











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En la actualidad, las actividades antropogénicas han provocado una serie de problemas ambientales que ponen en riesgo el desarrollo de la agricultura. Uno de los principales problemas es la contaminación del agua y suelo con metales pesados, que por sus características (toxicidad, biodisponibilidad, bioaccesibilidad, persistencia y alta solubilidad) ocasionan diversos problemas en los organismos vivos (Li et al., 2015; Ruíz-Huerta et al., 2017; Thapa et al., 2012; Xian et al., 2015). El arsénico (As) es uno de los principales contaminantes del mundo (Sarkar & Paul, 2016). Es un metaloide tóxico que se libera al ambiente como resultado de procesos tanto naturales como antropogénicos (Kalita et al., 2018). Este metaloide tiene un impacto negativo en plantas, animales y humanos (Zvobgo et al., 2019). En las plantas, la mayor parte del As se retiene en las células de la raíz y, aunque la translocación a los brotes es relativamente baja, varía sustancialmente entre especies e incluso dentro de la misma especie (Finnegan & Chen, 2012). El arsénico induce alteraciones nutricionales en las plantas al afectar la absorción de nutrientes por competencia directa con otros nutrientes, esto a su vez altera los procesos metabólicos (Gomes et al., 2014). En función de su estado de oxidación, existen dos especies inorgánicas de As, arseniato pentavalente completamente oxidado (As V) y arsenito trivalente reducido (As III) (Sánchez-Pardo et al., 2015; Xu et al., 2015). El arsénico (V) actúa como un análogo de fosfato debido a la similitud química entre ambos, ingresa a la célula mediante transportadores de fosfato, afectando la absorción de este elemento (Panda et al., 2010; Tripathi P. et al., 2013). En el caso del As (III), los transportadores que se han reportado en el cultivo del arroz son los de silicio y algunas acuaporinas relacionadas con el silicio (Chen et al., 2017). El As ingresa a las células a través de sistemas de absorción de nutrientes como permeasas de fosfato (arseniato) y/o acuagliceroporinas (Garbinski et al., 2019). Uno de los principales problemas del As es que puede reemplazar al fósforo, pero no puede realizar sus funciones biológicas (Gunes et al., 2009). Esto desencadena una serie de problemas como la absorción reducida de otros nutrientes, alteraciones en el metabolismo de los carbohidratos, reducción de la tasa fotosintética e incluso muerte celular (Gomes et al., 2012; Panda et al., 2010; Stoeva et al., 2005; Zvobgo et al., 2014).
El silicio (Si) es un elemento con múltiples beneficios, ya que puede mejorar las condiciones del suelo y la absorción de nutrientes en las plantas (Zargar et al., 2019). El silicio puede aumentar la absorción de Ca y K en las hojas de maíz bajo estrés abiótico (Kaya et al., 2006). Incluso concentraciones bajas de silicio pueden mejorar la absorción de K tanto en hidroponía como en suelo, debido a la activación de la H+ATPasa (Mali & Aery, 2008). El mecanismo de absorción de silicio varía entre diferentes especies de plantas y aparentemente depende de la presencia de transportadores específicos de este elemento (Kaur & Greger, 2019). La tolerancia inducida por el silicio al estrés por metales pesados se produce a través de mecanismos que incluyen la disminución de la concentración de iones metálicos en el sustrato o suelo, coprecipitación de metales tóxicos, quelación, estimulación de antioxidantes y alteraciones estructurales en plantas (Bhat et al., 2019).
El uso de la nanotecnología en cultivos agrícolas y la protección del ambiente ha crecido en los últimos años debido a las propiedades físico-químicas exclusivas que poseen los nanomateriales (Cui et al., 2017). Las nanopartículas de silicio (Si NPs) pueden reducir el efecto tóxico del As en las plantas (Tripathi D. K. et al., 2015). Por ejemplo, la aplicación de Si NPs aumenta la fotosíntesis de la planta en presencia de cadmio, facilitando el transporte de nutrientes a través del xilema (Gao et al., 2018). También se ha informado que aumentan la eficiencia en el uso de agua y nutrientes (Alsaeedi et al., 2019). En cuanto al tomate (Solanum lycopersicum L.), es una hortaliza muy importante, que se produce en invernadero y campo abierto en México. Sin embargo, las concentraciones más altas de As aparecen en los acuíferos aluviales de las zonas áridas y semiáridas de México (Alarcón-Herrera et al., 2020), que generalmente se utilizan para agua de riego. Por lo tanto, el objetivo de este estudio fue determinar el efecto de la aplicación de SiO2 NPs en la absorción de macro y micronutrientes en plantas de tomate regadas con agua contaminada con As (V).
Material y Métodos
Crecimiento del Cultivo
El experimento se llevó a cabo en un invernadero de policarbonato con control automático de temperatura en la Universidad Autónoma Agraria Antonio Narro, Coahuila, México. Como material vegetal se utilizaron semillas de tomate (Solanum lycopersicum L.) híbrido “Sun 7705”, tipo saladete y crecimiento indeterminado. Las plántulas se trasplantaron a bolsas de polietileno negro de 12 L de capacidad que contenían una mezcla de peat moss y perlita (1:1) como sustrato de crecimiento. El cultivo se manejó a un solo tallo y se desarrolló durante 150 días después del trasplante.
