al NE de Lupinus montanus, b) plantación al NE de Penstemon gentianoides, c) junto a zacates (Festuca) sin chaponeo, d) junto a zacates con chaponeo y e) en pequeños claros naturales (PCR). Los árboles fueron plantados a 20–30 cm de distancia de la base de las plantas potencialmente nodriza. La densidad media de plantación fue de 1,000 ha–1.
Las variables se midieron a los seis (marzo, primavera, época de sequía) y doce meses (septiembre, otoño, época de lluvias) de establecida la plantación. Para ello, fueron extraídos cinco brinzales de cada tratamiento. A esta muestra se le realizó la medición de diámetro (con vernier), altura (con regla), biomasa y concentración de nutrientes en el follaje. Para medir la biomasa total las muestras fueron secadas en un horno a 70 °C hasta obtener peso constante; el peso se determinó con una balanza electrónica digital. Para las determinaciones de nutrientes, se empleó el método micro Kjeldahl para el N (digestado con mezcla diácida, determinado por arrastre de vapor); el de fotocolorimetría para P total (digestado con mezcla diácida, determinando por fotocolorimetría y reducción con molibdo–vanadato); y el de espectrofotometría para el K total (digestado con mezcla diácida y determinada por espectrofotometría de emisión de flama). Las determinaciones de nutrientes fueron realizadas en el Laboratorio Central Universitario de la Universidad Autónoma Chapingo.
Para medir la radiación fotosintéticamente activa (RFA), se tomó una fotografía en cinco puntos de cada tratamiento en la plantación (donde estaban los mismos brinzales que fueron cosechados para medir nutrientes). Se utilizó una cámara digital, con una lente hemisférica y las imágenes digitales fueron analizadas con el programa HemiView V.2.1, utilizando el modelo simple molar, para obtener RFA en MJ m–2 año–1. Mediante revisión de todos los árboles plantados se determinó la supervivencia por tratamiento.
En el análisis de mortalidad se empleó regresión logística con el siguiente modelo (Hosmer y Lemeshow, 1998):
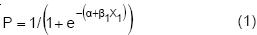
Donde: P=probabilidad de mortalidad, e=base de los logaritmos naturales, α=constante, β1= constante asociada a la variable independiente X1, X1=variable independiente.
Para determinar diferencias entre tratamientos se utilizó el análisis de varianza, con el modelo:

Donde: Yij =respuesta en la j–ésima repetición con el i–ésimo tratamiento; µ =media general; α =efecto del i–ésimo tratamiento del factor especie vecina; εij error experimental.
También se usó análisis de varianza multivariado. El modelo, con criterio de clasificación para p respuestas, fue:

Donde: yij = vector de respuestas en la j–ésima repetición con el i–ésimo tratamiento; µ = vector general de medias; Ti = vector de los efectos del i–ésimo tratamiento; εij = vector de errores aleatorios, que se suponen independientes, con distribución Np (0, Σ).
La comparación de medias entre tratamientos se hizo con la prueba de t (para el análisis de varianza) y con la prueba de T2 de Hottelling (Rencher, 2002) para el análisis multivariado.
A efecto de analizar los datos, se usaron el procedimiento logístico (Proc Logistic), el Procedimiento GLM (Proc GLM) para el análisis multivariado, así como el procedimiento ANOVA (Proc ANOVA) del programa SAS, v. 8, para microcomputadoras (SAS Institute, 1999).
RESULTADOS Y DISCUSIÓN
Mortalidad
El análisis de regresión logística no mostró diferencias significativas en la probabilidad de mortalidad de P hartwegii en los diferentes tratamientos (P>0.10). Las supervivencias promedio fueron: 85 % (tratamiento Penstemon), 82 % (Lupinus), 89 % (pequeños claros), 84 % (zacatón), 83 % (chaponeo). Myers (1992) menciona que los arbustos ayudan a abatir las condiciones desecantes y aumentan la supervivencia de Pinus palustris Mill. de regeneración natural en el sureste de Estados Unidos. En un estudio realizado con Quercus humilis Mill. en Francia, Rousset y Lepart (2000), encontraron mayor germinación de encinos bajo los arbustos Buxus sempervirens L. y Juniperus communis Lam., con respecto a las áreas abiertas, lo que sugiere que el mecanismo de facilitación está relacionado con los cambios en el microclima ocasionados por los arbustos. Helgerson (1990), menciona que la sombra de arbustos puede facilitar el establecimiento de plántulas durante las temporadas de sequía en el verano, por proveer mayor humedad o temperatura más fresca ocasionada por el efecto del micrositio. De igual manera, Castro et al. (2002) refieren mayor supervivencia de Pinus nigra J. F. Arnold, cuando se encontraba bajo el dosel del arbusto Salvia lavandulifolia Vahl. de una plantación realizada en España, mientras que en las áreas abiertas la supervivencia fue menor.
