











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Cloud forest rats belonging to genus Nephelomys (Cricetidae: Sigmodontinae) distribute in the Andes (900 to 3000 m); they can be found in wooded paramo, montane cloud and elfin forest ecosystems (Percequillo 2003, 2015). Species of genus Nephelomys are present in South America from Colombia to Bolivia, including costal Venezuela, extending to the Cordilleras of Panama and Costa Rica in Central America (Percequillo 2003, 2015; Prado and Pecrequillo 2013). Four of the thirteen species of the genus, N. childi, N. maculiventer, N. pectoralis and N. meridiensis are known from Colombia (Percequillo 2003, 2015; Solari et al. 2013; Ramirez-Chavez et al. 2016).
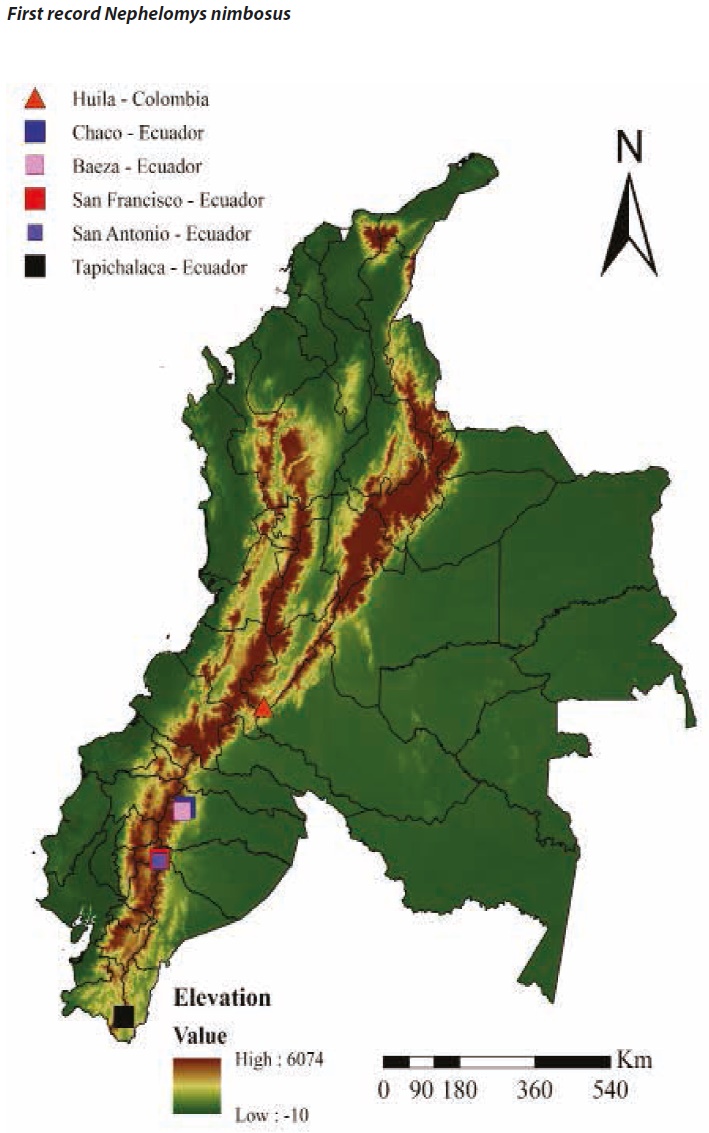
Historically the species N. nimbosus (Anthony 1926) was considered endemic to Ecuador. It was known from five localities along the eastern slope of the Andes Cordillera, specifically two locations in the Napo Province (Baeza and Chaco Oyacachi River) and two in the Tungurahua Province (San Antonio, San Francisco; Percequillo 2003, 2015). Later, a new locality record at southeastern Ecuador (Tapichalaca, Zamora Chinchipe Province) was reported (Brito et al. 2015; Figure 1). Given the scarcity of records, the natural history of N. nimbosus is poorly known. Here we present the Colombian first record for N. nimbosus.
