











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The phytochemical content of Annona muricata L. (A. muricata), is related with its pharmacological and toxicological evaluation; therefore, the measuring of antioxidant, antibacterial, antifungal and toxic properties from pulp give to us, its potential as functional, nutraceutical or toxicological food. Several antioxidant screenings have been conducted with A. muricata crude extracts from pulp, leaf and seed. Coria-Téllez et al. (2018) compiled studies on the antioxidant activity of A. muricata concluding that its antioxidants have gained interest due to their protective effect against oxygenderived free radicals involved in the development of many diseases.
On the other hand, it has been reported that A. muricata crude extracts from leaves, peel, seed and stem, have antibacterial activity against gram-positive and gramnegative bacteria. Different kind of bacteria such as: B. subtilis, E. coli, S. aureus, M. tuberculosis, S. Thyphimurium and K. pneumonia have been inhibited by A. muricata extracts (Solomon-Wisdom et al., 2014; Radji et al., 2015; Roger et al., 2015; Coria-Téllez et al., 2018). Antimicrobial bioactivity of A. muricata extracts is attributed to flavonoids, steroids and alkaloids. The mechanism of action is probably due to a synergism of these compounds (Radji et al., 2015). Microbial infections produce approximately 15 million deaths per year worldwide, with 70 % of deaths caused by bacterial infections (Essama et al., 2016). Unfortunately, some bacteria have developed resistance to antibiotics (extrinsic resistance) (Sanchez-Maldonado et al., 2017). Therefore, the study of new antimicrobial agents can be address to global public health threats due to the high resistance of pathogens to antimicrobials and the lack of development of new antimicrobials (Reis et al., 2015). In the same way, fungal infections have been increasing in the last years, due to the resistance of phytopathogens to fungicides, which has a significant impact on the economy due to the post-harvest losses caused by various fungi (Yang & Jiang, 2015). In the last decade, there is a great interest in the study of natural compounds as substitutes for synthetic fungicides and a large number of in vitro studies has demonstrated that the antifungal effects of plant extracts reduce the incidence of post-harvest pathogens (Ramos-Guerrero et al., 2018). The scientific studies on the antifungal effect of A. muricata extracts are scarce; however, Bautista-Baños et al. (2000a, 2000b, 2002) reported that A. cherimola, A. muricata and A. reticulata crude extracts from leaf and stem, had antifungal activity in vitro against Colletotrichum gloeosporioides and Rhizopus stolonifer. The authors concluded that the bioactive compounds present in Annonaceae crude extracts had a fungicidal potential.
It has been reported that aqueous extracts and isolated acetogenins from leaves or soursop pulp caused neurodegeneration or atypical Parkinsonism (Potts et al., 2012). However, Arthur et al. (2011) reported that doses greater than 5 mg/g of aqueous extract might cause kidney damage, unlike the 1mg/g dose that showed hypoglycemic and hyperlipidemia properties. The most toxic extracts that have been reported are methanol extracts of pericarp, fruit pulp or seed (Boyom et al., 2011). Therefore, more studies on the toxic effect in soursop pulp are required.
The biological activities reported in A. muricata are using crude extracts from leaf, stem, peel and seed. Therefore, the objective of this work was to evaluate the antioxidant, bactericidal, fungicidal and toxic effects of fractioned extracts obtained from fresh soursop pulp and pulp stored at different storage conditions.
Materials and Methods
Plant material
The soursop fruits (Annona muricata L.) were harvested in physiological maturity (light green skin, separate spines and firmness), of a uniform size, in orchards located in the community of “El Tonino” Compostela, Nayarit, Mexico. The fruits were ripened at 25 °C and 80-90% of relative humidity. The fruits were peeled and seeded and the pulp was used in the analysis as follows: fresh pulp (PF), pulp stored at -20 °C for 3 months (PC3), 6 months (PC6) and 12 months (PC12) and pulp treated at 70 °C for 1 h and 345.23 kPa under vacuum pressure (PT). All samples were freeze-dried at -50 °C and 0.12 mbar using a freeze dryer LABCONCO equipment (Model 77522020, Kansas, Misuri, USA).
