











 nueva página del texto (beta)
nueva página del texto (beta) Español (pdf)
Español (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El Tapir centroamericano (Tapirus bairdii; Perissodactyla: Tapiridae), también conocido como tixl (en idioma Q'eqchi'), es el mamífero terrestre de mayor talla de Guatemala y de la región Mesoamericana. Su distribución regional comprende desde sureste de México hasta el este de Ecuador y Colombia (Matola et al. 1997; Castellanos et al. 2008; Naranjo 2009; Schank et al. 2015). Se le puede encontrar desde el nivel del mar hasta los 3620 m de altura (Naranjo y Vaughan 2000), habitando en diversos ambientes tropicales y subtropicales, entre ellos bosques tropicales, manglares, bosques mesófilos, encinares y páramos (March 1994; Naranjo 2001).
A nivel internacional el tapir se encuentra en la lista roja de especies amenazadas de acuerdo a la Unión Internacional para la Conservación de la Naturaleza (IUCN) debido a la reducción de sus poblaciones (Castellanos et al. 2008). A nivel regional y local las principales amenazas son la pérdida y degradación del hábitat, y en menor medida la cacería (Matola et al. 1997; Castellanos et al. 2008). Debido a los requerimientos específicos de calidad hábitat, la presencia del tapir constituye un indicador del buen estado de conservación de las áreas donde aún está presente (Zapata y Dyer 2003; Botello et al. 2014).
Las primeras descripciones de la distribución de la especie para el país corresponden a la segunda mitad del siglo XIX, y abarcan la costa atlántica y pacífica del país, incluyendo las faldas de la cadena volcánica de la Sierra Madre, en el descenso del altiplano hacia la planicie costera del Pacífico, con elevaciones de 300 a 1,400 m (Alston 1918; Ximenez 1967; www.insivumeh.gob.gt). El conocimiento sobre la distribución del tapir se enriquece a mediados del siglo XX, cuando se reportó que el área de distribución también comprendía los departamentos de Petén, Quiché y Alta Verapaz (Saunders et al. 1950; Ibarra 1959). A finales de la década de 1990, Matola et al. (1997) reportaron que el tapir estaba presente solamente en cuatro departamentos, siendo éstos Quiché, Alta Verapaz, Izabal y Petén, evidenciando de esta manera su extinción en la región sur del país. A inicios del presente siglo se iniciaron los estudios específicos sobre el tapir en el país (al igual que otros estudios de fauna), por medio de los cuales fue posible verificar (a través de huellas, excretas, fototrampeo y material de colecciones científicas) que su distribución actual está restringida a los departamentos de Alta Verapaz, Baja Verapaz, Quiché, El Progreso, Izabal y Petén (Kawanishi 1995); Baur 1999; Bestelmeyer y Alonso 2000; Novack 2003; García y Radachowsky 2004; Phillips y Dudik, 2008; Morales et al. 2005; Ruíz y García 2005; Ruíz y Rodríguez 2006; García 2006; Moreira et al. 2007; García et al. 2008a, 2008b, 2009, 2010a, 2010b; Gómez 2011).
Siendo las principales amenazas para el tapir la cacería y la pérdida de hábitat, se puede deducir que la extinción local de la especie está asociada con el cambio del uso del suelo y la consecuente fragmentación del hábitat (Castellanos et al. 2008). En este escenario, los remanentes de hábitat y el patrón espacial de los mismos, son elementos de relevancia para la conservación de la especie en el país. En el presente estudio se busca desarrollar una clasificación del hábitat del tapir en el país que pueda ser aplicada en la planificación y ordenamiento del territorio. La clasificación propuesta se basa en tres aspectos: presencia de la especie en la última década (2005-2015), la importancia de cada remanente boscoso para la conectividad del sistema y la extensión bajo mecanismos de protección formal dentro de cada remanente boscoso.