Diseño Experimental y Nutrición de las Plantas
Se estableció un experimento con un diseño completamente al azar y arreglo factorial con dos factores: un factor fue la concentración de arsénico (seis niveles) y otro factor fue la concentración de SiO2 NPs (tres niveles), siendo un total de 18 tratamientos. Se añadió arseniato (As V) utilizando como fuente Na2HAsO4 * 7H2O, se mezcló con el agua de riego siempre que se preparó la solución nutritiva para obtener las concentraciones finales de 0, 0.2, 0.4, 0.8, 1.6 y 3.2 mg L-1 de As (V). Estas concentraciones se definieron con base a la Norma Oficial Mexicana NOM-001-ECOL-1996, que establece que los límites máximos permisibles (MPL) de arsénico en agua para uso agrícola son 0.2 mg L-1 promedio mensual y 0.4 mg L-1 promedio diario (DOF, 1997). Las SiO2 NPs se aplicaron vía suelo desde el trasplante, cada tres semanas haciendo un total de seis aplicaciones (10 ml por aplicación a cada planta), las concentraciones aplicadas fueron 0, 250 y 1000 mg L-1 de SiO2 NPs. La morfología de las SiO2 NPs es esférica, con un tamaño de 10-20 nm, un área superficial de 160 m2 g-1 y una densidad aparente de 0.08-0.1 g cm-3 (SkySpring Nanomaterials Inc., USA).
Siempre se suministró arsénico en el agua de riego para simular el agua contaminada de acuerdo a la concentración máxima permisible de As en el agua de riego en México (0.2 mg L-1) (DOF, 1997). La selección de las dosis de SiO2 NPs se llevó a cabo a partir de estudios publicados donde se describe una mejor comprensión de cómo las SiO2 NPs inhiben la absorción y transporte de As, y pueden proporcionar una nueva estrategia para diseñar mecanismos efectivos para disminuir la acumulación en las plantas (Cui et al., 2019). Además, las concentraciones de NPs fueron elegidas de un estudio previo con aplicación de SiO2 NPs en concentraciones altas (100, 500 y 2000 mg L-1) en algodón (Le et al., 2014).
La nutrición de las plantas se manejó a través de un sistema de riego dirigido usando la solución Steiner (Steiner, 1961). Esta se aplicó a diferentes concentraciones según la etapa fenológica: 25 % en etapa vegetativa, 50 % en floración, 75 % en cuajado de frutos y 100 % en llenado y cosecha de frutos. El pH de la solución nutritiva utilizada se mantuvo entre 6.0 y 6.5.
Muestreo de plantas
A los 150 días después del trasplante, se recolectaron las plantas y se tomaron muestras de hojas y raíces. Las muestras se secaron en un horno de secado a una temperatura constante de 80 °C durante 72 horas hasta que se alcanzó un peso constante. El material seco se molió hasta obtener un polvo para el análisis mineral. También se determinó la biomasa de raíces y brotes secos.
Análisis de Nutrientes
La concentración de macro y micro nutrientes en raíces y hojas se determinó mediante espectroscopia de fluorescencia de rayos X (XRF) con un instrumento ThermoScientific Niton FXL (límite de detección 2 ppm). Se analizaron 3 muestras y se evaluó la calidad de los resultados por triplicado para el control de calidad y para asegurar la confiabilidad de los datos analíticos, utilizando el material de referencia NIST 1573a para hojas de tomate.
Obteniendo 90 ± 10 % de recuperación.
Análisis Estadísticos
Se realizó un análisis de varianza (ANOVA) para determinar las diferencias entre tratamientos, y una prueba de medias de acuerdo a la Diferencia Mínima Significativa de Fisher (α = 0.05). Para realizar los análisis se usó el paquete estadístico InfoStat (v2019). Además, se realizó un análisis de regresión para evaluar la relación entre las variables, para esto se usaron los valores medios de cada factor (As (V) o SiO2 NPs), y no se consideraron las interacciones entre factores. Para este procedimiento se usó el software SigmaPlot (V12.0).
Resultados
Producción de Biomasa
La producción de biomasa se vio afectada tanto por las diferentes concentraciones de As (V) como por la aplicación de SiO2 NPs (Figura 1). La biomasa seca de plantas expuestas a bajas concentraciones de As (V) (0.2-0.4 mg L-1) se incrementó ligeramente en comparación con los controles. Las concentraciones de As (V) de 0.8-1.6 mg L-1 disminuyeron el peso seco de la raíz (RDW), sin embargo, la concentración de 3.2 mg L-1 indujo 3.3 % más biomasa (Figura 1A). En el peso seco de la parte aérea (SDW), la tendencia muestra un aumento en la biomasa a medida que aumenta la concentración de As (V) (Figura 1C). La aplicación de SiO2 NPs presentó una tendencia similar tanto en RDW como en SDW, aunque no en la misma magnitud, en ambos casos la biomasa disminuyó; El RDW disminuyó hasta en un 49 % (Figura 1B) y el SDW disminuyó en un 18 % (Figura 1D), con 1000 mg L-1 de SiO2 NPs en comparación con el control.

