











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
Mesoamerica is the center of origin, domestication and diversification of several crops that currently are major components of world food, especially for Mexico. In this sense outstand species are maize (Zea mays L.) and beans (Phaseolus sp.) due to their contribution to familial economy and feeding, as well as their social-cultural value for marginalized communities of Mexico and low costs (Hernández-López et al., 2013; Chávez-Servia et al., 2016).
The bean species with a broad distribution in Mexico as wild, semi-domesticated or domesticated forms are: P. vulgaris L., P. coccineus ssp., P. lunatus L., P. acutifolius Gray, and P. dumosus (Hernández-Delgado et al., 2015). In Oaxaca have been found the species P. lunatus, P. coccineus, P. vulgaris and P. dumosus (Gepts, 2014). Domesticated and semi-domesticated species are known as natives, traditional or landraces, too and they own unique traits due to their adaptation to specific agroecological niches, derived from human or natural selection which combined with mutations, migration and/or genetic drift, resulted on special genotypes or genetic structure of populations. These genotypes are known by farmers by using special phenotypic traits or names as broad or narrow adaptability to diverse agroecosystems, tolerance to diseases, pod traits, precocity, the use of pods as ‘green beans’ or some other special uses. The study of traditional cropping and their species in its center of domestication allows us the understanding of evolution and diversification patterns and helps us to keep records about gene reservoir traits and its exploitation for breeding (Hernández-López et al., 2013).
Farmers mix seeds from different bean populations including varied seed coats as strategy to cushion variable climatic conditions (erratic rainfall patterns, extreme temperatures, pests, diseases, etc.) (Castillo-Mendoza et al., 2006; Espinosa-Pérez et al., 2015). Seed mixing modifies the genetic structure of bean populations, but the evolution of beans under this scheme is poorly understood (Chávez-Servia et al., 2016).
The study of genetic diversity into the center of origin and domestication of common beans is an easy and successful method to identify primary sources (populations) of useful genes for breeding, as well as knowledge to improve our understanding about evolution, diversification and in situ conservation of Phaseolus (Hernández-López et al., 2013). That information allows us to generate strategies for the genetic base broadening of released cultivars and for conservation, management and exploitation of the species (Hernández-Delgado et al., 2015).
The aim of this work was to characterize the morphological variability of one bean collection with indeterminate growth from the state of Oaxaca, México under greenhouse conditions. The hypothesis was that there are significant morphological and agronomic variability in the bean germplasm from Oaxaca, and variability is related to geographic origin.
Material and Methods
Genetic material
The work included 75 bean populations collected from November 2015 to March 2016 through different agroecological regions of Oaxaca. During collection was geopositioned the community and municipality of each collection, sample origin, local name, outstand trait, seed coat color, among other collection descriptors. The ethnolinguistic group to which belongs each collection was also registered (Figure 1).

Seed and plant description
Plants were developed under greenhouse conditions at CBG-IPN in Reynosa, Tamaulipas. Pots of 5 L were filled with 5-kg regional agricultural soil and irrigated with 750 mL of tap-water every 2-4 days as been needed. Sowing was carried out by August 30, 2016, five seeds per pot and three pots per accession (replications). Pots were randomized in a completely randomized design. Four controls were included: cultivars Negro Jamapa, and Negro Comapa; Pinto Villa, and Pinto Saltillo (released cultivars) (Rosales-Serna et al., 2004).
After sowing days to emergence and color of hypocotyls, are registered as well as days to the appearance of primary leaflets, the first, second and third trifoliate leaves. After, we registered the days to flowering, to pod formation and filling, and days to physiological maturity; color of flowers, growth habit (Debouck & Hidalgo, 1984), and leaf area. To harvest the primary and secondary seed colors were registered (Munsell color Company, 1977), as well as brightness, mottle patterns and seed shape (IBPGR, 1982). After, length, width and thickness of seeds were measured (Espinoza-Pérez et al., 2015) and seed sizes were classified (Singh, 1989; Singh et al., 1991).
