











 nueva página del texto (beta)
nueva página del texto (beta) Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroduction
The Maya Mountains are a heavily forested mountain range in Belize and Guatemala, and form the southernmost part of La Selva Maya, the largest tropical forest block in the Americas after the Amazon (Radachowsky et al. 2012; Briggs et al. 2013). The diverse geology accounts for a range of different soil types and a dramatic topography, which result in a multitude of microhabitats. These characteristics, combined with tropical rainfall regimes, yield some of the highest levels of biodiversity in the region (Brewer and Webb 2002; Dourson 2012). The Maya Mountains are also part of the Mesoamerican Biological Corridor, a regional initiative to conserve connectivity among forests in Central America (Herrera 2003). On a national level, the Maya Mountains cover 22.2 % of Belize's land mass, forming the largest contiguous forest in the country, and are regarded as a Key Biodiversity Area (Meerman 2007). They are estimated to provide up to 1 billion USD worth of ecosystem services to the people of Belize (Hammond et al. 2011), and constitute one of biggest resources in the country for local livelihoods, forestry and eco/agritourism (Briggs et al. 2013). Despite the fact that most of the Belizean side of the mountain range consists of protected areas of various legal categories (Figure 1), increasing industrial and small scale clear-cutting have severed the connections between the Maya Mountains and the forests to the north in both Guatemala and Belize. Consequently, the Maya Mountains are becoming increasingly isolated (Briggs et al. 2013).
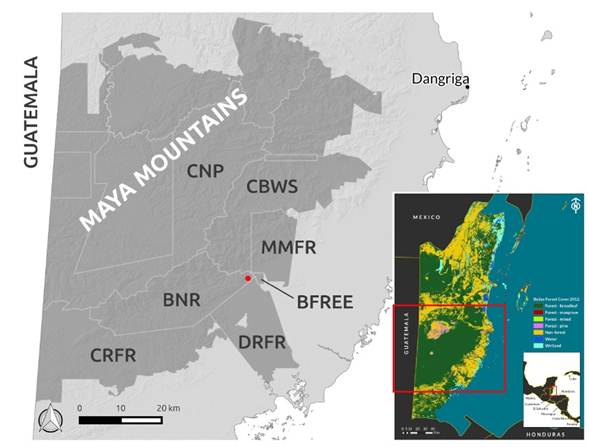
Figure 1 Location of Belize and the Maya Mountains. The protected areas of the Maya Mountains (as delineated by Briggs et al. 2013) are shaded grey and the trapping area is indicated by a red dot. Protected areas that are directly relevant to the study area are annotated. BFREE: Belize Foundation for Research and Environmental Education; BNR: Bladen Nature Reserve; CNP: Chiquibul National Park; CBWS: Cockscomb Basin Wildlife Sanctuary; CRFR: Columbia River Forest Reserve; DRFR; Deep River Forest Reserve; MMFR: Maya Mountain Forest Reserve.
The white-lipped peccary Tayassu pecari (Link 1795) is often one of the first mammals to disappear with increasing forest fragmentation (Sowls 1997; Moreno and Meyer 2014). White-lipped peccaries are gregarious ungulates travelling in herds of 10 to 300 individuals (Sowls 1997; Altrichter et al. 2012), and although they are known to occur in mosaic landscapes, they are regarded as typical forest dwellers (Fragoso 1999; Reyna-Hurtado and Tanner 2005; Keuroghlian and Eaton 2008; Desbiez et al. 2009; Tobler et al. 2009; Reyna-Hurtado et al. 2009). Due to the large herd size, they need to move frequently among different feeding spots, and in the process cover large distances through the forest. Home range sizes throughout the species' range vary substantially, ranging from 20 to 200 km2 (Fragoso 2004; Keuroghlian et al. 2004; Reyna-Hurtado et al. 2009; de Almeida Jácomo et al. 2013). This variation reflects the environmental variability over the range of the species and illustrates its ability to cope with different conditions. Nevertheless, this flexibility seems to consist mainly of varying herd size (Fragoso 2004) or splitting up in sub-herds (e. g.Keuroghlian et al. 2004), rather than of changing habitat preferences. Thus, despite their ability to deal with different environmental conditions, white-lipped peccaries always depend on intact forest habitats of sufficient size and connectivity, rendering them highly susceptible to forest conversion and fragmentation. The Maya Mountains are assumed to offer continuous habitat and are expected to be home to several herds of white-lipped peccaries. However, no studies have been conducted on this species in Belize; their distribution, population size, herd dynamics and movement patterns are unknown for the country. The rugged topography of the Maya Mountains might render some parts of the forest unsuitable, and species-specific connectivity among suitable forest areas remains unstudied.