En nuestro caso las condiciones ambientales, como la sequía y bajas temperaturas en el año de estudio, no fueron tan extremas como para afectar la supervivencia en los tratamientos.
Diámetro, altura y biomasa total
Temporada de sequía (marzo, a 6 meses de la plantación)
El análisis de varianza multivariado no mostró efectos significativos entre los tratamientos para las variables diámetro, altura y biomasa total, en el periodo de sequía (P>0.10).
Temporada de lluvias (septiembre, a un año de la plantación)
El análisis de varianza multivariado para las variables analizadas, muestra que hay por lo menos dos tratamientos diferentes entre sí (Cuadro 1).

De acuerdo con la prueba T2– Hotelling los tratamientos que resultaron diferentes entre sí fueron el de chaponeo y el PCR (Figura 1). Con base en el ANOVA y la prueba de t, la altura de los pinos en los tratamientos Lupinus, chaponeo y Penstemon fue superior que la de aquellos en los tratamientos de pequeños claros naturales y de zacates (P<0.10), de forma semejante a lo referido por Baker et al. (1986), quienes señalan mayor crecimiento en árboles que fueron plantados cerca de plantas de Lupinus.

Ramírez y Rodríguez (2003), en una plantación realizada con Quercus rugosa en micrositios, también obtuvieron mayor incremento en altura, con respecto a los árboles que no contaban con alguna protección. De forma opuesta, Rousset y Lepart (2000) registraron diferencias estadísticamente significativas en la biomasa de Quercus humilis Mill., cuando se encuentra bajo el dosel de los arbustos (Buxus sempervirens L. y Juniperus communis L.), siendo mayor su biomasa cuando se le removió el dosel que le producía sombra. Las tendencias contrarias mencionadas, sin duda se relacionan con la tolerancia a la sombra: aquellas especies tolerantes pueden tener mayor crecimiento bajo sombra y las intolerantes mayor crecimiento cuando cuentan con más radiación solar. Pero también se relacionan con la reducción de extremos de temperatura y disponibilidad de agua y nutrientes. Así, Lenz y Facelli (2003) encontraron que cuando Orbea variegata Haw. se encuentra bajo la sombra de Atriplex vesicaria Heward ex Benth. tiende a aumentar su biomasa total.
N, P, K y radiación fotosintéticamente activa
Temporada de sequía (marzo)
El MANOVA mostró efectos significativos para las variables N, P, K y RFA (Cuadro 2). Con la prueba de T2– Hotelling los únicos tratamientos diferentes entre sí fueron los árboles plantados junto a Lupinus y el chaponeo (Figura 2), con una menor concentración foliar de N para el primer caso, lo cual puede deberse a competencia, pues Margolis y Brand (1990), sugieren que en sitios que se mantienen totalmente cubiertos por herbáceas, como los zacates, generalmente son pobres en nutrientes, y comúnmente los pinos que crecen junto a herbáceas tienden a competir por N (Robinson et al., 2002). La competencia por N frecuentemente ha sido citada como una razón por la cual las plantas tienden a crecer lentamente en algunas plantaciones de coníferas (Nambiar y Sands, 1993). No obstante, en nuestro caso, esta menor concentración foliar de N no fue crítica (1.4 %), fue temporal (ver siguiente subtítulo) y no afectó el crecimiento.


Temporada de lluvias (septiembre)
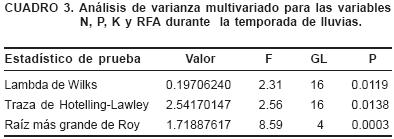
El MANOVA muestra que hay efectos significativos entre tratamientos para las variables N, P, K y RFA (Cuadro 3). En nuestro trabajo, los árboles junto a Lupinus mostraron la mayor concentración de N, K y P con respecto al tratamiento con zacates y más N que el tratamiento de chaponeo (Figura 3).

