










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista bio ciencias
versión On-line ISSN 2007-3380
Revista bio ciencias vol.7 Tepic 2020 Epub 18-Nov-2020
https://doi.org/10.15741/revbio.07.e720
Artículos originales
Impacto del cambio climático en las áreas con aptitud ambiental para Bouteloua gracilis y Bouteloua repens en México
1 Profesor Asociado, Universidad de Guadalajara. Departamento de Ciencias Pecuarias y Agrícolas, Centro Universitario de Los Altos.
2 Profesor Investigador, Universidad de Guadalajara. Centro Universitario de Ciencias Biológicas y Agropecuarias.
Las áreas de pastizal se caracterizan por tener una cobertura de suelo en la que predominan especies de la familia Poaceae, entre las que sobresale el género Bouteloua por su calidad como forraje y sus características ecológicas. La temperatura y la precipitación son determinantes para la productividad primaria neta de los pastizales, en los que el cambio climático podría provocar cambios en la estructura de la vegetación, por lo cual esta investigación se realizó con el objetivo de analizar el impacto del cambio climático en las áreas con aptitud ambiental (EF) para las gramíneas B. gracilis y B. repens en México. Se analizaron dos escenarios de cambio climático, con las rutas de concentración representativas 4.5 y 8.5 en los periodos 2040-2060 y 2060-2080. Se utilizaron 28 variables ambientales con el modelo Maxent para analizar las áreas con EF para ambas especies. Los resultados mostraron que la temperatura media anual aumentará entre 2.8 °C y 5.0 °C, con cambios más acentuados en zonas áridas y semiáridas; la precipitación anual disminuirá entre 3.1 % y 14.4 %, con cambios principalmente en zonas con promedios menores a 600 mm. Debido al cambio climático, se presentarán nuevas áreas con EF en algunas zonas del país y habrá pérdida en otras, aunque de manera general se tendrá mayor área con EF, con incrementos entre 6.2 % y 9.1 % en relación con la superficie actual para Bouteloua gracilis y entre 13.4 % y 16.5 % para B. repens.
Palabras clave: Bouteloua gracilis; Bouteloua repens; aptitud ambiental; cambio climático; Maxent
Grassland areas are characterized for having a ground cover which species from the Poaceae family predominate, among which Bouteloua genus stands out due to its quality as a fodder and its ecological characteristics. Temperature and precipitation are determinants for net primary productivity of grasslands, in which climate change could cause changes in the vegetation structure. Therefore, the objective of this research was to analyze the impact of climate change on areas with environmental fitness (EF) for Bouteloua gracilis and B. repens grasses in Mexico. Two climate change scenarios were analyzed, with representative concentration pathways of 4.5 and 8.5 for 2040-2060 and 2060-2080 periods. Twenty-eight environmental variables were used with Maxent model to analyze areas with EF for both species. Results showed that the annual average temperature will increase from 2.8 °C to 5.0 °C, with noticeable changes in arid and semiarid zones; the annual average precipitation will decrease from 3.1 % to 14.4 %, mainly in zones with averages lower than 600 mm. New areas with EF will appear in some zones of the country and some others will disappear due to climate change, although in generally there will be greater area with EF, with increases from 6.2% to 9.1 % for Bouteloua gracilis and from 13.4 % and 16.5 % for B. repens compared with the current surface.
Keywords: Bouteloua gracilis; Bouteloua repens; environmental fitness; climate change; Maxent
Introducción
Las áreas de pastizal nativo y las áreas de pastoreo de otros tipos de vegetación se caracterizan por tener una cobertura de suelo en la que predominan especies herbáceas de la familia Poaceae, de las cuales existen en México 205 géneros que agrupan a 1,216 especies (Dávila et al., 2018). Diversas investigaciones han mostrado que los géneros con mayor presencia en estas áreas son: Bouteloua, Aristida, Andropogon, Muhlenbergia, Sporobolus y Heteropogon, entre otros, que tienen una cobertura del suelo entre el 7.3 % y 49.8 % de la superficie, en los estados de Chihuahua y Durango (Rodriguez-Maturino et al., 2017); así como Boteloua gracilis y Muhlenbergia rigida en el noreste de Jalisco, con cobertura entre 35 % y 40 % en áreas con pastoreo moderado y de 5% y 10 % en áreas con sobrepastoreo (Delgado-Balbuena et al., 2013).
Las especies del género Bouteloua están entre las más importantes en los pastizales de México, por su calidad alimenticia para el ganado y sus características ecológicas (Herrera et al., 2011), entre las que destacan el zacate Chino Bouteloua repens (Kunth) Scribn. & Merr. y el zacate Navajita Bouteloua gracilis (Kunth) Lag. ex Griffiths, esta última ha sido incluida en programas de mejoramiento de pastizales por sus características forrajeras sobresalientes (Beltrán et al., 2010; Morales et al., 2009). Además, el género Bouteloua destaca por su diversidad con 37 especies y 14 variedades en México y se han encontrado sitios donde las plantas de este género producen hasta el 90 % de la biomasa vegetal (Giuliani et al., 2014).