Materiales y métodos
Área de estudio. El área de estudio comprendió toda la superficie del territorio guatemalteco, entre las latitudes 13.7° y 17.8° N y longitudes -88.2° y -92.2° O, abarcando una extensión continental de 108,889 km2. Se recopilaron registros de presencia de tapir a través de visitas a 26 de las 34 áreas protegidas ubicadas dentro del ámbito de distribución reportado por Matola et al. (1997), durante las cuales se recorrieron los sitios sugeridos previamente por los guías, pobladores, registros históricos y conversaciones con técnicos, guarda recursos y administradores de las áreas protegidas. Durante las visitas de campo se realizaron caminatas para buscar y referenciar los rastros que evidenciaban la presencia de la especie (huellas, excretas, comederos, echaderos y caminos). La fase de campo fue realizada entre los años 2007 y 2009 en áreas del Sistema Guatemalteco de Áreas Protegidas (SIGAP). Estos datos fueron complementados con registros de presencia de tapir en el país, correspondientes a registros actuales e históricos de huellas, excretas, comederos, echaderos y fototrampeo, generados a partir de investigaciones científicas (Ruíz y García 2005; Ruiz y Rodríguez 2006; García et al. 2008b; 2009), bases de datos de los programas de monitoreo biológico de instituciones locales (Fundación Mario Dary -FUNDARY- datos del 2005-2007; Centro de Estudios Conservacionistas de la Universidad de San Carlos de Guatemala (CECON) datos del 2014-2015; Fundación Defensores de la Naturaleza (FDN) datos del 2014) y revisión bibliográfica. Todos estos datos fueron compilados en una base de datos digital.
Modelado de la distribución potencial. El modelado de distribución potencial de tapir se realizó utilizando el algoritmo Maxent Ver. 3.3.3 k (Phillips y Dudik, 2008). Para ello se utilizaron 10 variables climáticas (BIO1, BIO4, BIO5, BIO6, BIO12, BIO15, BIO16, BIO17, BIO18 y BIO19) y de la elevación (ALT), en formato raster con resolución de 1 km2, provenientes de la base de datos versión 1.4 de WorldClim (Hijmans et al. 2005), siguiendo a Schank et al. (2015).
Dado que se cuenta únicamente con tres registros para la vertiente del Pacífico, donde de acuerdo con Alston (1918) la especie aún era abundante a principios del siglo pasado, se asume que existe un sesgo en los registros de presencia actual con respecto a su distribución potencial, ya que no se cuenta con registros de localidades que fueron adecuadas para la especie pero que en la actualidad han sido deforestadas (GIMBOT, 2014). Debido a lo anterior, con el fin de contar con una distribución más uniforme de los registros de presencia para reducir este sesgo y el efecto de un mayor muestreo adentro de áreas protegidas, utilizando ArcMap ver. 10 (ESRI, Redlands, CA, EE. UU.) se generó una cuadrícula de 10 x 10 km2 que se sobrepuso a la capa de registros de presencia existentes y se procedió a seleccionar únicamente un registro para cada celda, identificando de manera visual el registro (de la base de datos compilada) ubicado más cerca del centro de la celda. Por último se seleccionaron las opciones de curvas de respuesta, Jacknife y los ajustes preseleccionados por Maxent (default settings).
Análisis morfológico de los patrones espaciales. El mapa de distribución potencial generado con Maxent se reclasificó en ArcMap 10 a partir el umbral de corte de 0.3 que se estableció con base en las categorías utilizadas por Mendoza et al. (2013) y Schank et al. (2015) (incluyendo 3-5 low suitability, 5 - 7 very suitable y > 7 high suitability). El mapa binario resultante fue transformado a formato vectorial para realizar una extracción del Mapa de Cobertura Forestal por Tipo y Subtipo de Bosques en Guatemala (GIMBOT, 2014), y así obtener un mapa de los remanentes de hábitat para el tapir para el año 2012 con una resolución de 5 x 5 m. El resultado fue transformado de nuevo a un mapa binario formato raster con una resolución de 250 x 250 m.
