











 text new page (beta)
text new page (beta) English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroduction
The black bear (Ursus americanus, Pallas 1780) is a species that has inhabited North America for approximately three million years, during which it has extended across a broad geographical range (Kurten and Anderson 1980). This distribution changed significantly during the Pleistocene as a result of the southward shift in the distribution pattern and extent of forests due to glaciation and post-glaciation events (Wooding and Ward 1997). This phenomenon favored the evolution of two clades - coastal and continental - within the species (Pelletier et al. 2011). Within the continental clade, the distribution and topology of the diversity of mitochondrial DNA has revealed the existence of two subclades - eastern and western - that diverged in isolation between 1.8 and 0.8 million years ago (Wooding and Ward 1997; Van Den Bussche et al. 2009). These subclades have shown disjunct distributions over a long period of time, with refuges associated with forests to the east and west of their distribution range (Wooding and Ward 1997). Contact between both subclades in areas of Texas and New Mexico is considered a recent event (Wooding and Ward 1997).
The difference between these subclades is also evident within the southern limit of the distribution range of this species. The dominant haplotypes found for black bear populations in the Trans-Pecos region, Texas, have been identified as belonging to the eastern subclade (Onorato et al. 2004), while haplotypes described for northern Sonora, in western Mexico, belong to the western subclade (Varas et al. 2010).
Currently, due to the high genetic differentiation between subclades, it has been postulated that the gene flow between the populations inhabiting Sierra Madre Oriental and Sierra Madre Occidental in Mexico is non-existent (Onorato et al. 2004). The above has been proposed due to the existence of desert environments surrounding montane habitats, which act as barriers, in addition to present human activities and presence of towns and/or cities that prevent the displacement of individuals within an array of highly transformed habitats (Atwood et al. 2011).
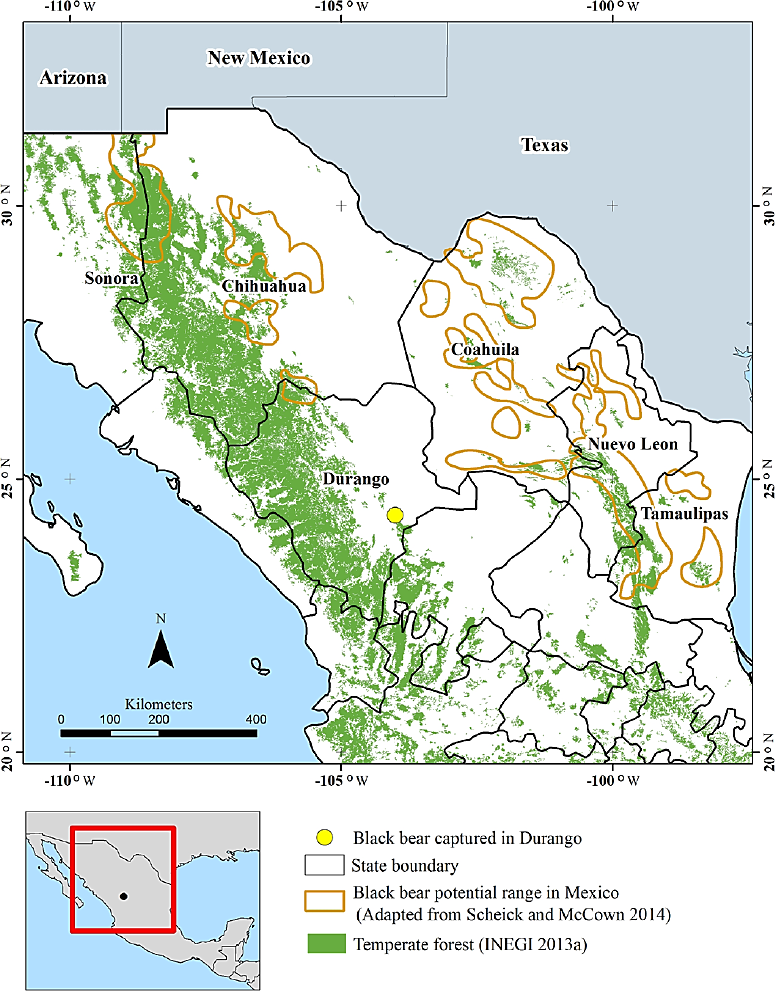
Figure 1 Record of a black bear specimen (Ursus americanus) captured in the municipality of Guadalupe Victoria, Durango, compared with the area of distribution of the species in Mexico (Scheick and McCown 2014).