Preparation of crude extracts
The lyophilized sample (300 g) from each treatment (PF, PC3, PC6, PC12 and PT) was mixed with 600 mL of chloroform; then the mixture was treated with 3 cycles (1 cycle for 1 h) at a constant frequency (42 kHz) on an ultrasonic equipment (Cole-Palmer8891, Illinois, USA). The extracts were filtered and the solvent evaporated to dryness in a Yamato rotary evaporator (RE300, Santa Clara, California, USA) (León-Fernández et al., 2017a).
Fractionation of crude extracts
The dry crude extract from each treatment (PF, PC3, PC6, PC12 and PT) was eluted through a chromatographic column (6.4 x 57.0 cm, 80 g SiO2 60 mesh), with CHCl3/MeOH (chloroform/methanol). Three fractions of 200 mL of each treatment were obtained in a ratio chloroform:methanol of 95:5, 90:10 and 85:15 v/v. Each fraction (F) was evaporated to dryness in a rotary evaporator, obtaining in total 15 fractions: PF-F1, PF-F2 and PF-F3 for PF sample; PC3-F1, PC3-F2 and PC3-F3 for PC3 sample; PC6-F1, PC6-F2 and PC6-F3 for PC6 sample; PC12-F1, PC12-F2 and PC12-F3 for PC12 sample; PT-F1, PT-F2 and PT-F3 for PT sample.
Bactericidal activity
Salmonella Typhimurium CCM 5445 (S. enterica subs enteric Bioserotipo Typhimurium), Enterobacter aerogenes CCM 7797 (E. aerogenes), Enterococcus faecalis CCM 2541 (E. faecalis) and Bacillus subtillis ATCC 663 strains were donated by the Public Health State Laboratory of Tepic Nayarit, Mexico. The bactericidal effect of the fractions was evaluated using the protocol of Bauer et al. (1966), with some modifications. The strains were re-growth into nutritive agar and incubated at 37 °C for 24 h. The bacterial suspensions were inoculated in a tube with saline-solution (0.1 %). These suspensions were compared with the turbidity pattern McFarland 0.5 % that correspond to homogenous suspension of 1.5x108 CFU/mL. Sterile disks (5 mm) were put on blood base agar. The fractions (2 mg) were re-dissolved in 1 mL of dimethyl sulfoxide (DMSO). Then, 20 μL of each fraction (2 mg/mL) were placed on a sterile disk. The same microliters of ciprofloxacin antibiotic (2 mg/mL) were used as positive control and DMSO as negative control. All the treatments were incubated at 37 °C for 24 h. The diameter of inhibitory halo (mm) was measured after the incubation time.
Antifungal activity
Rhizopus stolonifer and Colletotrichum gloeosporioides were isolated from soursop fruit and donated by the National Center for Genetic Resources (CNRG, Tepatitlán, Jalisco, México) and by The Laboratory of Biotechnology of Instituto Tecnológico de Tepic (Tepic Nayarit, Mexico), respectively. The strains were re-growth in PDA agar 28 ºC for 72 h. The fractions (5 mg) were re-dissolved in 1 mL of DMSO. After, each fraction was incorporated into Petri dishes. Mycelial inhibition was expressed as the percentage of inhibition of radial growth relative to the positive control (ketoconazole, 5 mg/mL). A disc agar (5 mm) containing the respective pathogen was placed at the center of each dish. Each of them was incubated for four days for Rhizopus stolonifer and seven days for Colletotrichum gloeosporioides at 28 ºC. At the end of incubation period, radial growth of the cultures was measured. The negative control was DMSO (1 mL). Three replicate dishes were used for each treatment (Bautista-Baños et al., 2002).