Figura 1 Relación entre la producción de biomasa seca [raíz (A, B) y parte aérea (C, D)] de S. lycopersicum con los factores individuales de concentración de As (V) y concentración de SiO2 NPs. La curva de regresión de As (V) se refiere a los valores medios de tres repeticiones. La curva de regresión de las SiO2 NPs se refiere a los valores medios de seis repeticiones.
La interacción de As (V) con las SiO2 NPs tuvo un efecto negativo en la producción de biomasa, ya que el RDW disminuyó considerablemente, llegando a 61.9 % y 63.4 % con la concentración de 0.8-1.6 mg L-1 de As (V) más 1000 mg L-1 de SiO2 NPs (Figura 2A). El efecto de la interacción As (V)-SiO2 NPs en la producción de SDW mostró la misma tendencia decreciente, aunque en menor proporción que el RDW. La concentración de 3.2 mg L-1 de As (V) más la aplicación de 250 mg L-1 de SiO2 NPs disminuyó el SDW en 3.1 %. Sin embargo, esta misma concentración de As (V) junto con 1000 mg L-1 de SiO2 NPs aumentó en 6.89% la producción de SDW en relación al control (Figura 2B).

Figura 2 Efecto de la interacción de los factores concentración de As (V) y concentración de SiO2 NPs sobre la producción de biomasa de raíces y brotes en plantas de S. lycopersicum. N= 6 ± error estándar. *indica diferencias significativas entre tratamientos de acuerdo con la prueba de Diferencia Mínima Significativa de Fisher (α = 0.05).
Efecto del Arseniato y las SiO2 NPs sobre la Absorción de Micronutrientes en Raíz y Hojas
La concentración de As (V) en la solución nutritiva afectó la concentración de Fe y Cu en la raíz, ya que a medida que aumentó la concentración de As (V), disminuyó la concentración de ambos micronutrientes (Figura 3A y 3C). En la concentración de Zn se observó un comportamiento diferente, las bajas concentraciones de As (V) (0.4-0.8 mg L-1) indujeron una mayor concentración de este micronutriente en 4.9 % y 15.5 % respectivamente, mientras que con la concentración más alta (3.2 mg L-1) disminuyó la concentración de Zn en la raíz (12.6 %) (Figura 3E). Un efecto muy similar se observó en la concentración de Fe, Cu y Zn en las hojas, con altas dosis de As (V) (1.6 y 3.2 mg L-1) disminuyó la concentración de Fe (9.7 % y 30.6 %), Cu (27.2 % y 41.8 %) y el Zn (23.5 % y 22.4 %) (Figura 3E).

Figura 3 Relación entre la concentración de Fe (A, B), Cu (C, D) y Zn (E, F) con los factores individuales concentración de As (V) y concentración de SiO2 NPs en raíces y hojas de S. lycopersicum. La curva de regresión de As (V) se refiere a los valores medios de tres repeticiones. La curva de regresión de las SiO2 NPs se refiere a los valores medios de seis repeticiones.
Las SiO2 NPs modificaron la concentración de Fe. En la raíz se observó una disminución de 14.4 % y 8.7 % de la concentración de este micronutriente debido a la exposición a 250 y 1000 mg L-1 respectivamente. En contraste, el efecto contrario se presentó en las hojas, ya que la concentración de Fe aumentó 45.7 % y 27.2 % con la aplicación de 250 y 1000 mg L-1 de SiO2 NPs respectivamente (Figura 3B). La aplicación de 250 y 1000 mg L-1 de SiO2 NPs indujo un aumento de Cu (10.8 % y 14.8 %) y Zn (18.5 % y 22.5 %) en la raíz; en las hojas no se observó ningún efecto (Figura 3D y 3F).
La interacción de As (V) con las SiO2 NPs presentó un efecto negativo ya que la concentración de Fe en la raíz disminuyó un 45.7 % con 3.2 mg L-1 de As y 1000 mg L-1 de SiO2 NPs (Figura 4A). Sin embargo, en ausencia de As (V) se observó que ambas concentraciones de SiO2 NPs (250 y 1000 mg L-1) indujeron una mayor concentración de Fe (8.3 % y 14.1 % más que el control) en las hojas. Cuando la concentración de As (V) fue de 0.2-1.6 mg L-1, la aplicación de 250 mg L-1 de SiO2 NPs indujo un aumento del contenido de Fe en las hojas de 13.1 %, 49.6 %, 37.9 % y 20.9 % respectivamente, en relación al control (Figura 4B).

Figura 4 Efecto de la interacción de los factores concentración de As (V) y concentración de SiO2 NPs sobre la concentración de Fe, Cu y Zn en raíz y hojas de S. lycopersicum. N= 6 ± error estándar. * indica diferencias significativas según la prueba de Diferencia Mínima Significativa de Fisher (α = 0.05).