Statistical analysis
Data were registered in Excel (MS-2010). For qualitative variables relative frequencies by each class, the region of origin, and the ethnolinguistic group were registered. Quantitative data were subjected to principal component analysis (PCA) by using the statistical software SAS® version 9.0.
Results and Discussion
All regions of origin of bean germplasm showed several seed colors, mixtures or ‘ensaladilla’ (mixture type which includes till eight seed types) which belongs to Mixes. Ethnolinguistic groups that showed the highest diversity of seed colors were Mazatecos in Cañada and Mixes at Sierra Norte. Seed coat colors more commons were yellow (5), black (1), and red (3) and purple (2) tonalities. Mottle pattern more frequently was absent for most of the regions, followed by the pattern ‘splashed’ dots. Region Mixteca presented the highest variation on mottled patterns, while Istmo and Sierra Sur reported bean populations with one single color of seeds. All regions presented populations with shiny seed coat, populations with matte seed coat were found at Mixteca and Sierra Sur regions. The other populations maintain seed mixtures of shiny and matte seed coats. Purple was the most frequent color of flowers and hypocotyls; the most common growth habit was indeterminate-climber (type IV) while seed shape was variable among regions (Table 1).
Table 1 Frequencies of qualitative traits among bean populations from Oaxaca’s farmers by region and ethno-linguistic group.
| Origin of populations | Brightness | Primary color of seed | Secondary color of seed | Flower color | Growth habit | Hypocotyl color | Mottled pattern | Shape | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Class | FR(%) | Class | FR (%) | Class | FR(%) | Class | FR(%) | Class | FR(%) | Class | FR(%) | Class | FR(%) | Class | FR(%) | |
| Cañada (Mazatecos) |
1 | 3.3 | 5 | 66.6 | A | 63.3 | 2 | 6.67 | 3 | 23.3 | 1 | 26.6 | 0 | 83.3 | 1 | 46.7 |
| 2 | 96.6 | 3 | 13.3 | 1 | 23.3 | 5 | 33.3 | 4 | 76.6 | 5 | 40.0 | 4 | 10.0 | 2 | 23.3 | |
| Others (3) | 20.0 | Others (3) | 13.3 | 6 | 60.0 | 7 | 33.3 | Others (2) | 6.7 | |||||||
| Istmo (Zapotecos) |
1 | 100 | 1 | 77.7 | A | 100 | 1 | 22.2 | 1 | 100 | 1 | 26.6 | 0 | 100 | 3 | 22.2 |
| 5 | 22.2 | 6 | 77.7 | 2 | 40.0 | 5 | 33.3 | |||||||||
| 7 | 33.3 | Others | 44.4 | |||||||||||||
| Mixteca (Mixtéeos) |
0 | 9.0 | 3 | 30.0 | A | 89.0 | 1 | 75.0 | 2 | 6.0 | 1 | 61.0 | 0 | 91.0 | 2 | 41.0 |
| 1 | 16.0 | 1 | 28.0 | 5 | 6.0 | 6 | 16.0 | 4 | 94.0 | 4 | 9.0 | 4 | 3.0 | 5 | 20.0 | |
| 2 | 75.0 | Others (3) | 42.0 | Others (2) | 5.0 | 3 | 9.0 | Others (4) | 29.6 | Others (3) | 6.0 | Others (3) | 39.0 | |||
| Sierra Norte (Mixe) |
1 | 15.7 | 2 | 26.3 | A | 57.8 | 1 | 10.5 | 3 | 36.8 | 1 | 15.7 | 0 | 75 | 1 | 10.5 |
| 2 | 84.2 | 5 | 42.1 | 1 | 31.5 | 4 | 84.2 | 4 | 63.1 | 7 | 84.2 | 3 | 15 | 2 | 31.6 | |
| Others (3) | 31.5 | 5 | 10.5 | 6 | 5.2 | Others (2) | 10 | 3 | 57.8 | |||||||
| Sierra Norte (Zapotecos) |
1 | 22.7 | 7 | 31.8 | 0 | 86.3 | 4 | 27.2 | 4 | 100 | 4 | 36.3 | 0 | 86.4 | 2 | 72.7 |
| 2 | 77.2 | 5 | 22.7 | 1 | 9.0 | 6 | 72.7 | 5 | 22.7 | 8 | 4.5 | 4 | 13.6 | |||
| Others (3) | 45.4 | 5 | 4.5 | Others (2) | 40.9 | 9 | 9.1 | Others (2) | 13.6 | |||||||
| Sierra Sur (Zapotecos) |