Antioxidant activity (AOX)
The 1,1’-diphenyl-2-picrylhydrazyl (DPPH) radical scavenging assay was performed using the method reported by Prior et al. (2005). The DPPH radical (5 mM) was dissolved in methanol at a concentration of 190 mM and kept in the dark. Each fraction was re-suspended methanol (1 mg/mL). Resuspended fractions (250 μL) were mixed with 1.5 mL of DPPH and as a blank was used methanol. The mixtures were incubated for 5 min and absorbance at 517 nm was measured. The results were expressed as the percentage of inhibition (%).
The 2,2-Azinobis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS) radical scavenging assay was performed using the method described by Re et al. (1999). The ABTS radical (7 mM) was dissolved in potassium persulfate (2.42 mM) and kept in the dark at room temperature for 14 h. The solution was adjusted with phosphate buffer solution 7.0 pH to an absorbance of 0.7 (± 0.02) at 734 nm. The re-suspended fractions (250 μL) were added to 1.5 mL ABTS solution, to be incubated in the dark for 7 min and as a blank was used methanol. The absorbance at 734 nm was measured and antioxidant activity was reported as the percentage of inhibition (%).
Toxicity determination
The toxicity test was performed using early stages of Artemia salina (Barbosa et al., 2009). Artemia (10 Artemia organism in each treatment) with 3 weeks of growth, were conserved and analyzed in saline water (25 mg/L). The fractions were re-suspended in 1 mL of DMSO. The Artemia were exposed to different concentrations (5, 10, 20, 25, 50, 100 and 150 μg/ mL) of each fraction, at 28 ºC in darkness. Artemia with 1 mL of DMSO was used as negative control. The toxicity was determined after 24 h of exposition. The number of survivors was counted and dead Artemia were considered whether they did not present internal or external movements during 1 min of observation. The average lethal concentration (LC50) was calculated using a PROBIT regression (Software SAS System v.9.0. North Carolina, USA).
Statistical analysis
All data were analyzed using an ANOVA and LSD means tests (a = 0.05), under a completely randomized factorial design 3x5 (three fractions x five treatments) with three replicates. The STATISTICA software was used (v.8 StatSoft, Tulsa, Oklahoma, USA). All assays and analysis were performed in triplicate.
Results and Discussion
Bactericidal activity of fractions from fresh, stored at -20 ºC and heat-treated soursop pulp is shown in Table 1. There was a significant effect (p<0.05) of the treatments in bactericidal activity. Although, the positive control had the highest antibacterial activity, a significant antibacterial activity bactericidal from fractions was observed. E. aerogenes was mostly inhibited by the PFF3 and PC6-F3 fractions, with the highest inhibitory halo of 17.55 to 20.25 mm, followed by the PC6-F1 and PT-F1 fractions (17.50 and 18.25 mm). On the other hand, S. Typhimurium was mostly inhibited with the PT-F3 with an inhibitory halo of 20 mm. Also, B. subtillis and E. faecalis treated with either of fractions had an inhibitory halo between 10.75 and 14.25 mm, without significant differences (p>0.05) among treatments.
Table 1 Bactericidal activity (mm of inhibitory halo) of fractions from soursop fresh pulp (PF-F1, PF-F2, PF-F3), pulp stored for 3 (PC3-F1, PC3-F2, PC3-F3), 6 (PC6-F1, PC6-F2, PC6-F3) and 12 (PC12-F1, PC12-F2, PC12-F3) months at -20 ºC and heat-treated pulp (PT-F1, PTF2, PT-F3) on gram positive and gram negative bacteria.