La concentración de Cu en la raíz se vio afectada positivamente por la aplicación de SiO2 NPs en ausencia de As (V), presentando un aumento de 9.7 % y 25.2 % con 250 y 1000 mg L-1 de SiO2 NPs respectivamente respecto al control. La presencia de As (V) en general induce un efecto negativo sobre la concentración de Cu en la raíz. Sin embargo, se observó un efecto positivo cuando se aplicaron SiO2 NPs con respecto a las dosis de As (V) (0.8 y 1.6 mg L-1); pero en relación al control la dosis de 3.2 mg L-1 en combinación con 250 y 1000 mg L-1 de SiO2 NPs redujo en 19.1 % y 32.4 % respectivamente la concentración de Cu (Figura 4C). La concentración de Cu en las hojas aumentó 154.6 % con la combinación de 0.4 mg L-1 de As (V) y 250 mg L-1 de SiO2 NPs, en el resto de las interacciones As (V)-SiO2 NPs el efecto fue nulo o negativo (Figura 4D).
La concentración de zinc en la raíz aumentó por la aplicación de SiO2 NPs hasta en 104.5 % en relación al control y en ausencia de As (V), mientras que en presencia de As (V) se observaron incrementos de hasta 58 % aproximadamente (Figura 4E). En las hojas, el Zn aumentó con la presencia de 0.4 mg L-1 de As y en ausencia de SiO2 NPs, sin embargo, en combinación con SiO2 NPs se observó una menor concentración (Figura 4F).
La concentración de zinc aumentó 45.75 % con la concentración de 0.4 mg L-1 de As (V) con 250 mg L-1 SiO2 NPs. Sin embargo, también la combinación de 0.2 mg L-1 As (V) con 1000 mg L-1 SiO2 NPs indujo un incremento del 55 % en relación al control (Figura 4F).
Efecto del Arseniato y las SiO2 NPs sobre la Absorción de Macronutrientes en Raíz y Hojas
La concentración de K tanto en la raíz como en las hojas disminuyó a medida que aumentó la concentración de As (V) en la solución nutritiva; las disminuciones fueron de hasta 17.8 % en raíz y 22.5 % en hojas con la dosis de 3.2 mg L-1 de As (V) (Figura 5A). Sin embargo, la aplicación de 250 y 1000 mg L-1 de SiO2 NPs indujo el efecto contrario, ya que la concentración de K se incrementó en 46.1 % y 68.2 % en raíz, y 23.5 % y 33.3 % en hojas respectivamente (Figura 5B).

Figura 5 Relación entre la concentración de K (A, B), S (C, D) y P (E, F) con los factores individuales concentración de As (V) y concentración de SiO2 NPs en raíces y hojas de S. lycopersicum. La curva de regresión de As (V) se refiere a los valores medios de tres repeticiones. La curva de regresión de las SiO2 NPs se refiere a los valores medios de seis repeticiones.
La concentración de azufre en la raíz no se vio afectada por las diferentes concentraciones de As (V), ni por la aplicación de SiO2 NPs (Figura 5C y 5D). Sin embargo, la concentración de azufre en la raíz aumentó 10 % con la concentración más alta de As (V) (3.2 mg L-1) y 10.5 % con la concentración de 0.8 mg L-1. La concentración más baja de As (V) (0.2 mg L-1) indujo una disminución del 14.7 % en la concentración de azufre en las hojas (Figura 5C). Por otro lado, la aplicación de 1000 mg L-1 de SiO2 NPs incrementó la concentración de azufre en las hojas en un 6.7 % (Figura 5D).
La concentración de P se vio afectada por el As (V), sin embargo, el efecto fue diferente entre los órganos de la planta de tomate. En las hojas, la concentración de P disminuyó 41.4 %, 40.4 %, 11.8 %, 54.7 % y 31.1 % con las diferentes concentraciones de As (V) (0.2-3.2 mg L-1 respectivamente). En la raíz, la concentración de P aumentó 24.6 % con el As (V) a una concentración de 1.6 mg L-1 (Figura 5E). En el caso de la aplicación de 250 y 1000 mg L-1 de SiO2 NPs se indujo un aumento de 26.9 % y 54.9 % en la concentración de P en hojas, y 40.3 % y 36.2 % en raíces, respectivamente (Figura 5F).
La combinación de As (V) con las SiO2 NPs indujo cambios en la concentración de potasio tanto en la raíz como en las hojas (Figura 6). En ambos órganos cuando sólo se aplicaron SiO2 NPs se observó un aumento significativo en la concentración de potasio, siendo 70 % en raíces y 65.2 % en hojas (Figura 6A y 6B). Cuando se aplicaron SiO2 NPs en combinación con As (V), generalmente se observó un aumento de potasio en comparación con la aplicación de solo As (V), siendo el efecto mayor con la concentración de 0.2 mg L-1 de As (V) y 1000 mg L-1 de SiO2 NPs con un incremento de 106.7 % en raíz.