Con el fin de determinar el patrón morfológico espacial del hábitat potencial remanente, el mapa binario de presencia-ausencia correspondiente a las zonas con mayor probabilidad de distribución de la especie se procesó con la herramienta Morphological Spatial Pattern Analysis (MSPA) del Programa GUIDOS ver. 1.3, la cual utiliza una secuencia de operadores matemáticos para clasificar los objetos de una imagen en clases de acuerdo a su geometría, arreglo espacial y conectividad estructural (Vogt et al. 2007; Soille y Vogt 2009). El resultado de este procesamiento proporcionó al mapa del hábitat potencial valores correspondientes a las siete clases del MSPA: área de interior (core area), relicto (islet), perforación (perforation), borde (edge), puente (bridge), lazo (loop) y rama (branch) (Vogt et al. 2007; Correa y Mendoza 2013). A partir de este mapa se generó un mapa con los principales remanentes de hábitat del tapir de acuerdo a su patrón morfológico espacial, incluyendo solamente las áreas interiores, sus bordes y las perforaciones.
Conectividad de los remanentes de hábitat. La capa de remanentes de hábitat resultante se transformó en formato vectorial para ser procesada con el programa CONEFOR Sensinode ver. 2.5.8 (Pascual-Hortal y Saura 2006; Saura y Pascual-Hortal 2007). A través de este programa se evaluó la conectividad funcional potencial para el tapir, estimando el diferencial del Índice Integral de Conectividad (dIIC) a cada uno de los remanentes de hábitat, el cual se calcula a partir de su conectividad interna, su contribución con el flujo y su contribución como conector (stepping stones). La distancia umbral de dispersión utilizada entre dos remanentes de hábitat para el tapir fue de 1,500m, con base a los datos de Medici (2010) para el tapir de tierras bajas Tapirus terrestris en Brasil. Este procedimiento permitió identificar los remanentes de hábitat de mayor importancia de acuerdo a su aporte a la conectividad de todo el sistema.
Clasificación de principales remanentes de hábitat potencial. Con base en la información generada a partir del modelado de distribución potencial, el análisis morfológico de los patrones espaciales, el Índice Integral de Conectividad (dIIC) y el mapa de distribución del tapir a nivel regional (Schank et al. 2015), se desarrolló una propuesta de clasificación para los principales remanentes de hábitat potencial del tapir en el país. Tomando estos tres aspectos como base, se generó un cuadro de ponderación para los remanentes de hábitat potencial (modificado de García et al. 2008b, 2009). Se evaluaron los 25 remanentes con la mayor extensión con el fin de limitar el análisis a un número determinado. Los remanentes con afinidad espacial y administrativa, pertenecientes a una misma área protegida o región específica fueron reclasificados en complejos.
En la clasificación desarrollada se proponen cinco categorías (A1, A2, A3, B1 y B2) para la clasificación del hábitat potencial del tapir, las cuales representan la relevancia de los principales remanentes para la conservación de la especie en el país a partir de la presencia actual de la especie, el Índice Integral de Conectividad y la presencia de mecanismos formales de protección de la cobertura forestal y la diversidad biológica (áreas protegidas). Los remanentes tipo A corresponden a localidades donde la especie está presente en la actualidad y las B donde ya no está presente o no se tienen reportes recientes. Los remanentes A1 corresponden a zonas de gran extensión territorial, donde la conectividad es alta y existen mecanismos formales de protección (áreas protegidas oficialmente declaradas). Los remanentes tipo A2 son aquellos que albergan poblaciones de tapires pero que su conectividad es media. En los remanentes A3, la especie está presente y poseen una baja conectividad. En los remanentes tipo B1 la conectividad es baja y existen mecanismos formales de protección y los remanentes B2 se diferencian de los primeros por no presentar mecanismos formales de protección.
