











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
Andean bears (Tremarctos ornatus) are large carnivores (1.5 - 2.1 m total length; 70 - 130 kg body weight), that inhabit paramo, puna and Andean forests between 250 and 4,750 m along the Andes mountain range (Peyton 1999). This species ranges from Colombia through Ecuador, Peru and Bolivia (Peyton 1980, Rodríguez et al. 2003), to northern Argentina (Del Moral and Bracho 2009). The mountainous habitat that this species inhabits makes the study of its natural history difficult (Jones 2010).
A number of research studies on the Andean bear have been conducted throughout its distribution range (García-Rangel 2012, Reyes-Amaya 2015). However, key aspects of their natural history such as population dynamics, habitat use and ethology in the wilderness (e. g., courtship and reproduction) have been scarcely described (Reyes-Amaya 2015). The large area requirements (home range: 59 km2 for males, 15 km2 for females) and heterogeneity of the Andean bear habitat, along with their vulnerability to land use by humans and prolonged parental care, all increase the extinction risk for this species (Stern 1998; Cardillo et al. 2004; Castellanos 2011). Therefore, it is crucial to gain a better understanding of the Andean bear biology for formulating and implementing suitable strategies for their conservation and management (Rodríguez et al. 2003).
Mammalian identification based on natural marks of the specimens (e. g., external coloration patterns) has been extensively used to estimate population patterns from trap camera images (Foster and Harmsen 2012). The Andean bear coat varies from black to dark brown, usually with white markings or other colour variants on the snout, neck, chest and around the eyes (although some individuals do not show any markings), which show distinct variations between specimens (Roth 1964; Peyton 1999). Previous studies have used camera traps to address population aspects of the Andean bear (e. g., catch rates, population density, abundance) and some of those have evaluated the effectiveness of this technique for identifying specimens of this species (Ríos-Uzeda et al. 2007; Zug 2009; Van Horn et al. 2014). However, due to the recent implementation of video format in camera traps, no information is yet available on the comparative efficiency of using either photographs or videos in camera traps for identifying Andean bear specimens. In this study we examine photographic and video records of Andean bears acquired by camera traps in a wild population to compare the efficiency of the two visual file formats for identifying individuals.
Materials and methods
The study area includes parts of the Gachetá and Junín municipalities in the Cundinamarca Department. These municipalities are located on the western slope of the eastern cordillera of the Colombian Andes, under the jurisdiction of the Corporación autónoma Regional del Guavio (CORPOGUAVIO), the State environmental authority in this part of the country. The mountainous topography of the study area comprises moderate to steep slopes (undulating to rugged terrain) in colluvial landscapes and denudative structural mountains, covered by fragmented Andean forest, sub-paramo and paramo ecosystems, between 2,500 and 3,200 m (Cleef 1978). Mean annual temperatures range between 5.7 and 16 ° C (Rangel-Ch 2000; Vargas and Pedraza 2004), with mean annual precipitation of up to 1,861 mm and humidity up to 90 %, in a unimodal seasonal rainfall regime; the rainy season runs from April to October (with a peak in June and July) and the dry season from November through March (minimum rainfall between December and January; INDERENA 1986, Aguilar and Rangel-Ch 1996).
Natural trails made by Andean bears within the forest were located based on traces of habitat use by this species (e. g., evidence of climbing and presence of camaretas on trees, feeders and feces) and the knowledge of community research monitors (local residents). We selected those trails showing the most signs of recent use by the species (≤ 6 months, see Rodríguez 2006), in order to obtain the largest possible number of visual records. Trap cameras were located at sampling stations along the trails within the forest (Zug 2009; Jones 2010). Each sampling station included a single camera attached to a tree trunk at 0.6 m height, with the bait (honey or panela) placed 2 m from it at 1.5 m height, to encourage the specimens to raise on their hind limbs to reach the bait.
A total of 18 camera traps of different brands and models, but with similar photographic qualities or the same video quality, were used (Table 1). The cameras were active for two years (November 2011 to November 2013), programmed to operate 24 hours a day, taking a sequence of photographs or videos every time they were activated by motion in front of them (recording event). Visual files in which an Andean bear was not recorded were excluded from further analysis. Photograph sequences were taken with 1 second interval between shots for as long as the subject was in front of the camera. For video recordings, 60-second videos were shot with 1 second interval between them for as long as the subject was in front of the camera. The cameras were checked every 15 days to verify their proper functioning, download the visual files acquired and replace batteries. Visual files acquired were tagged with the recording date and time.
Table 1 Brands, models and quality of photographs and videos acquired by camera traps used in the study. Photograph quality is measured in megapixels (Mp), video quality is measured in pixels (p). Bushnell Trophy Cam ™ HD HD 119476 (Bushnell), Reconyx Hyper Fire Infrared Digital Game Camera HC600 (Reconyx), Wildview® SCT-TGL5IR (Wildview).