Resultados de investigaciones indican que en las siguientes décadas se presentarán cambios en las variables del clima por efecto del calentamiento global en la mayor parte del territorio mexicano y del planeta (Durán et al., 2014; Ruíz et al., 2011, Zarazúa et al., 2011); entre tales cambios sobresalen la temperatura y la precipitación pluvial debido a que son determinantes climáticos para los procesos fisiológicos de las plantas, lo que se manifiesta de manera notable en la productividad primaria neta de los ecosistemas (Pérez-Ramos et al., 2012).
El clima es un determinante de la productividad primaria en los ecosistemas, por lo que el cambio climático tendrá efecto en las comunidades vegetales modificando a su vez los servicios y productos que proveen estas áreas (Sage & Zhu, 2011; Reeves et al., 2014), lo que ha motivado realizar estudios para evaluar el impacto del cambio climático en comunidades vegetales. Al respecto, Bocksberger et al. (2016), al analizar la distribución de 302 especies de gramíneas encontraron que la precipitación fue la variable que ejerció mayor influencia en la distribución de las especies y que la adaptación en ambientes más secos está dada por el tamaño más pequeño de las plantas y por la ruta fotosintética tipo C4; por su parte, Martinson et al. (2011), consignan que las especies de Bouteloua mostraron una relación negativa a la precipitación, lo que sugiere una ventaja adaptativa a condiciones de sequía. La presente investigación se realizó con el objetivo de analizar el impacto del cambio climático en las áreas con aptitud ambiental para las gramíneas B. gracilis y B. repens en México.
Material y Métodos
La Investigación se llevó a cabo utilizando el modelo de Máxima Entropía (Phillips et al., 2009), referido como Maxent, para modelar el nicho ecológico y predecir el área con EF para las gramíneas B. gracilis y B. repens; este modelo se ha utilizado ampliamente para estimar las distribuciones potenciales de especies vegetales en general (Durán et al., 2016; Franklin et al., 2013) y para gramíneas (Meneses et al., 2014), con base en las condiciones ambientales más significativas, usando solamente datos de presencia de la especie.
La investigación incluyó el análisis de los datos de clima de referencia y clima futuro obtenidos del portal de Worldclim versión 2 (Fick & Hijmans, 2017), correspondientes al modelo de circulación general GFDL-CM3; se usaron los datos del periodo 1960-1990 como clima de referencia y de los periodos 2041-2060 y 2061-2080, referidos más adelante como periodos 2050 y 2070, respectivamente, con una resolución espacial de 30 segundos de arco terrestre (Hijmans et al., 2005). Así mismo, en la simulación se usaron las rutas de concentración representativas (RCP, por sus siglas en inglés) de 4.5 y 8.5 W m-2 (Van Buuren et al., 2011).
A partir de los datos de clima mundial obtenidos se extrajeron los correspondientes a México y se manipularon a través de capas en formato raster con el uso de la aplicación ArcMap 10.2, posteriormente se transformaron a formato ASCII, para obtener las capas de las variables ambientales con el sistema Idrisi Selva 17.0 (Eastman, 2016). Las capas resultantes a su vez fueron analizadas con el modelo Maxent para analizar las áreas con EF para las especies de B. gracilis y B. repens.
El análisis de distribución potencial de las dos especies realizado con el programa Maxent se basó en referencias geográficas proporcionadas en grados decimales de latitud y longitud, de los sitios de colecta de ambas especies. Se utilizaron 501 referencias geográficas de B. gracilis y 451 de B. repens, que fueron obtenidas de Global Biodiversity Information Facility (GBIF, 2017).
El modelo utilizado en Maxent incluyó el análisis de 28 variables ambientales: precipitación acumulada anual (mm), precipitación acumulada de mayo a octubre (mm), precipitación acumulada de noviembre a abril (mm), precipitación acumulada de diciembre a febrero (mm), precipitación del mes más húmedo (mm), precipitación del mes más seco (mm), temperatura máxima anual (°C), temperatura máxima de mayo a octubre (°C), temperatura máxima de noviembre a abril (°C), temperatura media anual (°C), temperatura media de mayo a octubre (°C), temperatura media de noviembre a abril (°C), temperatura media del mes más cálido (°C), temperatura media del mes más frío (°C), temperatura mínima anual (°C), temperatura mínima de mayo a octubre (°C), temperatura mínima de noviembre a abril (°C), fotoperiodo de mayo a octubre (H), fotoperiodo de noviembre a abril (H), índice de humedad anual (mm), índice de humedad de mayo a octubre (HI), índice de humedad de noviembre a abril (HI), índice de humedad de diciembre a febrero (HI), oscilación térmica anual (°C), oscilación térmica de mayo a octubre (°C), oscilación térmica de noviembre a abril (°C), oscilación térmica de diciembre a febrero (°C) y textura del suelo.