Resultados
Modelado de la distribución potencial. La base de datos de registros de presencia de tapir que se generó contiene un total de 206 registros con referencia espacial, de los cuales 148 corresponden a la verificación en campo durante el período 2008-2009, en 15 áreas protegidas del país (Biotopo Protegido Naachtún-Dos Lagunas, Biotopo Protegido San Miguel La Palotada-El Zotz, Concesión Forestal Carmelita (ZUM), Concesión Forestal Uaxactún (ZUM), Parque Nacional Laguna del Tigre, Parque Nacional Laguna Lachuá, Parque Nacional Mirador-Río Azul, Parque Nacional Sierra del Lacandón, Parque Nacional Tikal, Parque Nacional Yaxhá-Nakum-Naranjo, Refugio de Vida Silvestre Machaquilá, Reserva Biológica San Román, Reserva Comunitaria Indígena Bioitzá (ZUM), Reserva de Biosfera Montañas Mayas Chiquibul y Reserva de Biosfera Sierra de las Minas), 15 corresponden al programa de monitoreo biológico desarrollado por instituciones locales, 28 a comunicaciones personales y 15 a registros bibliográficos. Para el modelo se utilizaron 62 registros de presencia seleccionados con base a la metodología para obtener una distribución uniforme de los registros. El modelo de distribución potencial dio como resultado un área bajo la curva (AUC) de 0.815. Las variables con los mayores porcentajes de contribución son BIO17 = Precipitación del trimestre más seco (38.8 %), BIO4 = Temperatura estacional (desviación estándar * 100) (27.9 %) y BIO15 = Coeficiente de variación de las precipitaciones estacionales (8.4 %).
Los valores de Maxent se presentan en un rango que va de 0 a 1, donde los valores más cercanos a 1 indican mayor probabilidad de ocurrencia del tapir, debido a una mejor condición del hábitat. De acuerdo a lo anterior en el modelo se observa que la mayor probabilidad de presencia de la especie está focalizada en la región nororiente del país (Petén, Izabal, Alta Verapaz, Baja Verapaz y una región de Quiché), siendo aún mayor en el noreste y oeste de este bloque. Además, sobresalen pequeñas zonas con alta probabilidad de distribución ubicadas a lo largo de la cadena volcánica, al sur del territorio nacional (Figura 1). El resto del territorio presenta una baja probabilidad de distribución.

Figura 1 Modelo de distribución potencial de Tapirus bairdii en Guatemala generado con Maxent. Las áreas de color rojo muestran mayor probabilidad de ocurrencia de la especie y las áreas en color azul muestran la menor probabilidad de ocurrencia.
Análisis morfológico de los patrones espaciales. El mapa binario que se generó indica que la extensión total de hábitat potencial para el tapir abarca alrededor de 58,456.87 km2, es decir el 53.68 % del territorio nacional, sin embargo, al eliminar las áreas donde no existe cobertura forestal, la extensión del hábitat potencial se reduce a 26,095.82 km2, equivalente al 44.54 % de su extensión potencial (GIMBOT, 2014).
La clasificación morfológica de los patrones espaciales permitió conocer mejor la dinámica de los fragmentos de hábitat. En la Tabla 1 y Figura 2 se presentan los resultados de las siete clasificaciones de la cobertura, correspondientes al mapa binario de presencia-ausencia.
Tabla 1 Clasificaciones del análisis morfológico de los patrones espaciales del hábitat potencial de Tapirus bairdii en Guatemala.