| Fraction | Diameter of inhibitory halo (mm) | |||
|---|---|---|---|---|
| Enterobacter aerogenes | Salmonella tiphymurium | Bacillus subtillis | Enterococcus faecalis | |
| PF-F1 | 8.50 ± 1.91c | 11.50 ± 1.00a | 10.75 ± 0.96a | 9.25 ± 2.75b |
| PC3-F1 | 13.25 ± 2.65a | 12.00 ± 2.94a | 11.75 ± 2.87a | 13.75 ± 1.89a |
| PC6-F1 | 17.50 ± 1.29b | 12.75 ± 2.22a | 12.00 ± 0.82a | 12.00 ± 1.41a |
| PC12-F1 | 13.25 ± 2.36a | 14.75 ± 2.50a | 10.75 ± 1.89a | 13.25 ± 1.89a |
| PT-F1 | 18.25 ± 1.50b | 14.50 ± 2.38a | 11.25 ± 1.50a | 11.00 ± 1.63b |
| Positive control | 31.44 ± 2.48c | 33.5 ± 1.15b | 19.38 ± 2.05b | 34.38 ± 1.70c |
| PF-F2 | 12.50 ± 1.73ab | 11.50 ± 2.52a | 11.25 ± 2.63a | 12.00 ± 1.41ab |
| PC3-F2 | 13.50 ± 2.38ab | 13.50 ± 1.91a | 13.50 ± 2.89a | 11.00 ± 1.41a |
| PC6-F2 | 13.25 ± 2.63ab | 13.50 ± 2.38a | 13.50 ± 2.38a | 14.00 ± 3.37ab |
| PC12-F2 | 10.50 ± 2.38a | 12.75 ± 1.26a | 13.50 ± 1.50 | 12.50 ± 2.08ab |
| PT-F2 | 12.50 ± 2.99b | 13.75 ± 0.96a | 13.75 ± 1.26a | 14.25 ± 1.26b |
| Positive control | 30.13 ± 2.28c | 34.13 ± 1.65b | 20.38 ± 3.24b | 34.63 ± 2.60b |
| PF-F3 | 17.75 ± 1.71b | 15.75 ± 1.71c | 11.50 ± 0.58a | 11.00 ± 0.82ab |
| PC3-F3 | 13.25 ± 2.22a | 12.25 ± 1.89a | 13.25 ± 2.63a | 11.00 ± 2.83ab |
| PC6-F3 | 20.25 ± 2.06b | 18.50 ± 3.11b | 13.25 ± 1.26a | 11.75 ± 2.36b |
| PC12-F3 | 14.25 ± 2.06a | 12.75 ± 0.50a | 11.38 ± 1.70a | 10.50 ± 1.29ab |
| PT-F3 | 13.50 ± 1.91a | 20.00 ± 0.01b | 12.50 ± 2.89a | 8.50 ± 0.58 a |
| Positive control | 28.25 ± 2.20c | 32.25 ± 4.44d | 11.50 ± 0.58a | 32.75 ± 1.32c |
Data are means ± standard deviation of three determinations by triplicate (n=9). Different letters in each file indicate significant statistical differences between treatments (α = 0.05). PC = Positive control.
Probably, the extracted compounds in the F1 fractions were mainly alkaloids, acetogenins, fatty acids and terpenoids due to its apolar nature (Coria-Téllez et al., 2018). Nevertheless, in F2 and F3 fractions, besides of the compounds mentioned above, polyphenols and other antioxidants such as vitamin C could be extracted when the polarity increase with the methanol (Park et al., 2010; Sulaiman et al., 2011); thus, F3 fractions had the highest antibacterial activity, but this was according to each bacteria. The gram-negative bacteria were mostly affected with F3 fractions due to the fact gram-negative bacteria have two lipid membranes and a thin cell wall of peptidoglycan, while gram-positive bacteria have only one lipid membrane, but the peptidoglycan wall is thicker, also there is presence of teichoic acids, which cause an intrinsic resistance (Chopra et al., 2001).
The bioactive compounds with microbicide activity in soursop extracts are acetogenins, alkaloids, terpenes, saponins, flavonoids and other antioxidants (Vieira et al., 2010; Raybaudi et al., 2015; Coria-Téllez et al., 2018). Although, there is not studies of the effect of soursop extracts on E. aerogenes and E. faecalis; saponins, steroids and triterpenes of other plant organic extracts are responsible to inhibit E. aerogenes (Reis et al., 2015). Moreover, Solomon-Winsdom et al. (2014) and Roger et al. (2015), reported that methanol and ethanolbased soursop extracts can inhibit S. thyphimurim and Bacillus subtilis bacteria.