El modelo Maxent se ajustó para 25 % de los datos para prueba aleatoria y 75 % para entrenamiento, 10 replicaciones con la prueba de validación cruzada y 2500 interacciones para convergencia; además, se solicitó el formato de salida Logistic y la prueba Jackknife para analizar la importancia de cada variable ambiental. Dado que los resultados obtenidos con el algoritmo Maxent son influidos por los sitios de muestreo con que se alimenta el programa (Phillips & Dudik, 2008; Elith et al., 2011), se requiere fijar un valor umbral para estimar las áreas con EF para cada especie (Norris, 2014; Liu et al., 2016), por lo que en esta investigación se utilizó como umbral logístico el valor 10 acumulativo fijo (Fixed cumulative value 10 logistic threshold, en el modelo Maxent). Las áreas con EF obtenidas con Maxent fueron examinadas con el sistema Idrisi y se generaron mapas con los valores correspondientes a este umbral, a partir de los datos arrojados por la aplicación del modelo en el clima de referencia y cada escenario de cambio climático.
Se excluyeron las áreas ocupadas por los cuerpos de agua y centros urbanos en el cálculo de la superficie potencial de distribución de ambas especies; estas capas temáticas se obtuvieron mediante la carta del uso del suelo y vegetación (INEGI, 2009).
Resultados y Discusión
Ajuste del modelo
En la presente investigación, los valores de la curva operacional o curva ROC, la cual representa el ajuste de los datos de muestreo, fueron de 0.957 para B. gracilis en todos los escenarios climáticos y de 0.928 a 0.930 para B. repens (Tabla 1), la curva de los datos de entrenamiento y la curva de los datos de prueba se sitúan cercanas entre sí, lo que indica que no hay error de omisión (Hernández et al., 2018). Datos congruentes con el valor de 0.946 obtenido en otra investigación para B. curtipendula (Martinson et al., 2011). Los valores obtenidos con este modelo son satisfactorios, e indican la bondad para clasificar las áreas con aptitud ambiental de ambas especies (Parolo et al., 2008).
Tabla 1 Área bajo la curva (AUC) y valores de la desviación estándar (SD) para Bouteloua gracilis y B. repens en el clima de referencia y cuatro escenarios de cambio climático en México.
| Climate period | B. gracilis | B. repens |
|---|---|---|
| 1960-1990 | 0.957 ± 0.006 | 0.928 ± 0.010 |
| rcp 4.5/2050 | 0.957 ± 0.007 | 0.930 ± 0.009 |
| rcp 4.5/2070 | 0.957 ± 0.007 | 0.930 ± 0.010 |
| rcp 8.5/2050 | 0.957 ± 0.007 | 0.930 ± 0.010 |
| rcp 8.5/2070 | 0.957 ± 0.007 | 0.929 ± 0.010 |
Cambio climático en México
El análisis de la temperatura media anual predice que habrá un aumento de 2.8 y 3.4 °C para el 2050 y de 3.4 y 5.0 °C en el 2070, con el rcp 4.5 y el rcp 8.5 respectivamente. Estos resultados concuerdan con investigaciones previas, como lo refieren Ruiz et al. (2011), Durán et al. (2014), quienes consignan un aumento en la temperatura entre 1.8 y 3.3 °C en diversas áreas de México y Galindo et al. (2015), mismos que mencionan un incremento de 2.5 °C.
Es importante señalar que los cambios en la temperatura mínima anual y temperatura máxima anual, serán diferentes para cada región geográfica, con tendencia general a que sea más acentuado en las áreas áridas y semiáridas que en las áreas templadas y tropicales; En el presente estudio los resultados mostraron un aumento en la temperatura máxima y un aumento en la frecuencia en los escenarios futuros; con el clima de referencia de pico de temperatura máxima de 30 °C con una frecuencia de 12.3 %; en el 2050 se pronostica que se elevará a 33 °C con frecuencia de 14.6 % y a 34 °C con frecuencia de 15.2 %, y para el 2070 se incrementará a 35.0 °C y frecuencia de 15.1 % y 36.0 °C y frecuencia de 17.3 % con el rcp 4.5 y rcp 8.5, respectivamente. La temperatura mínima también aumentará en los escenarios futuros, aunque no cambiará la frecuencia; con el rcp 4.5 la temperatura se incrementará en 2.25 °C en el 2050 y en 2.65 °C en el 2070, y con el rcp 8.5 el aumento será de 2.73 °C y de 4.11 °C.