Here, we provide a preliminary report on data gathered from a herd of ca. 60 by tracking a single white-lipped peccary that was GPS-collared to investigate its home range and movement patterns in the Bladen Nature Reserve, southern Belize. We assume that the location of the GPS-collared individual adequately represents the area used by its herd at the same point in time, because in many gregarious species the tendency of individuals to always move with the herd is very strong. In fact, this tendency has been used for management purposes in conservation. For example, Taylor and Katahira (1988) used the 'Judas goat' approach to help eradicate feral goats (Capra hircus) on the Hawaiian Islands, and Campbell et al. (2004) and Cruz et al. (2009) used it for similar reasons on the Galápagos Islands. That is, goats fitted with a radio-collars (the Judas goats) were used to get information about the location of an entire herd of feral goats. All members of the discovered herds were killed, with the exception of the Judas goats, which would go on to find another herd, where the same scenario was repeated. A similarly strong herding tendency was described by Byers and Bekoff (1981) for collared peccaries (Pecari tajacu), and we can confirm such cohesiveness for white-lipped peccaries too based on camera trap footage from within our study area (M. Hofman, unpublished data) and evidence from several earlier studies on the species elsewhere (Keuroghlian et al. 2004; Reyna-Hurtado et al. 2009). For example, in neighbouring Calakmul, Mexico, collared animals were always seen with their respective herds for more than 700 observations (Reyna-Hurtado pers. comm.). Hence, the collaring of additional individuals from the same herd would not necessarily yield additional information on home range and movement patterns of that herd.
Methods
Study area. The Bladen Nature Reserve (BNR; 16.48, -88.88) is a strictly protected area (IUCN Cat. Ia) gazetted in 1990, covering about 40.5 km2 of the southern Maya Mountains, and centred around the watershed of the Bladen branch of the Monkey River. Since 2008, the reserve has been managed by a local conservation NGO, the Ya'axché Conservation Trust, under mandate of the Belize Forest Department. Vegetation is almost exclusively lowland and submontane broad-leaf wet forest with relatively open understorey and a canopy height of 20-30 m (Iremonger et al. 1995). Lowland areas are characterised by abundant Astrocaryum mexicanum and Attalea cohune palms, while tree diversity is dominated by the Fabaceae, Rubiaceae, Sapotaceae and Lauraceae families (Iremonger et al. 1995; Brewer and Webb 2002; Stott 2014). The protected area also contains a sliver of seasonally inundated lowland savannah in its far eastern tip, and a pocket of pine forest. Due to its inaccessible terrain with steep slopes and extremely narrow valleys, the area holds some of the most undisturbed forests of Belize and is very high in biodiversity (Brewer and Webb 2002; Dourson 2012; Dourson 2012), with ongoing species discoveries (Polhemus and Carrie, 2013; Thompson and Dourson 2013). Its elevation spans from 30 to 1050m, some of the highest areas in Belize. It is one of the wettest areas in Belize receiving an average annual rainfall between 2,500 and > 3,000 mm, with 85 % falling between mid-May and the end of November (Brewer and Webb 2002), and an annual average temperature of around 26 ºC. The area has a distinct dry season from March to mid-May. It is bordered by six other protected areas with a lower protection status, which are equally likely to be used by white-lipped peccaries, given the contiguous habitat type and expected ranging behaviour of the species (Figure 1, 3). The nearest human settlements are Medina Bank (~ 300 inhabitants, mostly Q'eqch'i Maya) and Trio Village (~ 3,000 inhabitants, mostly Q'eqch'i Maya and Mestizos from Guatemalan origin), and their agricultural areas border the Maya Mountain Forest Reserve and Belize Foundation for Research and Environmental Education. BNR is separated from a two-lane highway to the south-east by a five km wide stretch of savannah and lowland broadleaf forest, mostly within the Columbia and Deep River Forest Reserves.