Materials and methods
Individuals were captured between February and August of 2014 as part of the first inventory of small non flying mammals for the Natural National Park Cueva de los Guacharos (PNNCG) located in the department of Huila, Colombia (1° 36.14’N, -76° 8.13’W). Capture survey consist in linear transects of Sherman traps placed on the ground, a total of 100 traps were active for 150 days, totalizing a trapping effort of 1,5000 traps night (Bautista et al. 2014). N. nimbosus individuals were captured at two of the five sampling localities. The first one was a subandine primary forest at 2000 m; this location presents a high canopy with lianas and palms; some of the most abundant species were plants of the genus Wettinia, Hyeronima, Alchornea, Miconia, and Inga (Prada and Stevenson 2010). The second trapping locality was ‘Cueva de los Guacharos’ at 1900 m, a small cave with no vegetation where nocturnal frugivore birds known as oilbirds (Steatornis caripensis) build their nests and spend most of the daylight. The accumulation of Oilbird seed wastes on the bottom of the cave (e. g., Prestoea acuminate, Geonoma sp, Dacryodes sp, and Oenocarpus bataua) serves as a food resource for small non-flying mammals, like N. nimbosus and N. childi (Bautista et al. 2014).
The N. nimbosus specimens were deposited in the mammal collection of the Universidad Distrital Francisco Jose de Caldas (MUD) in Bogota, Colombia. Analyzed specimens (MUD 877; MUD 883; MUD 884; MUD 893) were compared with the holotype of N. nimbosus (American Museum of Natural History, AMNH 67337; Anthony 1926; Percequillo 2003), and a specimen of this species from south eastern Ecuador deposited in the Ecuadorian Museum of Natural Sciences (DMMECN 1083; Brito et al. 2015). Also, specimens were compared with individuals of N. childi (MUD 876; MUD 876; MUD 880; MUD 887; MUD 889). Body and cranial measures were taken with a gauge ± 0.1 following the definitions of Voss (1988) and Patton et al. (2000).
Results
Four individuals from a total of 198 of the first inventory of small non flying mammals of PNNCG were identified as belonging to N. nimbosus using the description and key provided in Anthony (1926) and Percequillo (2003, 2015). Some co-occurring species were Marmosops parvidens, Marmosops caucae, Heteromys anomalus, Melanomys caliginosus, Chilomys instans, Neacomys tenuipes, Rhipidomys latimanus, Microryzomys minutus, Tramsandinomys talamancae and Nephelomys childi, most of them typical of montane forest.
Captured individuals present the following characteristics that separates them from those of N. auriventer and N. childi (Figure 2). Ventral patch of self - colored white hairs in the gular region (not present in N. auriventer), ventral pelage ochraceous yellow with a golden tone (N. childi presents ventral pelage grayish or whitish washed with gray), medium body size between 122 and 140 mm (N. auriventer is larger, measuring between 140 and 173 mm), incisive foramina short range: 4.6 to 5.1 mm with lateral margins rounded (N. auriventer has long incisive foramina range: 5.6 to 6.7 mm with teardrop shape), posterolateral palatal pits small to big shallow and simple (N. childi presents posterolateral palatal pits complex small numerous and deep), interorbital region hourglass shaped, with supra-orbital margins rounded or squared (N. auriventer has an interorbital region with strongly beaded supraorbital margins, which diverge posteriorly) and a thin and delicate face (N. nimbosus has a robust and strong face; Anthony 1926; Percequillo 2003, 2015). No comparison were made with N. pectoralis because of its distinctive large and wide white ventral patches and its numerous and deep posterolateral pits placed in deep palatal fossae (Allen 1912; Weksler et al. 2006). Most of cranial and body measures of the four individuals are within the known range for the species (Percequillo 2003, 2015; Weksler et al. 2006; Brito et al. 2015; Table 1). This new record of N. nimbosus expands the known distribution of the species 760 km towards north, and constitutes the first record of the species for Colombia.

Figure 2 Dorsal (A) and ventral view (B) of the skull and (C) skin of an adult Colombian female of Nephelomys nimbosus (MUD 883). Dorsal (D) and ventral view (E) of the skull and (F) skin of the holotype of Nephelomys nimbosus (AMNH 67337).