Los resultados para la precipitación anual reflejan que habrá una disminución de 3.1 % en el 2050 y 7.0 % en el 2070, con el rcp 4.5; y de 6.5 % y 14.4 % con el rcp 8.5. Al respecto, en otras investigaciones los resultados muestran que la precipitación en México disminuirá del 2 a 12 % en el mismo periodo (Durán et al., 2014), entre 3.8 y 4.5 % (Ruíz et al., 2011), y del 10 % (Galindo et al., 2015). Estos cambios se presentarán principalmente en las zonas con precipitación menor a 600 mm, donde prolifera en gran medida la vegetación nativa de matorral xerófilo y pastizal natural, y se ubican principalmente en el centro, noreste y noroeste de México; el estudio pronostica también que, habrá ligeros incrementos en las zonas con precipitación superior a los 2000 mm anuales. Por lo anterior, una menor disponibilidad de humedad en el suelo, junto con un incremento en la temperatura media anual, en las áreas de pastizal provocaría alteraciones en la estructura y en la productividad primaria neta (Edwards & Smith, 2010; Martinson et al., 2011; Bocksberger et al., 2016).
Áreas con aptitud ambiental
Las áreas con EF para B. gracilis estimadas, se distribuyen principalmente en la región del Altiplano Mexicano (Figura 1, lado izquierdo), ubicadas principalmente en Chihuahua, Durango, Zacatecas, Aguascalientes y Guanajuato, parte del norte de Jalisco, sur de Coahuila, sur de Nuevo León y pequeñas zonas en Querétaro, Hidalgo, Estado de México, Morelos y Puebla. De manera general, la vegetación nativa de estas áreas es pastizal y/o matorral xerófilo. Los resultados del estudio son congruentes con las áreas de distribución para la especie, descritas por Cortés et al. (2013), quienes indican que las áreas con mayor probabilidad para la presencia de B. gracilis tienen entre 400 y 800 mm de precipitación anual y una temperatura media anual de 13 a 19 °C; así mismo, las áreas definidas en la presente investigación son acordes con las áreas de distribución (Coahuila, Nuevo León y Tamaulipas) señaladas por Herrera et al. (2008).

Figura 1 Área con aptitud ambiental para Bouteloua gracilis (imagen del lado izquierdo) y B. repens (lado derecho) en el clima de referencia. Los puntos indican sitios de colecta para ambas especies.
Por su parte, B. repens (Figura 1, lado derecho) tienen una amplia distribución en la región central del país, ya que cubre gran parte de la superficie en los estados de Jalisco, Aguascalientes, San Luis Potosí, Guanajuato, Colima, Michoacán, Querétaro e Hidalgo, una menor proporción en el norte de Chihuahua, sur de Durango, centro y sur de Nuevo León y sur de Tamaulipas; pequeñas áreas en los estados de Guerrero, Oaxaca, Veracruz, Quintana Roo, Yucatán y Baja California Sur. La amplia superficie con EF para B. repens se puede deber a que se distribuye en terrenos abiertos de pastizales, planicies y/o laderas en suelos arenosos y pedregosos, en bosques de encino, de pino, pastizales y matorrales xerófilos (Herrera et al., 2008), condiciones que presenta gran parte del territorio mexicano. Los resultados de esta investigación pronostican también que el cambio climático provocará modificaciones en la distribución de las áreas con EF, de forma que se presentarán áreas de expansión, constricción o mantenimiento de distribución de las especies bajo estudio (Tabla 2), en algunas zonas del país (Figura 2). Esto concuerda con lo mencionado por Hu et al. (2015), donde refiere que las tasas del cambio climático afectarán dramáticamente los grupos de comunidades y la biodiversidad en las zonas tropicales de la tierra.
Tabla 2 Área con aptitud ambiental para Bouteloua gracilis y B. repens con el clima de referencia y el cambio relativo (%) con cuatro escenarios de cambio climático en México.