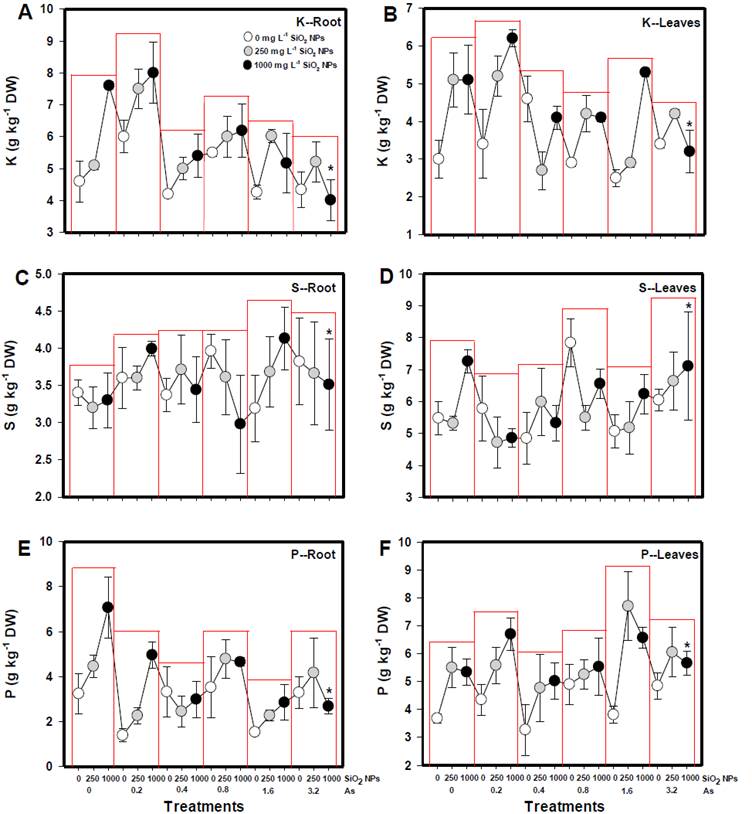
Figura 6 Efecto de la interacción de los factores concentración de As (V) y concentración de SiO2 NPs sobre la concentración de K, S y P en raíz y hojas de S. lycopersicum. N= 6 ± error estándar. * indica diferencias significativas según la prueba de Diferencia Mínima Significativa de Fisher (α = 0.05).
La concentración de azufre en las hojas aumentó 32.5 % con la aplicación de 1000 mg L-1 de SiO2 NPs en ausencia de As (V). Sin embargo, la concentración de 0.8 mg L-1 de As (V) sin la aplicación de SiO2 NPs también aumentó el contenido de este elemento en un 43.1 % (Figura 6C). En las hojas, la concentración de 0.8 mg L-1 de As (V) sin aplicación de SiO2 NPs aumentó el contenido de azufre (44.4 %). Además, la combinación de 1000 mg L-1 de SiO2 NPs con 1.6 y 3.2 mg L-1 de As (V) aumentó la concentración de azufre en 15.6 % y 31.7 % respectivamente (Figura 6D).
En ausencia de As (V), la concentración de fósforo aumentó significativamente en raíz (37.7 % y 117.6 %) y hojas (45.5 % y 45.1 %) con la aplicación de 250 y 1000 mg L-1 de SiO2 NPs (Figura 6E y 6F). Con la presencia de 1.6 mg L-1 de As (V) y con la aplicación de 1000 mg L-1 de SiO2 NPs, se observó un aumento de hasta 78.5 % en la concentración de fósforo en hojas (Figura 6F). En la raíz, la interacción de 1.6 y 3.2 mg L-1 de As (V) con 1000 mg L-1 de SiO2 NPs disminuyó la concentración de fósforo en 12.1 % y 17.3 % respectivamente (Figura 6E).
Discusión
Producción de Materia Seca
El problema de As en México es preocupante, se han detectado altos niveles de As en el agua potable en ciertas localidades de Coahuila (hasta 435 μg L-1) y Sonora (hasta 1,004 μg L-1); en aguas superficiales continentales en Puebla (hasta 780 μg L-1) y Matehuala, SLP (hasta 8,684 μg L-1); en aguas subterráneas en SLP (hasta 16,000 μg L-1) y Morelia, Michoacán (hasta 1,506 μg L-1); en suelos en Matehuala, SLP (hasta 27,945 μg g-1) y el área minera Xichú, Guanajuato (hasta 62,302 μg g-1); y en sedimentos de Zimapán, Hidalgo (hasta 11,810 μg g -1) y Matehuala, SLP (hasta 28,600 μg g-1) (Osuna-Martinez et al., 2021). Se ha informado que los fertilizantes que contienen macronutrientes también pueden aportar trazas de metales pesados (HM) y contaminantes potenciales no degradables y no destruibles, como el Cd y el Pb, que no tienen ningún propósito biológico establecido en las plantas, pero se reconoce que causan disfunciones fisiológicas, morfológicas y bioquímicas en las plantas (Tchounwou et al., 2012). Además, Salem et al. (2020) informaron que la aplicación de fertilizantes de fosfato y urea en suelos agrícolas durante cuarenta años de fertilización, proporciona concentraciones considerables de Cr, Cd y Ni, y que las concentraciones de los metales pesados en el suelo varían significativamente con las estaciones (invierno, primavera, verano y otoño). Sin embargo, la aplicación de fertilizantes agrícolas no aumenta significativamente el contenido total de metales pesados en el suelo por encima de los niveles de fondo durante varios años (Rutkowska et al., 2009). Los niveles altos de As en México están relacionados principalmente con el origen geogénico y minero.