The antibacterial mechanism can be related to alkaloids, these compounds have the ability to bind with DNA of microorganisms and inhibit RNA synthesis (Roger et al., 2015). Alkaloids have shown antimicrobial activity by glycosidase inhibition (Mohanty et al., 2008). On the other hand, it has been reported that flavonoids act by inhibiting both cytoplasmic membrane function and DNA synthesis, such as quercetin that binds to DNA subunits and inhibits the enzyme ATPase activity. Phenyl-phenol was reported to bind to the membrane proteins, or hydrogens of vital proteins such as microbial enzymes, to inhibit and change their functions (Radji et al., 2015). Similarly, the mechanism of action of the acetogenins is the inhibition of the cell respiratory chain (Lannuzel et al., 2003).
The storage of pulp at -20 ºC did not degraded the compounds with antibacterial activity, because fractions from PC samples had equal bactericidal activity than PF sample. The inhibition of S. Typhimurium increased with the thermal treatment of soursop pulp. The bioactive compounds most thermostable are acetogenins, alkaloids and saponins (Biesalski et al., 2007; García-Aguirre et al., 2008). It is possible that these compounds were concentrated during the thermal process of soursop pulp.
Table 2, shows that only the F3 fractions caused inhibition of mycelial growth of R. stolonifer and it was higher than the effect caused by the PC fractions. The highest inhibition of mycelial growth was observed with PF-F3 treatment (36.54 %). The time of storage at -20 ºC and thermal treatment of soursop pulp caused a decrease in the percentage of inhibition of mycelial growth in PC3-F3, PC6-F3, PC12-F3 and PT-F3 (27.50, 10.63, 10.0 and 1.19 %, respectively). Otherwise, the inhibition of mycelial growth of C. gloeosporiodes was observed by all fractions and in the same way, the time of storage at -20 ºC and thermal treated pulp decreased the antifungal activity of fractions, because PC12 and PT fractions had the lowest inhibition of mycelial growth in this pathogen. In contrast, PF-F3 fraction showed the highest inhibition of mycelial growth (59.90 %).
Table 2 Antifungal activity (% inhibition of mycelial growth) of fractions from soursop fresh pulp (PF-F1, PF-F2, PF-F3), pulp stored for 3 (PC3-F1, PC3-F2, PC3-F3), 6 (PC6-F1, PC6-F2, PC6- F3) and 12 (PC12-F1, PC12-F2, PC12-F3) months at -20 ºC and heat-treated pulp (PT-F1, PT-F2, PT-F3) on Rhizopus stolonifer and Colletotrichum gloeosporioides.
| Fractions | Percentage of inhibition % | |
|---|---|---|
| Rhizopus stolonifer | Colletotrichum gloeosporioides | |
| PF-F1 | NI | 32.29 ± 4.42c |
| PC3-F1 | NI | 22.28 ± 0.77b |
| PC6-F1 | NI | 24.48 ± 2.21b |
| PC12-F1 | NI | 14.13 ± 1.54a |
| PT-F1 | NI | 10.00 ± 2.83a |
| Positive control | NI | 16.00 ± 3.35b |
| PF-F2 | NI | 19.79 ± 4.42ab |
| PC3-F2 | NI | 24.46 ± 2.31b |
| PC6-F2 | NI | 15.48 ± 1.68ac |
| PC12-F2 | NI | 22.00 ± 2.83ab |
| PT-F2 | NI | 11.00 ± 1.41c |
| Positive control | NI | 15.67 ± 2.33b |
| PF-F3 | 36.54 ± 2.72d | 59.90 ± 0.74c |
| PC3-F3 | 27.50 ± 3.54c | 32.61 ± 3.07b |
| PC6-F3 | 10.63 ± 0.88a | 36.98 ± 3.68b |
| PC12-F3 | 10.00 ± 0.00a | 25.93 ± 0.00a |
| PT-F3 | 1.19 ± 1.68b | 21.00 ± 1.41a |
| Positive control | 11.93 ± 5.50c | 15.45 ± 1.89c |
Data are means ± standard deviation of three determinations by triplicate (n=9). Different letters in each file indicate significant statistical differences between treatments (α = 0.05). NI = Not inhibited
There is not literature available regarding the antifungal activity on the inhibition of R. stolonifer and C. gloeosporioides using soursop pulp extracts. However, studies on the effect of foliar and stem extracts of anonaceous was found. Bautista-Baños et al. (2000a and 2000b), demonstrated that leaf extracts of Annona reticulata and Annona cherimola had an inhibitory effect on Rhizopus mycelial growth; nonetheless, the sporulation was completely deterred. Bautista-Baños et al. (2002) found that leaf and stem extracts caused partial inhibition of mycelial growth, sporulation and germination on C. gloeosporioides. In the same way, the effect of three extracts (ethanolic, acetic and aqueous) of Annona squamosa pulp on Aspergillus niger was reported to have an inhibitory halo of 21, 15 and 17 mm, respectively (Vijayalakshmi & Nithiya, 2015).