| Climate scenarios |
B. gracilis (km2) |
% |
B. repens (km2) |
% |
|---|---|---|---|---|
| 1960-1990 | 385,305 | 100.0 | 714,837 | 100.0 |
| 4.5/2050 | 420,407 | 109.1 | 832,732 | 116.5 |
| 4.5/2070 | 414,051 | 107.5 | 829,877 | 116.1 |
| 8.5/2050 | 419,214 | 108.8 | 828,168 | 115.9 |
| 8.5/2070 | 409,327 | 106.2 | 810,655 | 113.4 |
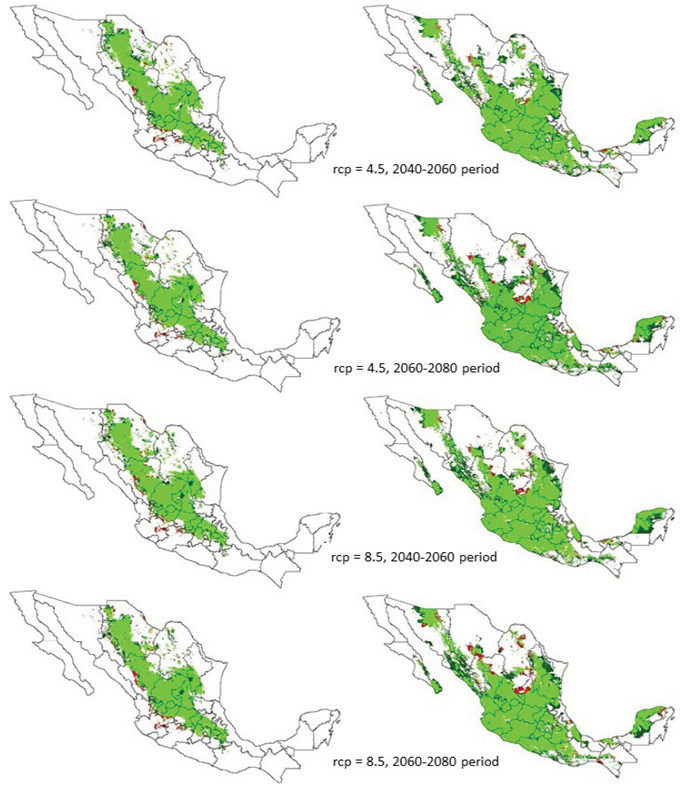
Figura 2 Área con aptitud ambiental para Bouteloua gracilis (lado izquierdo) y B. repens (lado derecho) en México debido al efecto del cambio climático con los rcp4.5 y 8.5, en los períodos 2050 y 2070. El área nueva se muestra en verde oscuro y las áreas perdidas en color naranja.
El incremento en el área con EF para B. gracilis y B. repens podría ser explicado por que ambas especies tienen ruta fotosintética C4 (Ogle, 2003; Giraldo-Cañas, 2010), que les otorga ventaja para prosperar y predominar en los ecosistemas de pastizales debido a que tienen eficiencia entre 1.3 y 4.0 veces mayor en el uso de agua y en el uso de nitrógeno, con respecto a las plantas C3 (Ghanoum et al., 2011; Wu et al., 2012; Polley et al., 2014).
Variables ambientales que en mayor medida contribuyeron a explicar la distribución de B. gracilis y B. repens.
Los resultados de esta investigación reflejaron que las variables ambientales que contribuyen a explicar la distribución de B. gracilis son: la oscilación térmica de noviembre a abril, la precipitación del mes más húmedo y el fotoperiodo de octubre a abril. Para B. repens, la precipitación del mes más húmedo y el fotoperiodo de noviembre a abril fueron las variables ambientales que más contribuyeron en la distribución de la especie.
Además, la prueba Jackknife del modelo Maxent analiza la importancia individual de cada variable, lo que contribuye a identificar las variables que tienen efectos importantes cuando se excluye el efecto de otras variables (Elith et al., 2011); de esta forma, para Bouteloua gracilis la contribución individual más importante fueron la temperatura mínima en los meses de mayo a octubre y la temperatura mínima anual (Figura 3, parte superior), con un efecto favorable con temperaturas entre 7 y 13 C, efecto menos favorable entre 3 y 7 °C y claramente desfavorables para la especie por arriba de 13 °C. El fotoperiodo fue la principal variable ambiental para Bouteloua repens (Figura 3, parte inferior) con mayor influencia en el periodo de noviembre a abril (Figura 3, parte de abajo) con un periodo de 11 horas de luz en los meses fríos y poco lluviosos del año, seguido del índice de aridez/humedad anual entre 0.3 y 0.7, que corresponde a las categorías semiárida y subhúmeda-seca, de acuerdo con la clasificación de UNEP (1997).

Figura 3 Principales variables ambientales que explican la presencia de Bouteloua gracilis (figuras superiores) y Bouteloua repens (figuras inferiores) en el clima de referencia. La línea roja muestra la zona de respuesta a la variable y la zona azul muestra la desviación estándar.
Es importante resaltar que los resultados de esta investigación se refieren a la distribución potencial de B. gracilis y B. repens en relación con las variables ambientales, pero sin tomar en cuenta factores ecológicos de la comunidad que también serán modificados por el cambio climático. Al alterar los patrones estacionales de lluvia pueden tener efectos directos en la composición y fenología de las plantas, así como efectos significativos indirectos de las interacciones entre plantas y competencia por los recursos disponibles (Prevey & Seastedt, 2015).
Conclusiones
El análisis del cambio climático indica que habrá un aumento en la temperatura media anual de 2.8 °C y 5.0 °C en relación con el clima de referencia, y una disminución acumulada anual de la precipitación entre 3.1 % y 14.4 % en los escenarios futuros; estos cambios se presentarán principalmente en las zonas con precipitación anual menor a 600 mm en el clima de referencia.
El área con aptitud ambiental para Bouteloua gracilis se incrementará entre 6.2 % y 9.1 % y para Bouteloua repens será entre 13.4 % y 16.5 % en el 2050 y 2070 respectivamente.