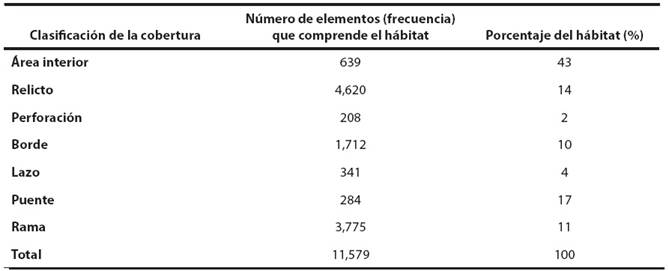
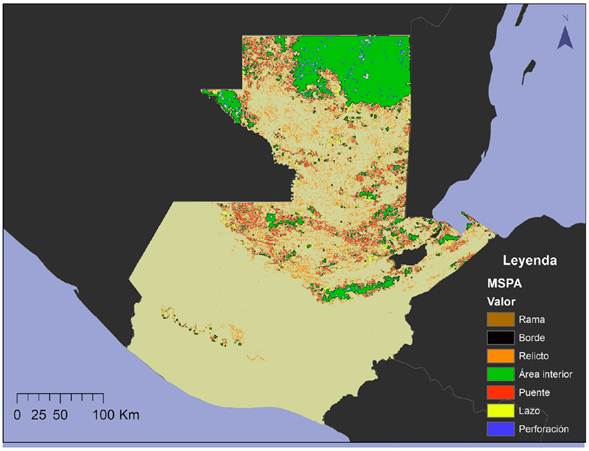
Figura 2 Mapa de las siete clasificaciones del análisis morfológico de los patrones espaciales del hábitat potencial de Tapirus bairdii en Guatemala.
La herramienta MSPA estimó que el mayor porcentaje del hábitat del tapir al 2012 corresponde a área interior (43 %), seguido de los elementos conectores, que incluye las clasificaciones lazo, puente y rama, que en su conjunto representan el 32 %. Sin embargo, es importante resaltar que la mayoría de los elementos fueron clasificados como relictos (remanentes de bosque aislados) y borde (4,620 y 1,712 respectivamente) a pesar de no presentar porcentajes elevados (14 % y 10 % respectivamente).
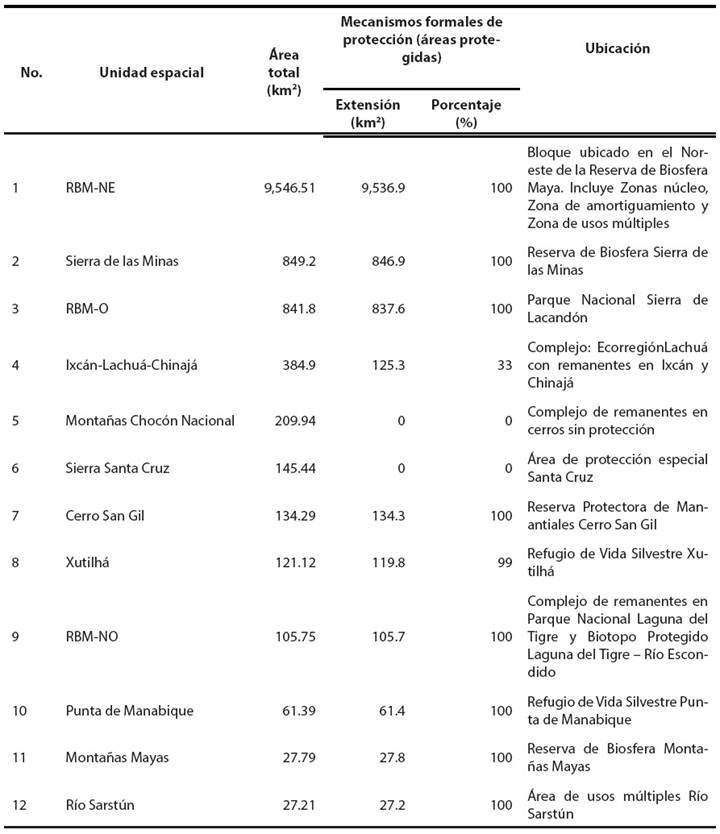
Se seleccionaron los 25 remanentes de mayor superficie que están ubicados en el ámbito de distribución de la especie, los cuales fueron organizados en 12 unidades espaciales (remanentes o complejos de remanentes) que se describen en la Tabla 2.
Tabla 2 Descripción de 12 remanentes boscosos de mayor tamaño dentro del área de distribución de Tapirus bairdii en Guatemala.