The fungistatic or fungicidal effect of anonaceous extracts is related to the presence of alkaloids, saponins, polypeptides, terpenes, essential oils and polyphenols (Deressa et al., 2015; Perumal et al., 2016) and they have a wide spectrum of action (Coria-Téllez et al., 2018). Therefore, it is highly probable that polyphenols, alkaloids and acetogenins are present in the fractions due to the solvents used. Yang & Jiang (2015) reported that there is a dose-dependent in polyphenolic concentration to inhibit mycelial growth and spore germination of R. stolonifer. The polyphenols caused morphological changes on the hyphae such as irregularly swollen appearance, increasing branching, wrinkles, entwining, collapse and breakage, while in the spores treated with tea polyphenols was observed swelling of germ tube tips, exfoliation of the surface layer and disorganization of cell organelles. The authors concluded that the mechanism of action of polyphenols might be attributed to the direct damage of the mycelium and spore indirect induction of defensive enzyme activities. The mechanism of action of acetogenins is the inhibition of enzymes such as NADH-ubiquinone oxide reductase and NADHubiquinone oxidase (Morré et al., 1994). The fungi and yeasts are organisms with enzymes NADH-ubiquinone6-oxidoreductase type II, or NADH dehydrogenases. The effect of acetogenins on Saccharomyses cerervisae was evaluated, and they caused altered morphogenesis affecting the pathways related to the biosynthesis of chitin (Seo et al., 2000; Dominguez-Martínez et al., 2011). Thus, it is probably that these compounds also cause the synthesis of defensive enzymes and/or inhibition of respiratory enzymes.
No significant effect of fractions (p>0.05) was observed on the percentage of inhibition of DPPH radical values (Figure 1A). The DPPH radical is principally neutralized by the hydrogen donation of the antioxidants (Floegel et al., 2011). The high scavenging of DPPH radical is mostly associated with the concentration of hydrophilic compounds (Pérez-Jiménez et al., 2008). Due to the type and polarity of solvents used during the fractionation of soursop pulp, the extracted compounds were principally hydrophobic; therefore, differences of AOX with DPPH assay were not observed among treatments. León-Fernandez et al. (2017a, 2017b) found a low content of polyphenols, and a high qualitative presence of acetogenins in chloroform crude extracts and a fraction from soursop pulp; as well as a low AOX with DPPH assay. The low AOX by the DPPH assay (45-65 % inhibition) was reported in chloroform fraction and isolated acetogenins from Annona cornifolia seed extract (Lima et al., 2010).