El arsénico puede reemplazar al fosfato en los procesos respiratorios, interrumpiendo el metabolismo celular, generando difosfato-arseniato de adenosina (ADP-As) en lugar de trifosfato de adenosina (ATP) (Meharg, 1994). Dentro del tejido vegetal, el As (V) se reduce a As (III) por la enzima arseniato reductasa que se une a los grupos tiol de enzimas y proteínas que conducen a la inhibición de las funciones celulares (Finnegan & Chen, 2012). La exposición de las plantas a As produce inhibición del crecimiento, detiene la acumulación de biomasa y provoca trastornos fisiológicos en las plantas, debido a la alta toxicidad de este metaloide (Garg & Singla, 2011; Stoeva et al., 2005). Se ha reportado una reducción del 50 % en la producción de biomasa seca en plantas de arroz cultivadas en solución nutritiva con 4.0 mg L-1 de As (III) (Wang et al., 2010). Las plantas de girasol cultivadas en suelo contaminado con As (V) redujeron el peso seco de las raíces hasta en un 60.5 %, y el peso seco de la parte aérea hasta en un 49.2 %, cuando se expusieron a concentraciones de 40 y 80 mg kg-1 de As (V) respectivamente (Azeem et al. al., 2017). El presente estudio encontró que con la aplicación de diferentes concentraciones de As a niveles bajos hay una leve estimulación de la biomasa, mientras que a niveles altos no hubo efecto negativo. También se ha reportado un aumento del 17 % de la biomasa seca de parte aérea en plantas de tomate injertadas sometidas a estrés por arsénico (100 µg L-1 de As (V) en solución nutritiva) (Stazi et al., 2016).
Existen mecanismos que utiliza la planta para desintoxicarse como la quelación con polipéptidos como el glutatión (GSH) y las fitoquelatinas (PC), una vez quelado, el arsénico se almacena en las vacuolas de las raíces y no causa daño a las plantas (Liu et al., 2010). Independientemente de los mecanismos de acumulación y desintoxicación de arsénico, la mayor parte del arsénico en los tejidos probablemente sea fisiológicamente inerte (Santos et al., 2010). Además, el efecto hormético podría ser otra respuesta provocada por el arsénico. Esta se caracteriza por la estimulación a bajas concentraciones de metales pesados e inhibición a altas concentraciones q ue toma la forma concentración-respuesta de U o U invertida (Agathokleous et al., 2019). Esta forma de U invertida puede ser causada por hormesis desencadenada por iones de elementos no esenciales (Poschenrieder et al., 2013).
El efecto estimulatorio de las NPs se reporta generalmente por dosis bajas, con respecto a las SiO2 NPs, se ha reportado un incremento del 25 % en la biomasa radicular seca y del 75 % en parte aérea en plantas de fenogreco (Trigonella foenumgraecum) con aplicación de estas NPs (Nazaralian et al., 2017). En plantas de Pisum sativum sometidas a estrés por cromo, la aplicación de 10 μM de Si NPs mejora la producción de biomasa seca (Tripathi D. K. et al., 2015). Se ha informado que las Si NPs son beneficiosas, ya que pueden mejorar el crecimiento de plantas sometidas a estrés por metales pesados (Cui et al., 2017; Tripathi D. K. et al., 2015). Los efectos positivos de las SiO2 NPs sobre el crecimiento se pueden atribuir a la mejora del transporte de algunos elementos en la savia del xilema, además de la mejora de la capacidad de absorción de agua y fertilizantes (Janmohammadi et al., 2016). También otro mecanismo de las Si NPs es la estimulación de exudados ácidos a través de la raíz como el ácido oxálico y los polifenoles, que podrían reducir los efectos tóxicos del aluminio en el maíz (De Sousa et al., 2019).
El efecto estimulatorio puede ser fisicoquímico, y ocurre cuando la energía y las cargas superficiales de las NPs interactúan con las paredes y membranas celulares, modificando la actividad de los receptores, transportadores y otras proteínas (Zuverza-Mena et al., 2017). El proceso de absorción de NPs incluye mecanismos celulares como la señalización, reciclaje y regulación de la membrana plasmática, también se considera como un transporte activo (Tripathi D. K. et al., 2017). Una vez que las NPs entran en contacto con las plantas, estas pueden ser absorbidas y transportadas, generando diversos efectos como la estimulación de compuestos antioxidantes, mejorar el crecimiento de plantas sometidas a estrés abiótico, y en el peor de los casos provocar toxicidad (Cox et al., 2016; Pérez-Labrada et al., 2019; Zhang et al., 2018).
Micronutrientes
El presente estudio encontró que la aplicación de As en agua de riego disminuyó la absorción de micronutrientes (Fe, Cu, Zn) en plantas de tomate. La disminución de micronutrientes por exposición a As es consistente con lo reportado por Carbonell-Barrachina et al. (1997), ellos reportaron que la absorción de B, Cu, Zn y Mg en las plantas de tomate se reducía cuando estaban expuestas al arsenito. Gomes et al. (2012) reportaron una reducción de Fe que fue directamente proporcional al aumento de As en Anadenanthera peregrina.