Figure 2 Semi-variogram of all GPS fixes of a single white-lipped peccary in southern Belize (n = 69). Circles represent the estimated semi-variance for each time-lag, with 95 % confidence intervals estimated from the standard error of the mean semi-variance shaded light grey. The fitted OUF model (see text) is shown as a red solid line with its 95 % confidence intervals estimated from the standard error of the best fit shaded light red.

Figure 3 Spatial representation of the home range estimates for a single white-lipped peccary herd in southern Belize. a) 95 % Kernel Density Estimate home range (dark grey line), Core area (80 % KDE - black line) and 100 % MCP home range (orange line) over the relocations (red dots), rivers (blue) and roads (red lines). Black dots represent villages, the yellow star is the corral trap location. Forest cover is shaded green, grey shaded areas are non-forest (savannah, shrubs and agricultural lands), and grey diagonally line-filled areas with straight black boundaries are protected areas. b) Semi-variogram home range estimate (black ellipse) with 95 % confidence interval (grey ellipses) and utilisation density (blue shading).
White-lipped peccaries are occasionally spotted by Ya'axché field staff throughout the reserve, with estimated herd sizes between 60 and 100 individuals, but it has been unclear how many herds roam the area and whether or not they interact. Anecdotal reports from farmers and hunters suggest a declining population over the last 10 years in southern Belize. There are no reports about crop raiding by white-lipped peccaries in southern Belize and we assume that the animals in the research area have little or no contact with humans, with the exception of enforcement personnel and visiting researchers. Hunting pressure in eastern BNR is expected to be low due to long-term intensive enforcement activities in the area. In other parts of the reserve and in the surrounding areas, hunting pressure is expected to be higher (M. Hofman, pers. obs.).
Trapping. White-lipped peccaries have been trapped either by tracking or ambushing free-range animals (Fragoso 1998; Fuller et al. 2002; Reyna-Hurtado et al. 2009), or by using baited box and/or corral traps (Sowls 1997; Carrillo et al. 2002; Keuroghlian et al. 2004; de Almeida Jácomo et al. 2013; C. Richard-Hansen, pers. comm.). We used one box trap (1 x 1 x 2m) and one corral trap (Ø ca. 6 m, 1.5 m high, with handling compartment), spaced about 2.5 km apart, each in the proximity of a creek showing signs of peccary activity. The traps were deployed in July and August 2014, during the peak rainy season in southern Belize. Camera traps had indicated previous occasional presence at the corral trap site during two previous years. The traps were baited with various fruits; as well as cassava (Manihot esculenta) and cocoyam (Xanthosoma spp.) roots. Because only one collar was available, one adult female of appropriate size (estimated > 25 kg) was selected and isolated from the captured individuals, using recommended pig handling tools, including a confined restraining compartment, board and snare (Defra 2003; Grandin 2013). With the aim of minimizing the duration of the process, we did not use sedation. All animal trapping and handling followed the guidelines of the American Society of Mammalogists (Sikes and Gannon 2011), and was permitted by the Belize Forest Department [Permit no. CD/60/3/14(37)]. The individual was fitted with a Telonics TGW-4570-3 GEN4 GPS/Iridium collar (Telonics, Mesa AZ, USA). The collar was programmed to start a fix attempt every three hours, theoretically yielding eight locations per day. If no GPS fix was obtained after 180 seconds, the fix attempt was aborted and classified as unsuccessful. Data were scheduled to transfer to the satellite every two days. Data remained on the sending list for 10 days, then would remain stored on-board but would not be sent.