0 | 38.1 | 1 | 71.4 | A | 100 | 1 | 4.7 | 4 | 100 | 1 | 19.0 | 0 | 100 | 5 | 38.1 |
| 1 | 23.8 | 4 | 14.2 | 6 | 95.2 | 5 | 14.2 | 3 | 33.3 | |||||||
| 2 | 38.1 | Others (2) | 14.2 | 7 | 66.6 | Others (2) | 28.2 | |||||||||
| Valles Centrales (Zapotecos) |
0 | 84.6 | 1 | 46.1 | A | 84.6 | 1 | 53.8 | 2 | 53.8 | 3 | 53.8 | 1 | 92.3 | 5 | 38.5 |
| 2 | 15.3 | 5 | 53.8 | 2 | 15.3 | 6 | 46.1 | 4 | 46.1 | 5 | 15.3 | 7 | 7.7 | 4 | 30.8 | |
| 7 | 30.7 | Others (2) | 30.8 | |||||||||||||
Brightness: 0= matte, 1= brilliance, 2= both; Primary and secondary seed color (Munsell color Company, 1977): 0, 1, 2, 3, 4, 5, 6, and 7; flower color: 1, 2, 3, 4, y 5; growth habit: 1= determined type I, 2= determined type II, 3= in-determined type III, 4= in-determined type IV; hypocotyl color 1, 2, 3, 4, 5, 6, 7; mottled patterns 0, 1, 3, 4, 7, 8, 9; shape (IBPGR, 1982): 1, 2, 3, 4, 5.
Seeds with the largest weight and thickness were from Mixteca while the smallest were from Istmo. Bean populations of Zapoteca ethnicity from Sierra Norte showed the highest seed length while collections from Mixe’s communities of Sierra Norte reported the highest thickness. Populations from Cañada and Mixteca exhibited the greatest variation in 100-seeds weight and the lowest variation was found in beans of Zapotecas from Valles Centrales. These last were the earliest accessions to flowering (76 days after sowing) while accessions from Mixes were the latest (102 days). Days to emergence varied from 4 to 5 days after sowing among regions and the first trifoliate leaf was produced to 25 days after sowing on average (Table 2).
Table 2 Averages of morphological and agronomical quantitative traits in bean populations grouped by region and ethno-linguistic group origins.
| Traits | Cañada | Istmo | Mixteca | Sierra Norte | Sierra Sur | Valles Centrales | Controls | |
|---|---|---|---|---|---|---|---|---|
| Mazateco | Zapoteco | Mixteco | Mixe | Zapoteco | Zapoteco | Zapoteco | ||
| 100-seed weight | 35.13 | 19.4 | 36.03 | 35.2 | 31.6 | 26.9 | 32.3 | 17.3 |
| Length | 7.76 | 6.00 | 8.27 | 7.5 | 8.01 | 7.57 | 8.02 | 5.55 |
| Width | 3.73 | 2.75 | 4.85 | 4.7 | 3.91 | 3.42 | 3.90 | 2.35 |
| Thickness | 2.08 | 1.41 | 2.50 | 2.5 | 2.15 | 1.5 | 1.60 | 0.75 |
| No. of colors | 7.5 | 2.2 | 2.9 | 8.0 | 4.4 | 2.1 | 3.2 | 1.0 |
| Days to flowering | 94.0 | -- | 85.0 | 102.0 | 88.4 | 95.9 | 76.2 | 73.0 |
| Days to emergence | 4.5 | 4.7 | 4.7 | 5.0 | 5.0 | 4.8 | 5.0 | 5.0 |
| Leaf area | 69.5 | 92.1 | 107.3 | 67.6 | 106.1 | 97.7 | 109.3 | 70.2 |
| Days to primary leaves | 10.2 | 9.5 | 8.18 | 8.5 | 9.4 | 8.6 | 8.5 | 11.5 |
| Days to first trifoliate leaves | 17.0 | 14.0 | 13.7 | 13.0 | 13.8 | 13.5 | 11.0 | 14.5 |
| Days to third trifoliate leaves | 21.2 | 23.0 | 20.7 | 20.0 | 20.2 | 21.6 | 19.7 | 32.0 |
The ACP explained 90 % of the total variance observed into the first two principal components (PCs). At first PC leaf area (0.99), 100-seeds weight (0.062) and days to pod formation (0.031) showed the highest descriptive value, while for the second PC the highest descriptive variables were 100-seeds weight (0.30); days to formation (0.42), filling (0.616), and pod maturity (0.58). The spatial distribution of bean populations by region of origin indicated the high variability on seed traits, leaf area, and physiological stages (Figure 2a).