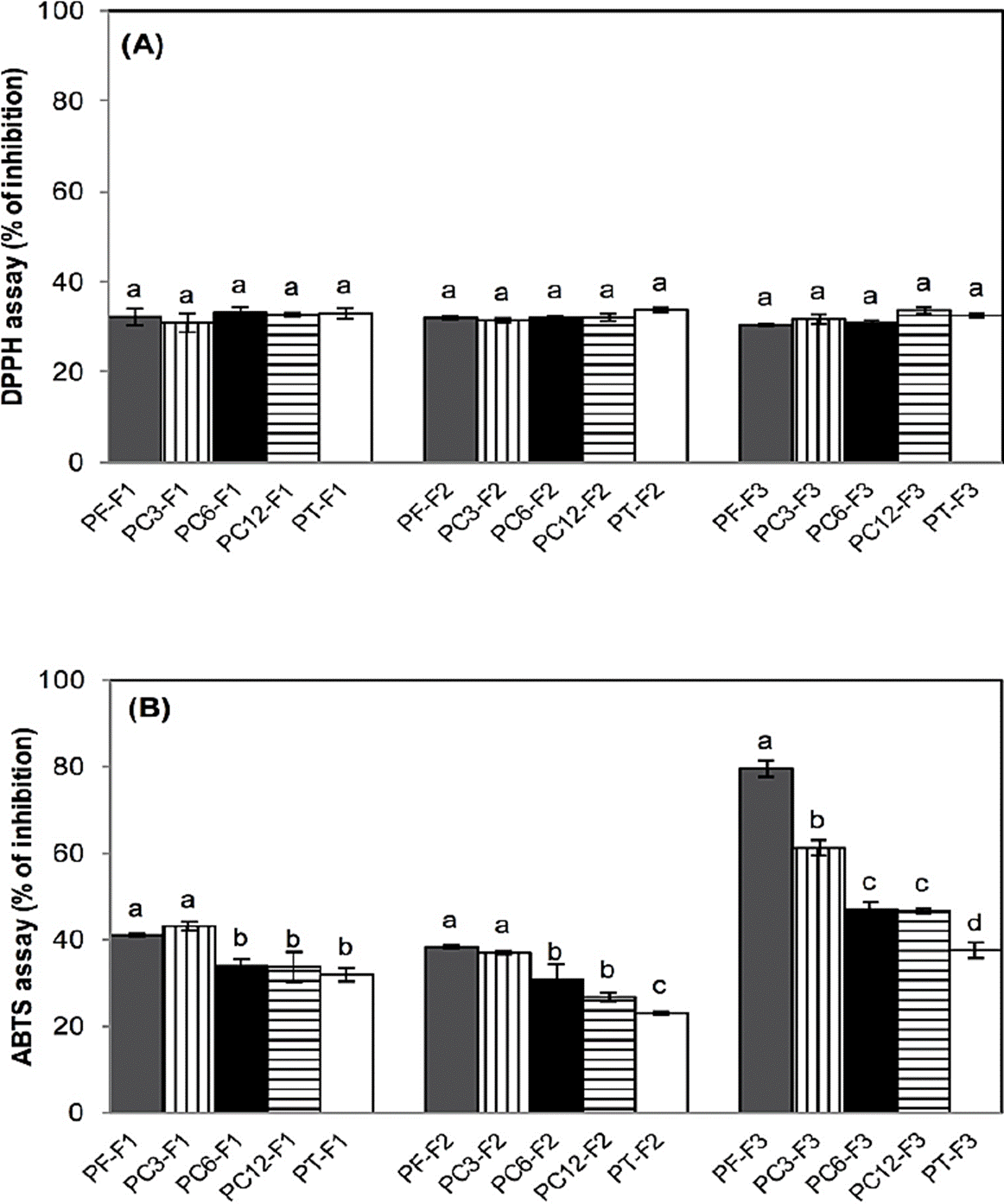
Figure 1 Antioxidant activity (% inhibition) by DPPH assay (A) and ABTS assay (B) of fractions from pulp stored for 3 (PC3-F1, PC3-F2, PC3-F3), 6 (PC6-F1, PC6-F2, PC6-F3) and 12 (PC12-F1, PC12-F2, PC12-F3) months at -20 ºC and heat-treated pulp (PT-F1, PT-F2. PT-F3). Data are means ± standard deviation of three determinations by triplicate (n=9). Different letters in each bar indicate significant statistical differences between treatments (α = 0.05).