Conectividad de los remanentes de hábitat potencial. El resultado del análisis de conectividad muestra que el remanente con el mayor dIIC es el Noreste de la Reserva de Biosfera Maya -RBM-NE- (33.505662). El resto de remanentes presentan una baja conectividad, a excepción de los remanentes Sierra de las Minas y el Oeste de la Reserva de Biosfera Maya -RBM-O- que son de mayor tamaño (0.51311405 y 0.38876015 respectivamente). El resto de remanentes presenta un Índice Integral de Conectividad menor a 0.1 (Tabla 3 y Figura 3).
Tabla 3 Índice Integral de Conectividad de los 12 remanentes boscosos de mayor tamaño dentro del área de distribución del Tapirus bairdii en Guatemala.

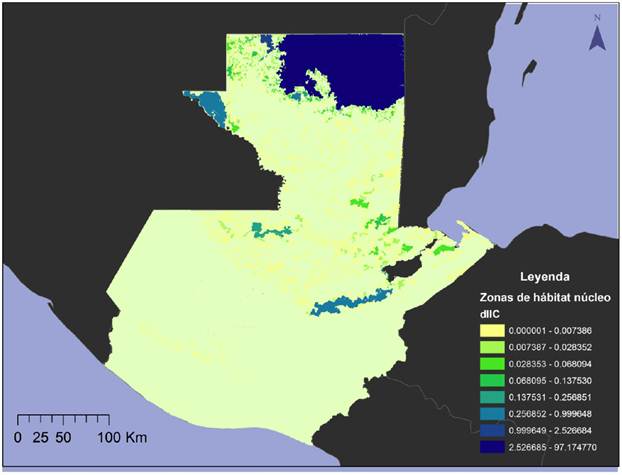
Figura 3 Mapa del Índice Integral de Conectividad para 12 remanentes boscosos de mayor tamaño dentro del área de distribución de Tapirus bairdii en Guatemala. En color amarillo claro se encuentran los remanentes con menor dIIC y en color azul los remanentes con mayor dIIC.
Clasificación de principales remanentes de hábitat potencial. La clasificación de las 12 unidades espaciales de hábitat potencial para el tapir en Guatemala se presenta en la Tabla 4 y Figura 4. Los remanentes de mayor importancia son el noreste de la RBM (incluyendo zonas núcleo, de usos múltiples y de amortiguamiento) y los remanentes correspondientes a la Reserva de Biosfera Sierra de las Minas y Parque Nacional Sierra del Lacandón.
Tabla 4 Clasificación de 12 remanentes boscosos de mayor tamaño en el hábitat potencial de Tapirus bairdii en Guatemala.


Discusión
Modelado de la distribución potencial. El resultado del modelo de distribución potencial del tapir en Guatemala es semejante al obtenido por Schank et al. 2015. En ambos casos se evidencia alta probabilidad de presencia de la especie en la sección noreste de la RBM, la cual forma parte del bloque transfronterizo de la Selva Maya (Reserva de la Biosfera Calakmul en México, RBM en Guatemala y Río Bravo en Belice), y la sección oeste donde se ubica el Parque Nacional Sierra del Lacandón (colindante con la Reserva de la Biosfera Montes Azules, México). De acuerdo con Naranjo (2009) estas zonas de México son de gran importancia ya que albergan alrededor del 50 % de la población de tapir en ese país.
Esta región de alta probabilidad de distribución incluye en su extremo inferior al Parque Nacional Laguna Lachuá, Reserva de Biosfera Sierra de las Minas y áreas protegidas de Izabal, donde su presencia ha sido reportada durante la última década. Al igual que Schank et al. 2015, el presente modelo sobreestima (aunque en menor escala) la presencia de tapir en la región central y sur del país (específicamente en la cadena volcánica), donde existe alta fragmentación y cambio de uso del suelo debido a las actividades agropecuarias (p.e. algodón, caña de azúcar, ganadería extensiva) que han contribuido a su extinción local, sin embargo, es un área de distribución histórica (Alston 1918; Saunders et al. 1950).