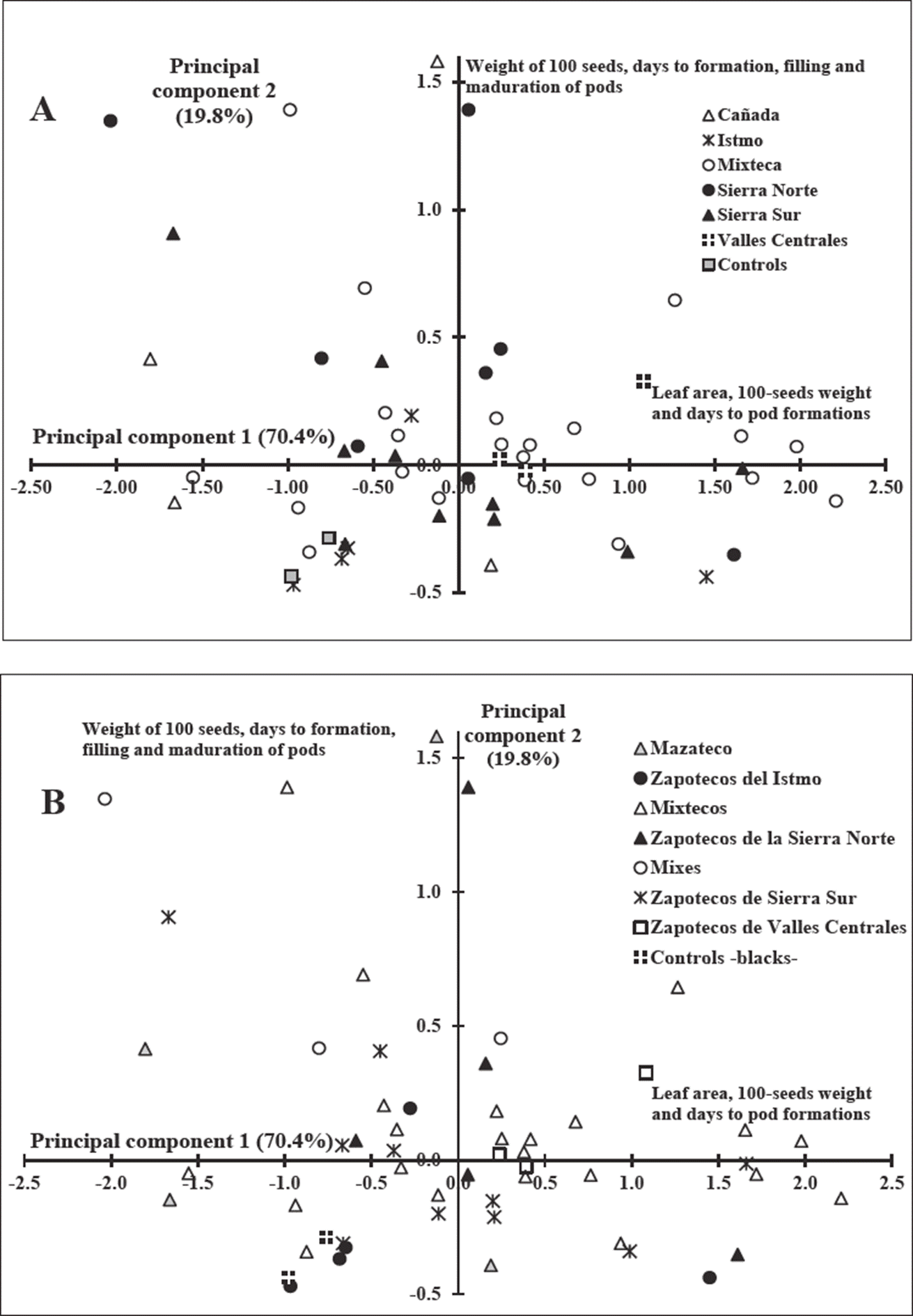
Figure 2 Dispersion of bean germplasm based on the principal components 1 and 2 of the principal component analysis of morphologic and agronomic data, classified by Oaxaca region (A) or ethno-linguistic group (B) origins.