Los descriptores ecológicos que de manera conjunta contribuyeron en mayor medida a explicar los cambios en las áreas con aptitud ambiental fueron la oscilación térmica, la precipitación del mes más húmedo (septiembre), el fotoperiodo y la temperatura mínima del periodo más húmedo.
REFERENCIAS
Beltrán, L. S., García, D. C. A., Hernández, A. J. A., Loredo, O. C., Urrutia, M. J., González, E. L. A. and Gámez, V. H. G. (2010). Navajita Cecilia Bouteloua gracilis H.B.K. (Lag.). Nueva variedad de pasto para zonas áridas y semiáridas. Revista Mexicana de Ciencias Pecuarias 1(2): 127-130. http://www.scielo.org.mx/pdf/rmcp/v1n2/v1n2a4.pdf [ Links ]
Bocksberger, G., Schnitzler, J., Chatelain, C., Daget, P., Janssen, T., Schmidt, M., Thiombiano, A. and Zizka, G. (2016). Climate and the distribution of grasses in West Africa. Journal of Vegetation Science 27: 306-317. https://doi.org/10.1111/jvs.12360 [ Links ]
Cortés, O. A., Herrera, A. Y., Herrera, C. J. and Hernández, V. D. (2013). Potential distribution of three native and one introduced grass species in semiarid highlands of Mexico using GIS techniques. Journal of Botanical Research Institute of Texas 7(1): 441-451. https://www.jstor.org/stable/24621094 [ Links ]
Dávila, P., Mejia-Saulés, M. T., Soriano-Martínez, A. M. and Herrera-Arrieta, Y. (2018). Conocimiento taxonómico de la familia Poaceae en México. Botanical Sciences 96(3): 462-514. https://doi.org/10.17129/botsci.1894 [ Links ]
Delgado-Balbuena, J., Arredondo, J. T., Loescher, H. W., Huber-Sannwald, E., Chavez-Aguilar, G., Luna-Luna, M. and Barretero-Hernandez, R. (2013). Differences in plant cover and species composition of semiarid grassland communities of central Mexico and its effects on net ecosystem exchange. Biogeosciences 10: 4673-4690. https://doi.org/10.5194/bg-10-4673-2013 [ Links ]
Durán, P. N., Ruíz, C. J. A., González, E. D. R. and Ramírez, O. G. (2014). Impact of climate change on grasses cultivation potential of three altitudinal strata-agricultural lands of Mexico. African Journal of Agricultural Research 9: 1396-1406. https://doi.org/10.5897/AJAR2013.8121 [ Links ]
Durán, P. N., Ruiz, C. J. A., González, E. D. R., Mena, M. S. and Orozco, R. G. (2016). Cambio climático y su impacto sobre la aptitud ambiental y distribución geográfica de Salvia hispánica en México. Interciencia 41(6): 407-413. https://www.redalyc.org/articulo.oa?id=33945816006 [ Links ]
Eastman, J. R. (2016). Idrisi Selva, Guía para SIG y procesamiento de imágenes. Manual versión 17. Clark Labs, Clark University. Obtenido el 15 de octubre de 2017 en Obtenido el 15 de octubre de 2017 en https://clarklabs.org/wp-content/uploads/2016/10/IDRISISelva-Spanish-Manual.pdf [ Links ]
Edwards, E. J., & Smith, A. (2010). Philogenetic analysis reveal the shady history of C4 grasses. Proceedings of the National Academy of Sciences (PNAS) 107: 2532- 2537. https://doi.org/10.1073/pnas.0909672107 [ Links ]
Elith, J., Phillips, S. J., Hastie, T., Dudik, M., Chee, Y. E. and Yates, C. J. (2011). A statistical explanation of MaxEnt for ecologists. Diversity and Distributions 17: 43- 57. https://doi.org/10.1111/j.1472-4642.2010.00725.x [ Links ]
Fick, S. E. & Hijmans, R. J. (2017). Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. International Journal of Climatology 37: 4302-4315. https://doi.org/10.1002/joc.5086 [ Links ]
Franklin, J., Davis, F. W., Ikegami, M., Syphard, A. D., Flint, L. E., Flint, A. L. and Hannah, L. (2013). Modeling plant species distributions under future climates: how fine scale do climate projections need to be? Global Change Biology 19: 473-483. https://doi.org/10.1111/gcb.12051 [ Links ]
Galindo, L. M., Reyes, O. and Alatorre, J. E. (2015). Climate change, irrigation and agricultural activities in Mexico: A Ricardian analysis with panel data. Journal of Development and Agricultural Economics 7(7): 262-273. https://doi.org/10.5897/JDAE2015.0650 [ Links ]
Ghannoum, O., Evans, J. R. and Caemmerer, S. (2011). Chapter 8 Nitrogen and water use efficiency of C4 plants. In: Raghavendra, A., Sage, R (eds). C4 photosynthesis and related CO2 concentrating mechanisms. Dordrecht, the Netherlands. Springer Publishers. https://doi.org/10.1007/978-90-481-9407-0_8 [ Links ]