El arsénico puede inducir estrés en la planta por el impacto que genera en la homeostasis de elementos esenciales, al reducir la absorción de algunos nutrientes (Kumar et al., 2015). Además, la exposición al As genera deformaciones anatómicas en la raíz, reduce las células del parénquima y el tamaño de las células del xilema (Tripathi P. et al., 2015), lo que puede afectar directamente la absorción de nutrientes. También se puede atribuir a la formación de complejos metálicos que no pueden ser absorbidos por la raíz, impidiendo así la absorción de elementos esenciales (Khan et al., 2019).
Los exudados de las raíces, pueden ser aminoácidos, ácidos orgánicos, azúcares, compuestos fenólicos y otros metabolitos secundarios (Haichar et al., 2014). Se ha informado que bajo estrés de cadmio la concentración de ácidos orgánicos (málico, cítrico, ácidos acético, oxálico, glutámico y fórmico) aumenta en plantas de maíz como mecanismo de tolerancia (Javed et al., 2017). Es probable que las plantas de tomate sigan esta misma estrategia, ya que la exposición al As induce la exudación de ácidos orgánicos por la raíz, y el transporte de As a los brotes es limitado, aumentando la acumulación en las raíces (Carbonell-Barrachina et al., 1997; Madeira et al., 2012; Stazi et al., 2016). La exudación radical puede estar involucrada en el proceso de estabilización, pero también podría secuestrar micronutrientes.
Bajo estrés metálico (iones metálicos, NPs metálicas), las plantas liberan más sustancias de bajo peso molecular como ácidos orgánicos (oxalato, acetato y malato) como defensa contra el estrés (Shang et al., 2019). En particular, los ácidos cítrico, oxálico y málico forman complejos con los metales, lo que afecta su fijación, movilidad y disponibilidad para las plantas (Xie et al., 2013). Se ha informado del efecto positivo de los ácidos orgánicos sobre la complejación del Cd, que se muestra hasta en un 85 % con ácido cítrico en la solución del suelo de plantas de Lupinus (Römer et al., 2000). Sin embargo, la modificación en la exudación de plantas sometidas a estrés metálico puede influir en la dinámica de los nutrientes del suelo y la actividad microbiana de la rizósfera (Jia et al., 2014). Además del hecho de que la capacidad de bombear protones por las H+-ATPasas en el plasmalema de la célula vegetal disminuye bajo estrés por metales, lo que puede afectar la asimilación de nutrientes (Javed et al., 2017).
Las SiO2 NPs indujeron una disminución en el contenido de Fe, sin embargo, se observó una tendencia positiva en la absorción de Cu y Zn. Le et al. (2014), reportaron que la concentración de nutrientes minerales como Cu y Mg en la parte aérea de algodón transgénico Bt se vio afectada por SiO2 NPs, el efecto negativo fue mayor en comparación con el algodón no transgénico. Los efectos adversos de las nanopartículas incluyen la reducción del alargamiento de la raíz, en diferentes especies de plantas como maíz, pepino, soya, repollo y zanahoria (Lin & Xing, 2007), esto podría provocar una disminución en la absorción de nutrientes. Sin embargo, los efectos positivos pueden deberse a la naturaleza mesoporosa de las Si NPs, esta característica puede ayudar en la agricultura debido a que actúan como nanoportadores de diferentes moléculas (Rastogi et al., 2019), y puede ser útil en nutrición vegetal. El nanosilicio aumenta los nutrientes del suelo, puede mantener el pH del suelo, e incluso estimula la comunidad bacteriana del suelo (Karunakaran et al., 2013). Estas mejoras en el sistema del suelo hacen que la disponibilidad de nutrientes para las plantas sea más eficiente.
Las características y la naturaleza de los nanomateriales, además de las especies vegetales, influyen en gran medida en la translocación y acumulación en los tejidos vegetales. El tamaño parece ser una de las principales restricciones para la penetración en los tejidos vegetales, además, en el suelo, las nanopartículas pueden interactuar con microorganismos y otros compuestos, lo que podría facilitar o dificultar su absorción (Pérez-de-Luque, 2017).
Tras la aplicación al suelo, las NPs pueden ingresar a la raíz, luego penetrar la pared celular/membrana plasmática, alcanzar la corteza de la raíz y entrar en los vasos del xilema, moviéndose así hacia arriba hasta las partes aéreas de la planta (Ma et al., 2010). Además, las NPs y NMs tienen una alta densidad de cargas superficiales capaces de interacciones inespecíficas con las cargas superficiales de las paredes celulares y membranas de las células vegetales que pueden provocar estimulación (Juárez-Maldonado et al., 2019). La adición de SiO2 NPs puede mejorar la proporción de células vivas al disminuir el estrés oxidativo después de la exposición al As. Estas NPs Pueden mantener la integridad de la célula, aumentar el espesor de la pared celular y la proporción de As en la pectina. Además, el suministro de SiO2 NPs regula la expresión de los genes que codifican los transportadores relacionados con As (Cui et al., 2019). Probablemente estas modificaciones que las SiO2 NPs inducen desde el punto de vista fisiológico-morfológico, bioquímico y molecular podrían equilibrar o incrementar la absorción de nutrientes esenciales.