In Mexico, the black bear is considered a species endangered in risk of extinction throughout most of its distribution range (DOF 2015), an area that was significantly reduced largely due to the impact of anthropogenic factors during the twentieth century. The presence of the species in the southern area of its distribution range in Mexico is characterized by a low number of records, and the precise limits of this range are still unknown (Delfín-Alfonso et al. 2011; Delfín-Alfonso et al. 2012; Scheick and McCown 2014). On April 2014, one black bear specimen was captured by local inhabitants in Felipe Carrillo Puerto, a town located in the municipality of Guadalupe Victoria, state of Durango. Durango is one of the states where there is a high uncertainty about the presence of the black bear (Delfín-Alfonso et al. 2012; Juárez-Casillas and Varas 2013); prior to this capture, there had been no recent records of the species in the area (Delfín-Alfonso et al. 2011). The presence of the specimen in this locality raise questions about its geographical origin, whether associated with the Sierra Madre Occidental or the Sierra Madre Oriental. This specimen brought the unique opportunity to investigate the possible existence of a recent connectivity event between both regions; accordingly, our aim was to determine the geographic origin of the black bear specimen by characterizing its haplotype.
Materials and Methods
Study area. Felipe Carrillo Puerto (24° 27' N, -40° 07' W) is one of the largest urban localities in the municipality of Guadalupe Victoria; it is located to the southeast of the state of Durango, with a population of 2,000 inhabitants. The town is surrounded by rainfed agriculture fields that make up 51 % of the municipality area, while natural grasslands rank second in terms of coberture (28 %), and pine and oak forests comprise less than 4% of the municipality area (INEGI 2013a). Mean annual precipitation ranges between 450 mm and 605 mm per year, and mean annual temperature, between 12.7 °C and 17.4 °C. The elevation within the municipality varies from 1,912 to 2,560 m (INEGI 2013b).
Capture and Maintenance of the Black Bear Specimen. The black bear specimen was captured on 23 April 2014 by two local inhabitants of Felipe Carrillo Puerto using ropes. Once the specimen was securely held, the local inhabitants informed the authorities concerned. On the first veterinary examination at the site of the capture, personnel from PRO-FEPA Durango and the Sahuatoba Zoo mentioned that the specimen was in good health condition (PROFEPA 2014).
The specimen captured was transferred by PROFEPA Durango to the Sahuatoba Zoo, in the city of Durango, where he was sedated by the local staff (2 mg/kg each of ketamine/xylazine). Once sedated, this specimen was identified as a young male (approximately two years old based on dental wear) and its somatic measurements were recorded (LT = 1.540, LC = 90, LP = 220, 116, W = 81.5); the specimen was in good physical condition. During the examination, samples of ear tissue, hair and feces (two replicates of each) were collected for processing at Universidad Autónoma de Queretaro; afterwards, the specimen was fitted with a radio transmitter collar.
Haplotype Characterization. For DNA extraction, 30 mg of tissue, 0.50 g from the surface of excrement, and three hairs were collected from each sample. For tissue and hair samples, the DNeasy Blood & Tissue mini kit (Qiagen Inc., Valencia, California) was used following the manufacturer's protocols. For stool samples, the QIAmp ®DNA Stool mini kit (Qiagen Inc., Valencia, California) was used following the protocol developed by Varas et al. (2010). To confirm the proper operation of the protocol and the amplification and sequencing procedure, we used a negative control (high purity water for PCR) and a positive control (previously identified black bear DNA sample).
Once the mitochondrial DNA was extracted, it was amplified by PCR. Two sequences of 418 base pairs were obtained using a portion of the cytochrome b region and D-loop of mitochondrial DNA, using the primer ADNmtCRf (CTCCACTATCAGCACCCAAAG) and its inverse ADNmtCRr (GGAGCGAGAGGTACACGT) with a volume of 15 ml, where 9 ml were green taq (PROMEGA Corp.), 0.2 ml BSA 7.5 % (Sigma-Aldrich, St. Louis, MO, USA), 0.2 ml MgCl2 (QIAGEN, Inc.), 0.3 ml of each primer, and 5 ml of DNA (Rinkevich 2012).
Initial denaturation was carried out at 95 °C for 10 minutes, followed by 40 denaturation cycles at 94 °C for 45 seconds, hybridization at 54 °C for one minute, extension at 72 °C for two minutes, final extension at 72 °C for 10 minutes, and cooling at 4°C (Short Bull et al. 2011). The resulting amplifications were subjected to electrophoresis through 1.5% agarose gel with TBE buffer (Onorato et al. 2004) and were visualized through exposure to UV rays. Amplifications were purified using the Wizard PCR Prep DNA Purification System (Promega, Madison, Wisconsin).
The amplified sequences were sent to Macrogen Sequencing Service http://dna.macrogen.com/esp for sequencing. Afterwards, sequences were reviewed and aligned in the program Sequencher 4.6 (Gene Codes Corporation, USA). To determine the haplotypes in the sequences analyzed, ADNmtCR was tested for neutrality using the program dnasp5 (Avin 2012). Finally, GenBank (NCBI http://www.ncbi.nlm.nih.gov/) was searched for potential matches with haplotypes previously described and characterized geographically (Onorato et al. 2004; Varas et al. 2010, Van Den Bussche et al. 2009; Pelletier et al. 2011), aiming to associate the origin of the black bear with either the Sierra Madre Oriental or the Sierra Madre Occidental.