Es importante resaltar que las zonas de alta probabilidad de distribución de la especie en el modelo coinciden en un alto porcentaje con áreas protegidas. De acuerdo al análisis del modelo, las áreas protegidas constituyen los últimos refugios donde se alberguen poblaciones de tapir en el país, aspecto que evidencia la importancia que tienen estos espacios en la supervivencia de esta especie y de las especies que están asociadas a su hábitat.
El alto porcentaje de contribución de la variable BIO17 (precipitación del trimestre más seco) puede sugerir, de manera similar a los resultados de Carrillo-Reyna et al. (2015) para la península de Yucatán, que los tapires no se distribuyen en las zonas más calientes con escasa precipitación. En este sentido la disponibilidad de agua puede ser un factor limitante de la distribución de la especie, especialmente en ambientes con poca disponibilidad de agua superficial como ocurre en la región RBM-NO, al igual que áreas aledañas de México en el estado de Campeche (Pérez-Córtez et al. 2012; Carrillo-Reyna et al. 2015).
Análisis morfológico del patrón espacial. Las zonas interiores constituyen el mayor porcentaje de acuerdo al análisis MSPA, situación que se evidencia principalmente en el gran bloque de la RBM-NE, donde la cobertura forestal es mayor y la fragmentación es baja. La pérdida de hábitat y la fragmentación son las principales amenazas para la especie en la región (Castellanos et al. 2008). Las poblaciones están quedando cada vez más aisladas, lo que las hace más vulnerables ante la cacería y las enfermedades, reduciendo su éxito reproductivo y la supervivencia de los juveniles, entre otros aspectos (Deyoung y Honeycutt 2005). En Brasil, Gonçalves (2007) evaluó el efecto de la fragmentación en cinco especies de mamíferos y encontró que la fragmentación las afecta en diferente escala de acuerdo con el tamaño corporal. Sin lugar a duda el patrón morfológico de la cobertura forestal tiene un efecto directo en la supervivencia de las especies (Akçakaya y Brook 2009), por lo que su análisis aporta información esencial para el manejo del hábitat del tapir.
Las categorías de relicto y rama fueron las clasificaciones más numerosas (4,620 y 3,775 respectivamente) pero no las más representativas (14 % y 11 % respectivamente) obtenidas a partir del Análisis Morfológico de los Patrones Espaciales. Estos dos elementos evidencian el proceso de fragmentación. En el caso de relicto los resultados muestran que existe una mayor área con pequeños fragmentos de hábitat, que son producto de la pérdida paulatina del hábitat; a pesar de constituir zonas potenciales de alimentación es importante recuperar la conexión de éstas hacia áreas interiores, puesto que por sí mismas no pueden albergar poblaciones viables (Correa y Mendoza 2013). Lo anterior se ve reflejado en la zona sur de la RBM, las Verapaces e Izabal, donde la fragmentación del hábitat se ha incrementado durante los últimos años reduciendo la cobertura forestal dentro de las áreas protegidas existentes y aislándolas aún más (GIMBOT 2014).
De acuerdo con Soille y Vogt (2009) las áreas categorizadas como puente contribuyen a conectar físicamente los fragmentos de hábitat, favoreciendo la conectividad entre áreas. Sin embargo, es importante realizar un análisis más detallado, ya que de acuerdo con Correa y Mendoza (2013) estos elementos pueden ser clave para la conectividad entre áreas pero también pueden ser el resultado de la reducción de los fragmentos de hábitat por intervención antrópica.
Conectividad de los remanentes de hábitat potencial. El resultado del análisis de conectividad concuerda con los obtenidos por García et al. (2011), quienes reportan que el dIIC más alto corresponde a la Selva Maya. Además de la integridad ecológica de la RBM, su colindancia con otras zonas boscosas de México y Belice (Mendoza et al. 2013; Carrillo-Reyna et al. 2015; Schank et al. 2015) hace de ésta la zona más importante por la conectividad que presenta tanto a nivel interno como externo contribuyendo de esta forma a mantener el flujo de las poblaciones de fauna silvestre como las de tapir.