Regarding to the ABTS assay (Figure 1B) the PF-F1 and PC3-F1 fractions had the highest AOX (40.9-43 %) than the other F1 fractions. In the F2 fractions, a similar result was observed with a 22.96 to 38.22 % of AOX; while in PF-F3 a 79.58 %, followed by PC3-F3 (61.27 %), PC6-F3 (47.09 %), PC12-F3 (46.73 %) and PT-F3 (37.47 %) was recorded. The fractions from fresh pulp might have a higher content of antioxidants than fractions from pulp stored at -20 ºC or thermal treated. The highest AOX of F3 fractions was probably caused by ratio 85:15 (v/v) chloroform: methanol; considering that in F3 fractions might exist hydrophilic and hydrophobic compounds, therefore there was a greater AOX (Lima et al., 2010).
The ABTS radical by hydrophilic and hydrophobic compounds can be neutralized (Floegel et al., 2011). León-Fernandez et al. (2017b) reported a greater antioxidant activity by ABTS assay in a chloroform crude extract and fraction from soursop pulp. The antioxidant activity in organic samples is probably due to the presence of flavonoids with low polarity, which can be extracted with chloroform, dichloromethane, diethyl ether or ethyl acetate (Sulaiman et al., 2011). It has been demonstrated that polyphenols may act via multiple mechanisms of radical-scavenging activity such as metal scavengers, transferring electrons or donating hydrogen ions. The high molecular weight phenolics (tannins) have more ability to quench free radicals such as ABTS•+ and DPPH•+ (Pérez-González et al., 2012; Paz et al., 2015); while, AOX through acetogenins may be related to the α, ß -unsaturated lactone ring moiety, similar to the in vitro mechanism of action of ascorbic acid, in which allylic hydrogens are involved. Acetogenins should act in the same way since they also possess allylic hydrogens as well as the stabilization via electron delocalization in the α ß-unsaturated lactone ring moiety. A piece of evidence to support these observations is ascorbate, which naturally occurs in plants and contains hydroxyl groups attached to a saturated lactone ring. This compound exhibited very little scavenging activity towards free radicals (Lima et al., 2010).
In the Table 3 is observed that the PF-F1, PC3-F1, PFF2 and PC-F2 fractions had the lower LC50 (2.60-3.18 μg/mL), followed by PC6-F1, PC12-F1, PT-F1, PC6-F2, PC12-F2 and PT-F2 fractions (4.57-7.04 μg/mL). The F3 fraction from fresh pulp showed the highest LC50 (32.76 μg/mL), followed of PC3-F3 (17.45 μg/mL), PC6, PC12 and PT (13.38-14.13 μg/mL) without significant differences among them (p>0.05). The storage at -20 ºC and thermal treatment of soursop pulp may affect the chemical structure of some bioactive compounds, which could cause a decrease in toxicity.
It has been reported the toxicity of aqueous and ethanolic crude extracts from Annonaceae leaf and stem. The leaf and stem aqueous extracts had a LC50 of 1000 and 865 μg/mL, respectively; while leaf and stem ethanolic extracts showed a LC50 of 831 and 200 μg/ mL, respectively (González-Esquinca et al., 2012). Vila-Nova et al. (2013), determined the toxicity of acetogenins isolated from soursop leaves (annonancinone and corossolone) on early stages of Artemia salina. They reported high activities of 7.09 and 17.05 μg/mL, concluding that the acetogenins isolated possess higher cytotoxic activity than extracts. The toxicity of fractions might be related to the presence of acetogenins due to the extraction solvents (León-Fernández et al., 2017a).
Conclusion
The present study demonstrated that some fractions from soursop pulp showed antibacterial activity and antifungal activity. In addition, the antioxidant activity and toxicity of all fractions were registered, although the results depended of each treatment. The data support that the use of extracts from soursop pulp may be considered for the development of products in herbal medicine as pre-treatment in the biological control of postharvest diseases. Nevertheless, further studies need to be performed, either aiming at the isolation of compounds in the search for new antimicrobials or antifungals or evaluating the safety and efficacy of fractions from soursop pulp for a possible pharmaceutical use.