Macronutrientes
Nuestros resultados mostraron en general un efecto negativo de la aplicación de arsénico en el agua de riego sobre los macronutrientes, aunque el P en las raíces tuvo una tendencia positiva, pero en la parte aérea la acumulación disminuyó. Roy et al. (2012), informaron que el As afecta la absorción de P, K y S en la parte aérea y la raíz de las plantas de amaranto, aunque solo la relación As y S mostró efectos negativos significativos. También se ha reportado que la aplicación de 5, 10, 20 y 50 mg L-1 de As como As (III) o As (V) en solución nutritiva, provocan una disminución de la concentración de K, Mg y P en las raíces y parte aérea de plantas de mezquite (Mokgalaka-Matlala et al., 2008).
La disminución en la acumulación de P probablemente se debió a la fitotoxicidad por As (Wang et al., 2002), o bien porque el As pudo haber reemplazado al P (Tu & Ma, 2005). Esto se debe a su analogía, ya que ambos tienen configuraciones electrónicas y propiedades químicas similares y compiten por los mismos transportadores de absorción (Meharg & Hartley-Whitaker, 2002). Por esta razón, el P también influye fuertemente en la absorción de As en las plantas (Anawar et al., 2018). Sin embargo, aunque el As puede reemplazar al P en la planta, no puede realizar sus funciones biológicas (Tu & Ma, 2005). En el presente estudio, la concentración de K mostró una tendencia creciente en raíces y disminuyó en la parte aérea. Resultados similares han sido reportados por Parson et al., (2008), ellos encontraron que el contenido de K en las plantas mejora en las raíces y disminuye la parte aérea, aquí el As redujo en 60% la concentración de potasio en la parte aérea. La relación positiva entre el arsénico y algunos nutrientes puede deberse a un “efecto de concentración” ya que altas dosis de As en solución disminuyen la biomasa de parte aérea y raíces (Melo et al., 2009). Mantener una nutrición suficiente con S puede ser particularmente importante en el ambiente contaminado con As, debido a que la desintoxicación de As a través de S ocurre por la formación de complejos del arsenito con péptidos ricos en tiol (Praveenet al., 2018).
El As causa diversos efectos sobre el contenido de macronutrientes, las respuestas serán diferentes en las especies de plantas y en cada nutriente, incluso para el mismo nutriente y especie (Khan et al., 2019). Además, también se ha sugerido que el estrés causado por el arsénico regula el estado nutricional de las plantas (Kumar et al., 2015). La aplicación de SiO2 NPs indujo un efecto positivo sobre la absorción y acumulación de macronutrientes. Tripathi D. K. et al. (2015), reportaron un aumento de K y P en la raíz y las hojas de las plántulas de Pisum sativum con la adición de solo Si NPs, y también mejoraron la absorción incluso cuando las plántulas estuvieron expuestas al cromo (VI). Alsaeedi et al. (2019) reportaron un aumento en la absorción y concentración de K (52 %, 75 % y 41 % en raíz, tallo y hojas respectivamente) de plantas de Cucumis sativus con la aplicación de Si NPs. El silicio tiene la capacidad de activar H+-ATPasas ubicadas en la membrana plasmática, lo que aumenta la absorción celular de potasio a través de gradientes electroquímicos, y canales y transportadores de K+ (Liang et al., 2006).
Las NPs son capaces de inducir reacciones de señalización en las células de la raíz (Sosan et al., 2016). Se sabe que la exposición de las plantas a las NPs puede modificar los patrones de crecimiento de las raíces al alterar su morfología (Dimkpa et al., 2015), generando efectos como la proliferación de pelos radiculares que podrían incrementar la absorción de nutrientes (Adams et al., 2017). Por otro lado, el aumento de las concentraciones de macronutrientes en las plantas por las SiO2 NPs puede estar relacionado con un “efecto de concentración”, ya que disminuyó la biomasa de parte aérea y raíces. Yang et al. (2020), informaron que la concentración de N y P aumenta en respuesta a la disminución de la altura de Leymus chinensis y Stipa krylovii. La concentración de Mg y Zn en la raíz, y K en las hojas de Pfaffia glomerata aumentó en respuesta a la disminución de la biomasa de raíces y hojas secas causada por la exposición a 50 μM de As (Gupta et al., 2013).
Conclusiones
Las plantas de tomate se vieron afectadas en su crecimiento por el As como por las SiO2 NPs. El As disminuyó la biomasa de raíces y parte aérea en concentraciones bajas, mientras que se estimularon en concentraciones altas. Las SiO2 NPs presentaron una influencia negativa por las concentraciones aplicadas. Sin embargo, las plantas no presentaron síntomas de toxicidad como clorosis o necrosis debido a la aplicación de As (V) y/o SiO2 NPs.
La absorción de Fe en raíz, y Cu en raíz y hojas disminuyó por la presencia de As (V) en la solución nutritiva. La concentración de Zn en la raíz aumentó, y la concentración de Fe en las hojas mostró un aumento, pero disminuyó con la dosis más alta de As (V). La concentración de P disminuyó en las raíces de manera dependiente de la concentración de As (V); en contraste, el efecto opuesto ocurrió en las hojas. El K y S no se vieron afectados por la presencia de As (V) en la solución nutritiva.
Las SiO2 NPs influyeron positivamente en la absorción y concentración de Zn y Cu en raíces, y Fe en hojas. Además, la absorción de macronutrientes en la raíz y las hojas aumentó con las SiO2 NPs.