Phenotypic diversity on seedling and seed traits as well as on phenological phases are the reflection of bean diversity preserved by the farmers of Oaxaca. The PCA based on ethnolinguistic group indicated similitude of germplasm of Zapotecos from Valles Centrales, Sierra Norte, and Sierra Sur. However, Zapotecos from Istmo have germplasm a little different from those mentioned (quadrant IV, Figure 2b). Bean populations cultivated by Mazatecos and Mixes were more dispersed. In addition, there were differences among communities and within ethnic groups regarding the way of conservation, selection and maintaining their bean diversity; and these differences were expressed on traits evaluated in this work.
The morphological and physiological descriptions of one bean collection from the state of Oaxaca, Mexico indicated variability on qualitative and quantitative traits and showed the differences among regions of origin and ethnic groups that conserved them. Phenotypic differentiation started since the identification with a name or trait supplied by the farmer; for example, beans of ‘milpa’ (blacks) or ‘ejotero’ and ‘grueso’ (‘thick’ bean) (P. coccineus) showing evident phenotypic differences. Also outstand ‘chivo’ (goat) beans (black seeded) from Sierra Sur and Valles Centrales; ‘machetudo’ (‘machete’) beans (Mixteca; purple seeds) or ‘cuarenteño’ (forty days) (Istmo, yellow seeds). Espinosa-Pérez et al. (2015) classified in 13 groups of bean genotypes based on seed colors and local names used by farmers in order to distinguish bean germplasm from several regions of Mexico, including Oaxaca, and where the more frequent were genotyped with seed colors yellow, ‘ensaladilla’, red, black and ‘vaquita’ (white or beige seeds with black spots).
Bean populations from Mixteca showed large seeds and highly variable on colors and mottled patterns, while populations with small seeds and less variable in colors come from Istmo and Sierra Sur regions. Morpho-agronomic variables that differentiated populations were leaf area; 100-seeds weight; days to pod formation, pod filling, and pod maturity. Espinoza-Pérez et al. (2015) reported that 100-seeds weight and seed thickness can discriminate bean populations from Oaxaca’s Phaseolus gene pools. Aquino-Bolaños et al. (2016) indicated that small seeds with low specific weight are also found in Oaxaca’s germplasm and additionally, bean populations with dark seed coats showed the highest levels of monomeric anthocyanins.
In this work, it was not detected any relationship between origin and clustering based on seed traits, contrary to that was reported by Rocandio et al. (2009). However, bean germplasm was clustered in accordance with ethnicity origins as Sierra Norte, and Zapotecas and Mixes, while the populations from Mixteca were more variable. Germplasm from Mixteca showed large seeds and were morphologically variables. Mixtecos have a higher number of local terms or classifications to its local bean diversity, compared with Mazatecos or Mixes.