Global Biodiversity Information Facility [GBIF]. (2017). GBIF Occurrence Download. Bouteloua curtipendula (Michx.), Bouteloua gracilis (Kunth) Lag. Ex Griffiths, Bouteloua repens (Kunt) Scribu. And Merr. [Last check 11th February 2017] [Last check 11th February 2017] https://www.gbif.org/es/ [ Links ]
Giraldo-Cañas, D. (2010). Distribución e invasión de gramíneas C3 y C4 en un ambiente altitudinal de los Andes de Colombia. Caldasia 32(1): 65-86. https://revistas.unal.edu.co/index.php/cal/article/view/36194/37660 [ Links ]
Giuliani, A. L., Kelly, E. F. and Knapp, A. K. (2014). Geographic variation in growth and phenology of two dominant central US grasses: consequences for climate change. Journal of Plant Ecology 7(3): 211-221. https://doi.org/10.1093/jpe/rtt036 [ Links ]
Hernández, R. J., Reynoso, S. R., Hernández, R. A., García, C. X., Hernández, M. E., Cob, U. J. V. and Sumano, L. D. (2018). Distribución histórica, actual y futura de Cedrela odorata en México. Acta Botánica Mexicana 124: 117-134. https://doi.org/10.21829/abm124.2018.1305 [ Links ]
Herrera-Arrieta, Y., Peterson, P. M. and Valdes-Reyna, J. (2008). Journal of the Botanical Research Institute of Texas 2(2): 917-981. https://www.jstor.org/stable/41971742 [ Links ]
Herrera, C. J., González, G. F., Carrete, C. O. F., Naranjo, J. N., Pereda, S. M. E. and Herrera, A. Y. (2011). Composition and quality of cattle diet under extensive grazing on grasslands in Northern Mexico. Journal of Animal and Veterinary Advances 10(21): 2831-2837. https://doi.org/10.3923/ eda.2011.2831.2837 [ Links ]
Hijmans, R. J., Cameron, S. E., Parra, J. L., Jones, P. G. and Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology 25: 1965-1978. https://doi.org/10.1002/joc.1276 [ Links ]
Hu, G. X., Jin, Y., Wang, R. X., Mao, F. J. and Li, Y. (2015). Predicting impacts of future climate change on the distribution of the widespread conifer Platycladus orientalis. PloS One, 1, 15. https://doi.org/10.1371/journal.pone.0132326 [ Links ]
Instituto Nacional de Estadística y Geografía [INEGI]. (2009). Guía para interpretación cartográfica: Uso de suelo-vegetación Serie III. D. F. Instituto Nacional de Estadística y Geografía, México. [ Links ]
Liu, C., Newell, G. and White, M. (2016). On the selection of thresholds for predicting species occurrence with presence-only data. Ecology and Evolution 6(1): 337-348. https://doi.org/10.1002/ece3.1878 [ Links ]
Martinson, E. J., Eddy, Z. B., Commerford, J. L., Blevins, E., Rolfsmeier, S. J. and McLauchlan, K. K. (2011). Biogeographic distributions of selected North American grassland plant species. Physical Geography 2011;32(6):583-602. https://doi.org/10.2747/0272-3646.32.6.583 [ Links ]
Meneses, R. I., Larrea-Alcázar, D. M., Beck, S. G. and Espinoza, S. (2014). Modelando patrones geográficos de distribución de gramíneas (Poaceae) en Bolivia: Implicaciones para su conservación. Ecología en Bolivia 49(1): 3-19. http://www.scielo.org.bo/pdf/reb/v49n1/v49n1_a02.pdf [ Links ]
Morales, N. C., Madrid, P. L., Melgoza, C. A., Martínez, S. M., Arévalo, G. S., Rascón, C. Q. and Jurado, G. P. (2009). Análisis morfológico de la diversidad del pasto navajita [Bouteloua gracilis (Willd. Ex Kunth) Lag. Ex Steud.], en Chihuahua, México. Técnica Pecuaria en México 47(3): 245-256. https://www.redalyc.org/articulo.oa?id=61312111002 [ Links ]
Norris, D. (2014). Model thresholds are more important than presence location type: Understanding the distribution of lowland tapir (Tapirus terrestris) in a continuous Atlantic forest of southeast Brazil. Tropical Conservation Science 7(3): 529-547. https://doi.org/10.1177/194008291400700311 [ Links ]
Ogle, K. (2003). Implications of interveinal distance for quantum yield in C4 grasses: a modeling and meta-analysis. Oecologia 136: 532-542. https://doi.org/10.1007/s00442-003- 1308-2 [ Links ]
Parolo, G., Rossi, G. and Ferrarini, A. (2008). Toward improved species niche modelling: Arnica montana in the Alps as a case study. Journal of Applied Ecology 45(5): 1410-1418. https://doi.org/10.1111/j.1365-2664.2008.01516.x [ Links ]