Se sabe que muchas leguminosas aumentan la cantidad de N que puede estar disponible para especies que se encuentran creciendo junto a ellas (Haines et al., 1978). De manera similar, se reporta mayor cantidad de N foliar en Juglansregia L., cuando crece junto a algunas leguminosas (Van Sambeek et al., 1986) y P. radiata D. Don. plantados junto a Lupinus arboreus (Baker et al., 1986).
En la evaluación realizada en septiembre, hubo mayor concentración de P en el follaje de los brinzales de Pinus hartwegii en el tratamiento de árboles plantados al NE de Lupinus montanus, con respecto al tratamiento zacates y es que acorde con Horst et al. (2001), los arbustos de Lupinus exudan quelatantes de malato y citrato, los cuales hacen más disponible el P para las plantas aledañas a estos arbustos. Además, Gardner y Boundy (1983) mencionan que en presencia de especies que presentan raíces proteoideas como las leguminosas, pueden movilizar fósforo y otros nutrientes hacia plantas de las especies que se encuentran cerca de éstas. Así mismo, las raíces proteoideas pueden llegar a inducir la formación de una capa de materia orgánica, generalmente rica en componentes de ácidos fosfáticos, fenólicos, agua, etc., que enriquecen el suelo (Watt y Evans, 1999). Los árboles plantados junto a Lupinus también mostraron mayores niveles de K que entre el zacatón por la menor competencia y la mayor disponibilidad del elemento a partir de la descomposición de partes muertas del Lupinus.
Por cuanto toca a la RFA, los tratamientos con arbustos exhibieron los niveles más bajos (Figura 3). El tratamiento PCR es el que presenta mayor RFA y menor concentración de nutrientes. En Fagus sylvatica L. y Pinus resinosa Sol. Ex Aiton la mayor eficiencia en el uso de nutrientes se ha encontrado con la mayor cantidad de luz disponible (Minotta y Pinzauti, 1996). De acuerdo con Franco y Nobel (1989), la sombra de las plantas nodriza disminuye la RFA disponible para las plántulas que se encuentran bajo sus copas, lo cual puede limitar la absorción de CO2, como ocurre con varias plantas del desierto. Chadzon (1988) refiere que una reducción del 20 % en la radiación solar directa puede afectar el crecimiento de algunas especies. Sin embargo, en nuestro trabajo la RFA bajo las plantas nodriza no era muy reducida, 9,000 MJ·m–2–año–1, en comparación con 10,500 MJ·m–2–año–1 en los claros pequeños, tratamiento que contó con los mayores niveles de radiación solar en el experimento.
La RFA para los brinzales plantados al NE de Penstemon fue menor que para los árboles plantados al NE de Lupinus, debido a que los primeros son de copa más densa que los segundos.
Los resultados del presente trabajo y considerando las variables estudiadas, dejan ver la conveniencia de utilizar a Lupinus montanus como nodriza de Pinus hartwegii en programas de reforestación. Tal uso podría ser mediante siembra previa de la primera especie o siembra o plantación al momento de plantar el pino o bien aprovechando la presencia de dicha especie en localidades incendiadas.
CONCLUSIONES
Supervivencia: Bajo las condiciones del presente estudio, no se hallaron efectos de los arbustos estudiados en la supervivencia de P. hartwegii.
Altura y biomasa: Gracias al chaponeo, los árboles exhibieron mayores alturas y biomasa que en claros naturales. Los árboles que crecieron asociados a arbustos o que recibieron chaponeo, tuvieron mayor altura que entre el zacatón y los pequeños claros naturales.
Concentración foliar de nutrientes: A seis meses de la plantación, durante la temporada de sequía, el tratamiento con chaponeo arrojó la mayor concentración de N foliar en los pinos, pero en la temporada de lluvias fue el tratamiento con Lupinus el que superó a los tratamientos zacatón y chaponeo en N foliar. El tratamiento Lupinus también superó al tratamiento zacatón en P y K.
Radiación fotosintéticamente activa: La RFA fue menor en los tratamientos con plantas nodriza, pero no tanto como para limitar el crecimiento de los pinos, intolerantes a la sombra.
El uso de arbustos nodriza como Lupinus para servir como nodriza a Pinus hartwegii plantados les otorga ventajas en crecimiento en altura y reservas de nutrientes (N, P, K). En el caso del K, les puede ayudar a tolerar mejor, durante los primeros años de plantados, las bajas temperaturas y limitaciones de humedad propios de las altitudes en las que se encuentra este pino.