Results
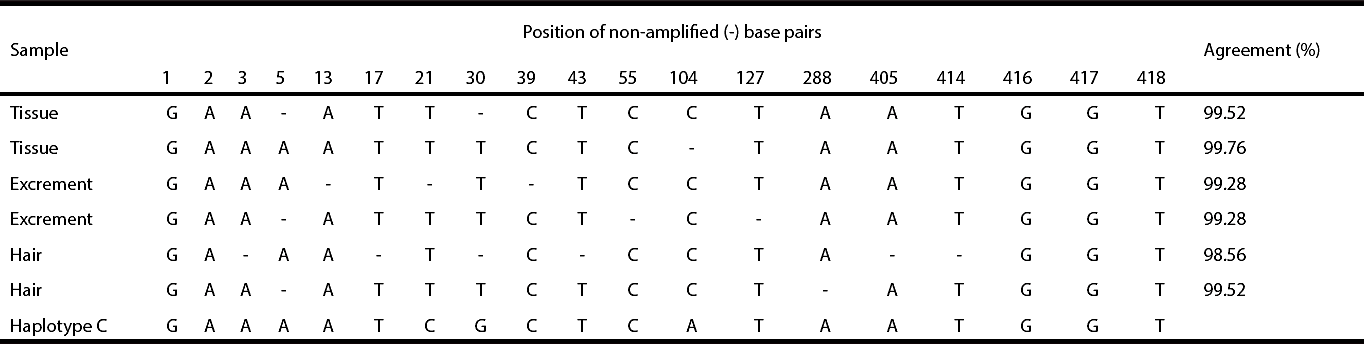
The DNA from the six samples of the black bear specimen were successfully amplified (Table 1); thus, all were used in the haplotype characterization analysis. The neutrality test yielded a single haplotype. Sequences of 418 base pairs were obtained and the number of base pairs that were not amplified per sample ranged from 1 to 6. Considering these base pairs as unknown, our samples yielded an agreement of 99.32 % ± 0.41 (std dev.) with haplotype C described by Onorato et al. (2004) (Table 1).
Table 1 Agreement of base pairs (bp) between samples of the black bear captured in Felipe Carrillo Puerto, Durango, and haplotype C described by Onorato et al. (2004). Base pairs 1-3 correspond to the start codon; bp 416-418, to the stop codon.
Based on the above findings, the black bear specimen captured in the municipality of Guadalupe Victoria, Durango, comes from the subclade inhabiting Sierra Madre Oriental.
Discussion
The bear captured in Felipe Carrillo Puerto, Durango, belongs to the eastern subclade representing the populations of Sierra Madre Oriental, since haplotype C has not been recorded in Sierra Madre Occidental (Varas et al. 2010). Haplotype C was first described in 2004 from a single specimen inhabiting the Trans-Pecos region, Texas, USA, and has been recently documented in Louisiana (Van Den Bussche et al. 2009). It is considered that haplotype B gave rise to haplotypes C and A. Haplotype B is considered the parent haplotype within the eastern subclade, being broadly distributed from Minnesota in northern United States to Mexico. Van Den Bussche et al. (2009) consider that colonization by this set of haplotypes (A, B, and C), which are characteristic of the eastern lineages, took place gradually from east to west given the absence of barriers that would restrain gene flow.
The specimen captured in Durango traveled more than 250 km from the area recognized as a part of the known distribution range of the black bear in Coahuila to reach the municipality of Guadalupe Victoria (Scheick and McCown 2014). This dispersal event by a relatively young individual strongly supports the fact that the connectivity between the Sierra Madre Occidental and the Sierra Madre Oriental is currently maintained for black bears.
Black bear males are capable of carrying out extensive dispersal events in desert environments within a mountain range. There are reports about the displacement of males covering between 45 and 282.2 km in straight line; these movements have been attributed to a fragmented environment where a suitable habitat is surrounded by grassland or arid areas (Costello et al. 2001; Hellgren et al. 2005; Liley and Walker 2015). The bear specimen reported herein followed a route similar to the one reported previously. However, this individual extensive displacement connected the two largest mountain ranges of Mexico passing through a landscape matrix dominated by desert conditions, which are considered for the species relative to temperate forests (Costello et al. 2001).
Since a single black bear individual - and hence a single haplotype - was available (Van Den Bussche et al. 2009), the presence of more individuals in Sierra Madre Occidental coming from Sierra Madre Oriental is currently unknown. Although we cannot be certain that the specimen captured comes from the wild, it showed some features that so indicate. On the one hand, the specimen lacked the calluses on pads and joints that are typical in animals that have spent time in captivity. However, the behavior of the specimen after its release was consistent with the one of a wild animal. The specimen was released by Durango PROFEPA and CONANP staff in La Michilía Biosphere Reserve less than a week after its capture. From the time of release, its displacements were recorded through satellite location over a period of 19 months; it traveled the desert between the two mountain ranges heading to Sierra Madre Oriental (unpublished data).
The genetic diversity, as well as the frequency of dispersal events between the two Sierras Madres, the time and distance between displacement events, the likely dispersal routes and the presence of patches of suitable habitats, are all factors that should be assessed to understand the current dispersal and gene-flow patterns between the subspecies of bears in Mexico.