Table 1 External and cranial measures (in millimeters) of four specimens of Nephelomys nimbosus from Colombia (MUD 877, MUD 884, MUD 893, MUD 883), Nephelomys nimbosus of Zamora Chinchipe province (DMMECN 1083; Brito et al. 2015), and the holotype of Nephelomys nimbosus (AMNH 1083) from Tungurahua province, Ecuador taken from Percequillo (2003).
| Measures (mm) | Colombia | Ecuador | ||||
|---|---|---|---|---|---|---|
| MUD 877 | MUD 884 | MUD 893 | MUD 883 | AMNH 67337 (Holotype) |
DMMECN 1083 |
|
| Total length | 284 | 292 | 302 | 242 | 300 | 289 |
| Tail length | 129 | 149 | 153 | 116 | 160 | 147 |
| Head and Body length | 155 | 143 | 149 | 126 | 140 | 142 |
| Hindfoot length | 30.8 | 32.0 | 31.6 | 30.7 | 34.0 | 33.0 |
| Ear length | 20.9 | 18.2 | 17.7 | 16.6 | - | 19.0 |
| Weight (grams) | 74.9 | 73.7 | 57.4 | 42.3 | - | 84.0 |
| Condylo insicive length | 33.02 | 30.73 | 30.37 | 27.86 | 32.04 | 30.50 |
| Superior molar series length | 6.46 | 5.54 | 5.38 | 5.61 | 5.57 | 5.50 |
| Diastema length | 10.35 | 8.69 | 9.30 | 7.77 | 9.48 | 9.10 |
| Incisive foramina length | 5.80 | 5.41 | 4.83 | 4.36 | 5.12 | 4.30 |
| Occipitonasal length | 36.61 | 34.25 | 33.65 | 30.81 | 35.33 | 34.10 |
| Molar 1 width | 1.78 | 1.72 | 1.62 | 1.78 | 1.64 | - |
| Nasal length | 14.32 | 12.38 | 13.91 | 12.69 | 13.13 | - |
| Incisive foramina width | 3.35 | 2.33 | 2.39 | 1.96 | 2.82 | - |
| Zygomatic plate width | 3.57 | 2.94 | 3.09 | 2.65 | 3.81 | 3.80 |
| Palatal bridge width | 3.48 | 3.16 | 3.98 | 3.05 | - | - |
| Inter orbital lesser width | 4.94 | 5.11 | 5.85 | 4.85 | 5.79 | 5.40 |
| Zygomatic width | 18.59 | 17.49 | 16.52 | - | 17.75 | 18.40 |
| Rostral length | 8.00 | 9.12 | 9.92 | 7.38 | - | 11.50 |
| Cranial width | 14.39 | 14.29 | 13.77 | 13.32 | 15.30 | 14.30 |
| Cranial height | 10.21 | 9.60 | 9.93 | 9.73 | 10.80 | 10.20 |
Discussion and conclusions
Colombian sigmodontines have recently suffered taxonomic changes, as well as improvements in their know distribution. Hence, the species number from Colombia has increased from 66 in 2013 (Solari et al. 2013) to 75 in 2016 (Ramirez-Chavez et al. 2016). The new records of N. nimbosus of the present study and of N. meridiensis mentioned by Percequillo (2015; previously identified as Nephelomys sp. by Solari et al. 2013 and Percequillo 2003), increase the number of species of genus from three to five for Colombia and contributes to the constant changes of the Colombian sigmodontines species list.
The discovery of N. nimbosus in Colombia is important due to the apparently restricted distribution of the species that was previously known only from five localities in Ecuador (Percequillo 2003, 2015; Prado and Pecrequillo 2013; Brito et al. 2015). The potential presence of the species extends to the northern Ecuadorian Cordillera, the Colombian Cordillera massif, and to the southern part of the Colombian eastern Cordillera. Although the Andes montane forests have been catalogued as hotspots of mammalian diversity and endemism (Myers et al. 2000; Ceballos and Ehrlich 2006), it is important to increase the sampling effort for small non-flying mammals in this poorly known region where deforestation rate is increasing alarmingly (Armenteras et al. 2011). The limited number of known individuals of N. nimbosus makes of it a rare species with a poorly known distribution (Percequillo 2003, 2015; Prado and Pecrequillo 2013). For instance, in the present study the species had an abundance of four out of a total 198 small mammals captured, with a 15,000 traps night sampling effort, during a five months continuous sampling period (Bautista et al. 2014). We suggest increasing the amount of studies in the montane forest near the borderline between Colombia and Ecuador, where the species potentially could be found. Due to these facts it is essential to increase studies of the genus Nephelomys, with the purpose of gathering information about species geographical distribution and ecology, which in turn would contribute useful material to clarify species limits and their phylogenetic relationships.