Los remanentes boscosos que le siguen en orden de importancia de acuerdo al dIIC son el oeste de la RBM, representado por Sierra del Lacandón, los bloques de Sierra de las Minas y la Ecoregión Lachuá. Estas áreas presentan los últimos remanentes de mayor tamaño donde a pesar de no tener conexión con otras áreas, a excepción de Lacandón que si presenta conexión externa con la Reserva de la Biosfera Montes Azules, la conectividad interna aún es relativamente alta.
Al igual que lo reportado por García et al. (2011), el resto de áreas ubicadas en Izabal y la región sur de Petén presentaron un dIIC bajo, evidenciando la baja conectividad con otras áreas circundantes, así como la baja conectividad que existe a nivel interno, resultado de la fragmentación de estos bloques. No debe descartarse que estas zonas puedan ser utilizadas como remanentes de paso (stepping stones) y como núcleos de restauración.
Muench (2006) llevó a cabo una caracterización del estado de la conectividad en tres zonas de la selva Lacandona en México e identificó 40 corredores, clasificados en seis categorías y localizados sobre seis unidades del paisaje. Los resultados de la conectividad de los remanentes boscosos en conjunto con la información de los patrones morfológicos obtenidos en este estudio pueden ayudar a identificar a mayor detalle las áreas donde la conectividad externa puede ser mejorada a través de diferentes mecanismos de manejo como el establecimiento de programas de restauración ecológica.
Clasificación de principales remanentes de hábitat potencial. El remanente que corresponde al noreste de la RBM ya ha sido identificado como el área de mayor importancia para la conservación del tapir (Matola et al. 1997; García et al. 2009; 2010b; Schank et al. 2015). Naranjo (2009) identificó como una zona de importancia global para las poblaciones de tapir al remanente de Calakmul, que colinda con este bloque de la RBM, por contar con la mayor población de tapir en vida silvestre a nivel global.
Con respecto a los remanentes tipo A2, Sierra de las Minas es un área con baja conectividad hacia otros remanentes de hábitat pero con alta conectividad interna debido a su extensión, mientras que Sierra del Lacandón presenta alta conectividad tanto interna por su extensión como hacia otros remanentes, ya que colinda con la Reserva de la Biosfera Montes Azules en la Selva Lacandona (Muench 2006; Naranjo 2009).
De acuerdo a los resultados obtenidos en la clasificación de los principales remanentes de hábitat potencial y a los análisis realizados, se puede evidenciar una disminución y deterioro del hábitat del tapir en el país en los últimos cinco años, al compararlos con evaluaciones previas realizadas para el Sistema Guatemalteco de Áreas Protegidas (SIGAP) por García et al. (2008b, 2009, 2010b). Algunos de los fragmentos que fueron considerados previamente han desaparecido o se ha reducido el área boscosa a fragmentos menores a 1 km2, como es el caso del remanente correspondiente a la Reserva Biológica San Román, en la cual García et al. (2009) obtuvieron en el año 2008 un registro de presencia de la especie, y para el 2012 la cobertura boscosa casi en su totalidad había sido reemplazada por monocultivo de palma africana (GIMBOT 2014). Incluso en la RBM-NE, el remanente de mayor extensión en el país, presenta una reducción de la cobertura y modificación del patrón morfológico espacial. Lo anterior, permite deducir que el estado de conservación de las poblaciones silvestres de la especie en el país es vulnerable, ya que existe una tendencia continua a la pérdida y degradación de hábitat con la consecuente reducción o extinción de poblaciones locales de tapires. Se hace evidente la necesidad del desarrollo de herramientas de gestión del territorio e implementación de acciones que reviertan esta tendencia así como el fortalecimiento del SIGAP.