Seed management practices for sowing and culture are different among ethnic groups; for example, Mixes mix up eight colors seeds (genotypes) to establish their ‘milpas’ every year. Mazatecos from Cañada and Mixes from Sierra Norte uses a higher color seed diversity of beans, outstanding yellow, black, red, and purple. In this work predominated genotypes with purple flowers and hypocotyls, as well as growth habit IV or called as indeterminate climber growth habit. Populations from Valles Centrales were earlier than the germplasm of Mixes from Sierra Norte. At regional markets, the most common seed colors are related to their commercial, cultural, and/or culinary value, either dark or light seed coat colors (Rocandio et al., 2009).
Chávez-Servia et al. (2016) reported 92 days to flowering on average for bean populations from Sierra Sur, 84.7 days in germplasm from Valles Centrales, and 44.3 days for beans from Mixteca. In this work, flowering happened to 95, 76, and 85 days for germplasm from each origin, respectively. Bean populations from Sierra Norte presented 88 days to flowering while beans from Zapotecos and Mixes showed 88-102 days, respectively.
Soleri et al. (2013) detected P. coccineus populations from Sierra Norte of Oaxaca and cultivated by Zapoteca ethnicity. In this work, ayocote beans were detected in the Mixe ethnicity in the same region, too. Likewise, we found that seed with the lowest weight belongs to P. vulgaris (30 g per 100 seeds), followed by P. coccineus (42 g), and P. dumosus (52 g). Worthington et al. (2012) reported that the genetic race of each accession is associated with the range of altitude where the genotype is cultivated, derived from its strong adaptation to specific agroecological niches.
The PCA performed two groups of genotypes in accordance to seed traits, one includes beans with small seeds and black seed coats that are commonly cropping as a monoculture, with determined growth habit; this germplasm is similar to controls Negro Jamapa and Comapa. The other groups included that germplasm showing less seed color and size diversity, as well as indeterminate growth habit (Soleri et al., 2013; Worthington et al., 2012; Espinoza-Pérez et al., 2015).
Beans from Oaxaca are one result of in situ conservation by local farmers, one process conducted by generations of agricultural families with limited economic resources to which bean grain constitutes the base of their feeding. In addition to the conservation of genetic resources, farmers preserve local knowledge associated with the cultural practices, grain conservation, and criteria for seed selection for the next crop cycle (Espinoza-Pérez et al., 2015). Current diversity was produced by selection of beans for specific agricultural systems, increasing its potential genetic value (Singh et al., 1991; Soleri et al., 2013).
Las milpas y backyards (home gardens) are spaces for bean production in Oaxaca, where farmers host the most of bean genetic diversity, although this diversity is not yet fully studied. The diversity is maintained and enriched with farmer’s practices as seed exchange (Aguilar-Støen et al., 2009; Espinoza-Pérez et al., 2015) and the likely genetic flux among P. vulgaris, P. coccineus and P. dumosus within the spaces where those species converge. These practices expand the diversity managed by each Farmer and have positive effects as the likely resilience to climate change or either, changes of soil use, erosion, or rainfall patterns (Aguilar-Støen et al., 2009).
Bean domestication increased the seed size, changed growth habit from indeterminate to determinate and reduced pod dehiscence; then it was produced genetic variability on size, shape, and color of seeds (Peña-Valdivia et al., 2012).
Beans from four regions of the state of Oaxaca showed significant differences on flowering, pod length or the number of seeds per pod; this difference was higher among populations from Mixteca compared with those from Valles Centrales and Sierra Norte (Chávez-Servia et al., 2016). Singh (1988) reported a 100-seeds weight of 27.5 g in bean germplasm from Oaxaca, and Espinoza-Pérez et al. (2015) informed more than 20 g per 100-seeds and seed coat mostly black. Worthington et al. (2012) described bean populations from one community at Sierra Juárez de Oaxaca differentiating germplasm of P. vulgaris, P. dumosus and P. coccineus species based on seed size, flower color, type of emergence (epigeal or hypogeal) or local classification system on Zapoteca’s language.
Conclusions
Morpho-agronomic variability analyses of the bean collection from different regions of Oaxaca, Mexico showed significant differences among and within populations, and these differences were less frequent among regions of origin. The identified groups were Mixteca-Istmo, Sierra Norte-Mixteca, and Cañada-Sierra Sur-Valles Centrales regions.