For each recording event, an attempt was made to identify the specimen within each visual file acquired. Specimens were identified based on the presence of four key external morphological features (attributes useful for specimen identification): presence, shape and colour of facial markings; presence, shape and colour of pectoral markings; estimated body size; and sex, as observed from three different viewpoints, as described by Zug (2009) (Figure 1). The body size of the specimen was estimated by comparison with a reference object of known size located within the visual field of the camera; sex was determined either based on the specimen’s genitalia (when visible) or the presence of other reproductive features (e. g., turgid mammary glands, presence of cubs). A visual file (photograph or video) was scored as “good” for identification of the specimen when at least three of the four key features could be clearly observed in it. A visual file was scored “bad” when only two or fewer identification features could be observed. Facial and pectoral markings are perhaps the most useful features for identifying specimens of this species (Roth 1964; Peyton 1999). However, not all the specimens show both kinds of markings, in which case the identification must be complemented withother features (e.g., estimated body size and sex). The use of complementary features allows discriminating between specimens even with basis on one single marking, which might appear rather similar in different specimens as it is not always possible to fully observe the entire marking in visual files (photograph or video). Additionally, some bear specimens do not display any marking, which makes their identification impossible (D. Rodríguez com. Pers.).
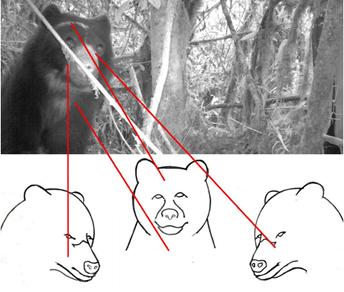
Figure 1 Scheme showing how facial features were recorded for the identification of Andean bear specimens using camera traps.
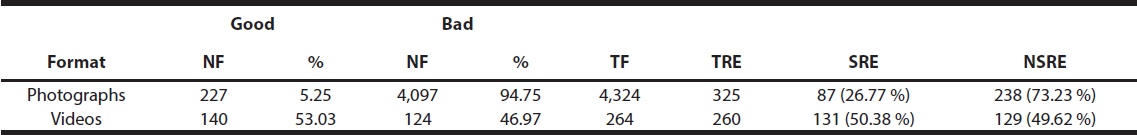
The percentage of recording events that included good visual files, and thus allowed the correct identification of the specimen recorded (successful recording events), as well as the percentage of recording events that did not include any good visual files, and thus made the specimen identification impossible (unsuccessful recording events), were separately calculated for each photograph/video data set acquired during the entire sampling period (Table 2). Similarly, the percentage of good and bad visual files acquired over the entire sampling period was separately calculated for each photograph/video data set (Table 2).
Table 2 Visual files and recording events acquired in the study, their quality and capacity to allow identification of Andean bear specimens. Number of files (NF), total number of files (TF), total number of recording events (TRE), number of successful events (SRE), and number of non-successful events (NSRE)
In addition, by considering each recording event as an independent event, we tested for statistically significant differences in the number of good and bad visual files acquired every time a camera was activated by the motion of an Andean bear in front of it. Comparisons were made within the photograph and video data sets and between the two sets. As the number of visual files acquired in each recording event was variable, prior to analysis the number of good and bad visual files was expressed as a percentage of the total number of files acquired in the recording event. As the data showed a non -normal distribution, the non-parametric Mann-Whitney test was used for the analyses.
Results
A total of 585 recording events were obtained, comprising a total of 4,588 visual files (4,324 photographs in 325 recording events and 264 videos in 260 recording events, Table 2). Of the 325 recording events that used photographs, 26.77% (87) were successful, allowing the identification of the specimen recorded; 49.62 % (129) of the recording events that used videos were successful (Table 2).
Of the total number (4,324) of photographs acquired, 5.25 % (227) were rated as good and 94.75 % (4,097) as bad (Table 2). There was a statistically significant difference in the percentage of (good) photographs that allowed the specimen identification when the camera trap was activated by the motion of an Andean bear in front of it and those (bad) photographs that did not (Mann-Whitney, P = 0.0001). Of all the videos acquired (264), 53.03 % (140) were rated as good and 46.97 % (124) as bad (Table 2). There was a statistically significant difference in the percentage of (good) videos that allowed the specimen identification when the camera trap was activated by the motion of an Andean bear in front of it and those (bad) videos that did not (Mann-Whitney, P = 0.0001). The combination of identification features that most frequently appeared in good photographs was facial marking-estimated body size-sex (in 51.98 % of these photographs); estimated body size (without other accompanying characteristics) was the identification feature that most frequently appeared in bad photographs (in 79.13 % of these photographs). Facial marking-pectoral marking-estimated body size-sex was the features combination that most frequently appeared in good videos (in 69.29 % of these); pectoral marking-sex was the combination appearing most frequently in bad videos (in 49.19 % of these).