Pérez-Ramos, I. M., Roumet, C., Cruz, P., Blanchaird, A., Autran, P. and Garnier, E. (2012). Evidence for “a plant community economics spectrum” driven by nutrient and water limitations in a Mediterranean rangeland on Southern France. Journal of Ecology 100: 1315-1327. https://doi.org/10.1111/1365-2745.12000 [ Links ]
Phillips, S. J., Dudik, M., Elith, J., Graham, C. H., Lehmann, A., Leathwick, J. and Ferrier, S. (2009). Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecology Applications 19: 181-197. https://www.jstor.org/stable/27645958 [ Links ]
Phillips, S. J. & Dudik, M. (2008). Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31: 161-175. https://doi.org/10.1111/j.0906-7590.2008.5203.x [ Links ]
Polley, H. W., Derner, J. D., Jackson, R. B., Wilsey, B. J. and Fay, P. A. (2014). Impacts of climate change drivers on C4 grassland productivity: scaling driver effects through the plant community. Journal of Experimental Botany 65: 3415-3424. https://doi.org/10.1093/jxb/eru009 [ Links ]
Prevey, J. S. & Seastedt, T. R. (2015). Effects of precipitation change and neighboring plants on population dynamics of Bromus tectorum. Oecologia 179(3): 765-775. https://doi.org/10.1007/s00442-015-3398-z [ Links ]
Reeves, M. C., Moreno, A. L., Bagne, K. E. and Running, S. W. (2014). Estimating climate change effects on net primary production of rangelands in the United States. Climate Change 126: 429-442. https://doi.org/10.1007/s10584-014-1235-8 [ Links ]
Rodríguez-Maturino, A., Martínez-Guerrero, J. H., Chairez-Hernández, I., Pereda- Solís, M. E., Villarreal-Guerrero, F., Renteria-Villalobos, M. and Pinedo-Álvarez, A. (2017). Mapping land cover and estimating the grassland structure in a priority area of the Chihuahuan Desert. Land 6, 70. https://doi.org/10.3390/land6040070 [ Links ]
Ruiz, C. J. A., Medina, G. G., Ramírez, D. J. L., Flores, L. H. E., Ramírez, O. G., Manríquez, O. J. D., Zarazúa, V. P., González, E. D. R., Díaz, P. G. and De la Mora, O. C. (2011). Cambio climático y sus implicaciones en cinco zonas productivas de maíz en México. Revista Mexicana de Ciencias Agrícolas Publ. Esp. 2: 309-323. http://www.scielo.org.mx/pdf/remexca/v2nspe2/v2spe2a11.pdf [ Links ]
Sage, R. F. & Zhu, X. G. (2011). Exploiting the engine C4 photosynthesis. Journal of Experimental Botany 62: 2989-3000. https://doi.org/10.1093/jxb/err179 [ Links ]
UNEP. (1997). World Atlas of Desertification, 2nd ed. United Nations Environment Programme. Oxford University Press, England. [ Links ]
Van Vuuren, D. P., Edmonds, J., Kainuma, M., Riahi, K., Thomson, A., Hibbard, K., Hurt, G., Kram, T., Krey, V., Jean-Francois, L., Masui, T., Meinshausen, M., Nakicenovic, N., Smith, J. S. and Rose, S. K. (2011). The representative concentration pathways: and overview. Climatic Change 109: 5-31. https://doi.org/10.1007/s10584-011-0148-z [ Links ]
Wu, Z., Dijkstra, P., Koch, G. W. and Hungate, B. A. (2012). Biogeochemical and ecological feedbacks in grasslands responses to warming. Natural Climate Change 2: 458-461. https://doi.org/10.1038/NCLIMATE1486 [ Links ]
Zarazúa-Villaseñor, P., Ruíz-Corral, J. A., González-Eguiarte, D. R., Flores-López, H. E. and Ron-Parra, J. (2011). Impacto del cambio climático sobre la agroclimatología del maíz en Ciénega de Chapala, Jalisco. Revista Mexicana de Ciencias Agrícolas Publ. Esp. 2: 351-363. http://www.scielo.org.mx/pdf/remexca/v2nspe2/v2spe2a14.pdf [ Links ]
Como citar este artículo: Martínez Sifuentes, J. A., Duran Puga, N., Ruiz Corral, J. A., G´nzález Eguiarte, D. R., Mena Munguía, S. (2020). Climate change impact on areas with environmental aptitude for Bouteloua gracilis and Bouteloua repens in Mexico Revista Bio Ciencias 7, e720. doi: https://doi.org/10.15741/revbio.07.e720
Recibido: 05 de Abril de 2019; Aprobado: 11 de Marzo de 2020
 This is an open-access article distributed under the terms of the Creative Commons Attribution License
This is an open-access article distributed under the terms of the Creative Commons Attribution License