AGRADECIMIENTOS
A la Comunidad de San Miguel y Santo Tomás Ajusco, por su autorización y apoyo para el establecimiento de este experimento. A la UACh, por los apoyos recibidos para la realización del presente trabajo, parte del Proyecto Ajusco de la UACh sobre ecología del fuego, manejo intergal del fuego y restauración de áreas afectadas por incendios forestales. Al Vivero San Luis Tlaxialtemalco y al Gobierno de la ciudad de México, por la donación de los árboles con los que se llevó a cabo la presente investigación.
LITERATURA CITADA
BAKER, T. G.; OLIVER, G. R.; HODGKISS, P. D. 1986. Distribution and cycling of nutrients in Pinus radiata as affected past lupin growth and fertilizer. Forest Ecology and Management 17: 169–187. [ Links ]
BARKER, A. V.; BRYSON, G. M. 2006. Nitrogen. In: BARKER, A. V.; PILBEAM, D. J. Handbook of plant nutrition. Taylor and Francis. U.S.A. pp. 21–50. [ Links ]
BEETS, P. N. 1996. Carbon partitioning in Pinus radiata stands in relation to foliage nitrogen status. Tree physiology 16(1–2): 131–138. [ Links ]
CASTRO, J.; ZAMORA, R.; HÓDAR, J. A.; GÓMEZ, J. M. 2002. Use of shrubs as nurse plants: a new technique for reforestation in mediterranean mountains. Restoration Ecology 10: 297–305. [ Links ]
CALLAWAY, R. M.; PUGNAIRE, F. I. 1999. Facilitation in plant communities. In: PUGNAIRE, F. I.; VALLADARES, F. (eds.). Handbook of functional plant ecology. Marcel Dekker. New York. pp. 623–649. [ Links ]
CALLAWAY, R. M.; DE LUCIA, E. H.; MOORE, D.; NOWAK, R.; SCHLESINGER, W. H. 1996. Competition and facilitation: contrasting effects of Artemisia tridentata on Pinus ponderosa and P. monophylla. Ecology 77: 2130–2141. [ Links ]
CAVIERES, L. A.: BADANO, E. I., SIERRA A., A.; GÓMEZ G., S.; MOLINA M., M. A. 2006. Positive interactions between alpine plant species and the nurse cushion plant Laretia acaulis do not increase with elevation in the Andes of central Chile. New Phytologist 169: 59–69. [ Links ]
CHADZON, R. L. 1988. Sunflecks and their importance to forest understory plants. Advances in Ecological Research 18: 1–63. [ Links ]
DE LUCIA, E. H.; W. H. SCHLESINGER; W. D. BILLINGS. 1988. Water relations and the maintenance of Sierran conifers on hydrothermally altered rock. Ecology 69 :303–311. [ Links ]
FRANCO, A. C.; NOBEL, P. S. 1989. Effect of nurse plants on the microhabitat and growth of cacti. Journal of Ecology 77: 870–886. [ Links ]
GARCÍA DE M., E. 1981. Modificaciones al sistema de clasificación climática de Kõeppen (para adaptarlo a las condiciones de la República Mexicana). Instituto de geografía, UNAM. México, D. F. 252 p. [ Links ]
GARDNER, W. K.; BOUNDY K., A. 1983. The acquisition of Phosphorus by Lupinus albus L. IV. The efecct of the interplanting wheat and white lupin on the growth and mineral composition of the two species. Plant and Soil 70: 391–402. [ Links ]
HAINES, S. G.; HAINES, L. W.; WHITE, G. 1978. Leguminous plant increase sycamore growth in Northern Alabama. Soil Science Society of America Journal 42: 130–132. [ Links ]
HELGERSON, O. T. 1990. Heat damage in tree seedlings and its prevention. New Forests 3: 333–358. [ Links ]
HOARE, A. 2005. Irrational numbers: Why the FAO's forest assessments are misleading. Rainforest Foundation. U.S.A. 23 p. [ Links ]
HORST, W. J.; KAMH, M.; JIBRIN, J. M.; CHUDE, V. A. 2001. Agronomic measures for increasing P availability to crops. Plant and Soil 237: 211–233. [ Links ]
HOSMER, J. R.; LEMESHOW, S. 1998. Applied logistic regression. Wiley and Sons. U.S.A. 307 p. [ Links ]