Comparison of the results obtained with the two visual formats (photograph and video) showed a statistically significant difference between the percentage of good photographs and good videos acquired every time the camera trap was activated by the motion of an Andean bear in front of it (Mann-Whitney, P = 0.0001), as well as between the percentage of bad photographs and bad videos (Mann-Whitney, P = 0.0001).
Discussion
Results from this study showed (Table 2) that the visual file format chosen for recording affects the ability to identify specimens of Andean bear in camera trap work. The video format provided a better capacity to identify specimens, with a higher percentage (49.62 %) of successful recording events that resulted in the correct identification of the specimen, compared to that obtained from cameras set in photograph mode (26.77 %; Table 2). These results are in agreement with those reported by Zug (2009) (25 % successful recording events, out of 28) in Ecuador using photographs, but differ from those obtained by Ríos-Uzeda et al. (2007) (42.86 % successful recording events, out of 7) in Bolivia with the same visual file format. This difference may be due to the small number of recording events acquired by Ríos-Uzeda et al. (2007) , which prevented recognizing a clear pattern in the effectiveness of photographs for identifying Andean bears.
In our study, the higher percentage of video recordings that allowed the correct identification of Andean bear specimens (53.03 % good videos), compared to that obtained with photographs (5.25 % good photos; Table 2), is supported by the statistically significant difference (Mann-Whitney, P = 0.0001) found between the percentage of photographs and videos that allowed the correct identification of the specimen (good visual files) every time a camera was activated by the motion of an Andean bear in front of it. These results are in agreement with those reported by Higashide et al. (2013) for the Asian black bear (Ursus thibetanus) in Japan, where the percentage of videos that allowed the specimen identification was up to 70.00 % in sampling stations including single-camera traps.
Facial and pectoral markings are considered as the key distinctive features of Andean bear specimens (Roth 1964; Peyton 1999). In our study, the combinations of identifying features most frequently observed in photographs (facial marking-estimated body size-sex) and videos (facial marking-pectoral marking-estimated body size-sex) that successfully allowed the specimen identification (good visual files), compared with those observed in photographs (estimated body size without other accompanying characteristics) and videos (pectoral spot-sex) that did not (bad visual files), showed the greater capacity of the video format to clearly record the body markings of Andean bears. Studies on this species using photograph camera traps recommend using sampling stations supplied with 2 to 3 camera traps to acquire images of the specimens from different viewpoints in which body markings can be more clearly observed (Ríos-Uzeda et al. 2007; Zug 2009). However, in our study on Andean bear, as well as in that by Higashide et al. (2013) on Asian black bear, single video camera traps were used per season, combined with the use of attractant baits placed in front of the camera at a height that prompted the specimen to raise on its hind legs to reach the bait. The use of this sampling station configuration made it possible observing the features necessary to correctly identify Andean bear specimens using a single camera trap set on video format, obtaining a greater number of successful recording events (49.62 %; Table 2) compared to those obtained with two- (Ríos-Uzeda et al. 2007; 42.86 %) or three- (Zug 2009; 25.00 %) camera sampling stations using photograph format.
Observing the Andean bear’s body markings can be made difficult by illumination conditions under the forest canopy, which cause reflections that make observation of the markings shape and color difficult. The video recording of the specimens allows this negative effect to be ameliorated, as variations of light reflection on the markings can be tracked as the video progresses and the specimen changes its position in front of the camera. In most of the recording events acquired in this study the specimens recorded sought the bait by standing on their hind legs to reach it and approached the camera trap to inspect it closely. This allowed observing the specimens in different positions and distances from the camera, providing sufficient information on the presence, shape and color of facial and pectoral markings and of other key features such as the specimen’s body size and sex, as well as of additional reproductive characteristics such as the presence of turgid mammary glands or cubs accompanying postpartum females. However, some specimens do not show any body markings and cannot, therefore, be identified (D. Rodríguez com. Pers.), regardless of the visual file format used. This study presents the first formal report of this condition in Andean bears.
In the course of this study two bears bearing particular physical conditions related to restricted mobility of their hind limbs (limping and rigidity of one of the limbs) were video recorded. This would allow the identification of these particular specimens, and could not have been possible with the use of photographs.
The use of camera traps has proved to be very useful in the study of Andean bear populations (Ríos-Uzeda et al. 2007; Zug 2009). However, the proper application of population models depends in part on having an adequate individualization of the specimens recorded. This fact high-lights the importance of having a good knowledge of the relative efficiency of the different data formats that can be acquired with camera traps (photographs or videos) for identifying specimens, as a key aspect for the inclusion of these records in population studies (Foster and Harmsen 2012). The use of camera traps using video format is recommended.