JONES, G. H. 1992. Plants and microclimate. A quantitative approach to environmental plant physiology. 2nd. ed. Cambridge University Press. Cambridge, Great Britain. 428 p. [ Links ]
KOZLOWSKI, T. T.; PALLARDY, S. 1997. Physiology of woody plants. 2nd ed. Academic Press, San Diego. 411 p. [ Links ]
LAMBERS, H.; CHAPIN III, F. S.; PONS, T. L. 1998. Physiological plant ecology. Springer. New York, USA. 540 p. [ Links ]
LENZ, I. T.; FACELLI, J. M. 2003. Shade facilitates an invasive stem succulent in a chenopod shrubland in South Australia. Austral Ecology 28: 480–490. [ Links ]
LUND, H. G.; TORRES, V.; TURNER, A.; WOOD, L. 2002. México. Análisis crítico de los estimados disponibles de deforestación. USAID– SEMARNAT. [ Links ]
MARGOLIS, H. A.; BRAND, D. G. 1990. An ecophysiological basis for understanding plantation establishment. Canadian Journal of Forestry Research 20: 375–390. [ Links ]
MATTSON, S. 2007. Pinus contorta growth in boreal Sweden as affected by combined lupin treatment and soil scarification. Silva Fennica 41(4): 649–659. [ Links ]
MENGEL, K. 2006. Potassium. In: BARKER, A. V.; PILBEAM, D. J. Handbook of plant nutrition. Taylor and Francis. U.S.A. pp. 91–120. [ Links ]
MINOTTA, G.; PINZAUTI, S. 1996. Effects of light and soil fertility on growth, leaf chlorophyll content and nutrient use efficiency of beech (Fagus sylvatica L.). Forest Ecology and Management 86: 61–71. [ Links ]
MYERS, R. L. 1992. Shrub and high pine. In: MYERS R. L.; EweI, J.J. (eds.). Ecosystems of Florida. University of Central Florida Press. Orlando, Florida. pp. 150–193. [ Links ]
NAMBIAR, E. K. S.; SANDS R. 1993. Competing for water and nutrients in forest. Canadian Journal of Forestry Research 23: 1955–1968. [ Links ]
NEUMANN, G.; RÖMHELD V. 1999. Root excretion of carboxylic acids and protons in phosphorus–deficient plants. Plant and Soil 211: 121–130. [ Links ]
RAMÍREZ C., A.; RODRÍGUEZ T., D. A. 2004. Efecto de calidad de planta, exposición y micrositio en una plantación de Quercus rugosa. Revista Chapingo. Serie Ciencias Forestales y del Ambiente 1: 5–11. [ Links ]
RENCHER, C. A. 2002. Methods of multivariate analysis. 2nd. Ed. Wiley. New York. 708 p. [ Links ]
ROBINSON, D. E.; WAGNER, R. G.; SWANTON, C. J. 2002. Effects of nitrogen on the growth of jack pine competing with Canada blue–joint grass and large–leaved aster. Forest Ecology and Management 160: 233–242. [ Links ]
ROUSSET, O.; LEPART, J. 2000. Positive and negative interactions at different life stages of a colonizing species (Quercus humilis). Journal of Ecology 88: 401–416. [ Links ]
SAS INSTITUTE. 1999. Statistical Analysis System. V. 8.0. Cary, North Carolina, U. S. A. [ Links ]
UNIVERSIDAD AUTÓNOMA METROPOLITANA. 2003. Evaluación externa del Programa Nacional de Reforestación en el Distrito Federal. Universidad Autónoma Metropolitana. 68 p. URL: http://www.conafor.gob.mx/programas_nacionales_forestales/evaluaciones/documentos_evaluacion.htm (fecha de consulta: 6 de mayo de 2009). [ Links ]
VANCE, C. P.; UHDE–STONE, C.;ALLAN, D. L. 2003. Phosphorus acquisition and use: critical adaptations by plants for securing a nonrenewable resource. New Phytologist 157: 423–447. [ Links ]
VAN SAMBEEK, J. W.; PONDER, F.; RIETVELD, W. J. 1986. Legumes increase growth and alter foliar nutrient levels of black walnut saplings. Forest Ecology and Management. 17: 159–167. [ Links ]
WATT, M.; EVANS, J. R. 1999. Proteoid Roots. Physiology and Development. Plant Physiology 121: 317–323. [ Links ]