Home range size and movements. We used the semi-variance approach developed by Fleming et al. (2014) to detect whether the herd displayed home ranging behaviour, and to estimate the home range size. The approach makes use of the auto-correlated nature of tracking data to investigate processes that occur over different time scales (e. g. foraging vs. migration or dispersal). The semi-variogram measures the distance between two relocations as a function of the time lag between them, and calculates the variability of distances among all relocation pairs with the same time lag. When an animal displays home ranging behaviour, increasing the time lag between relocations leads to an asymptote in distance covered, because animals would not usually travel further than their home range diameter, even when given more time. Hence, the approach can be used both to detect home range behaviour and to estimate home range size. Since movement distances are averaged within time lags, the approach is robust in the face of irregular and sparse relocation data, i.e. varying fix intervals and long data gaps (Fleming et al. 2014).
We used the ctmm package (Fleming and Calabrese 2015) for the R statistical environment (R Development Core Team 2010) for all semi-variance calculations. Following Fleming et al. (2014), we calculated the empirical variogram, and selected the best fitting Semi-Variance Function (SVF) from a set comprised of i) a null-model (Brownian motion, i. e. random, undirected movement), ii) an Ornstein-Uhlenbeck motion model (OU - Brownian Motion within a home range), and iii) an Ornstein-Uhlenbeck motion model with foraging included (OUF), i. e. assuming regular Brownian motion (while foraging) on a very short time scale, and Ornstein-Uhlenbeck motion on longer time scales. We used the Akaike Information Criterion corrected for small sample size (AICc) to select the best model.
Additionally, to allow comparisons with previous home range estimates for the species, we used the rhr package (rSigner and Balkenhol 2015) in R to estimate 50, 95, 99 and 100 % Minimum Convex Polygon (MCP) home ranges, as well as 50, 95, 99 and 100 % isopleths from a kernel density home range estimate (KDE) with bandwidth set to the reference bandwidth. From the latter calculations, a core home range area was estimated using the method of Seaman and Powell (1990) as provided in the rhr package (Supporting Information 1). To investigate movement rates, we calculated step characteristics using the rhr package and summarized step lengths to indicate movement distances over three time spans: 6, 12, and 24 hours.
Habitat preference. To investigate habitat selection within home ranges (i. e. third-order habitat selection; Johnson 1980), we used the most recent land use/land cover map available in vector format based on 30m-resolution Landsat data, specifically for Belize from the Belize Environmental Resource Data System of Belize (BERDS 2005), representing the 2011 situation with 18 land use/land cover classes. We considered the proportion of each land use/land cover class within the 100% MCP as available, and the proportion of relocations in each class as the proportion of habitat used. We tested for differences between available and used with a χ2-test. Additionally, we calculated the Jacobs index (Jacobs 1974) to detect which specific classes were used more often than expected from their availability in the landscape. The Jacobs index is a modified version of Ivlev's electivity index (Ivlev 1961) that takes into account the relative abundance of a resource when identifying an organism's resource preferences (Jacobs 1974). It is calculated as D = (r - p) / (r + p -2rp), where r is the proportion of habitat used and p is the proportional habitat availability. D varies from -1 under strong avoidance to +1 under strong habitat preference. Values close to 0 indicate that the habitat is used in proportion to its availability.
Results
Trapping success. On April 22nd, 2015, we trapped 12 individuals from a herd of an estimated 60 in the corral trap. The individual selected for collar deployment was an adult female, estimated to weigh just over 25 kg. Fitting the collar took ca. five minutes, after which the animal was released from the handling compartment to join the others in the corral. Camera trap footage showed the collared female among the rest of the herd less than four hours after the trapping event, and with its young, both in good health, two weeks later.
GPS fix rate and data transfer. We report here on data collected between 22 April and 22 August 2015, i. e. 121 days of collar deployment. Data for ca. 47 days did not transfer to the satellite, meaning that 38.7 % of the time we did not know whether a fix attempt had been successful, and where the animal was located. The average length of such data gaps was 6.7 (± 3.0 SD) days, with a maximum of 12 days. From all the fix attempts that were transmitted to the satellite (n = 593), only 11.6 % were successful, thus yielding a total of 69 GPS fixes over the study period.
Home range. The empirical semi-variogram was best approximated by the semi-variance function representing Ornstein-Uhlenbeck motion including foraging (OUF; 2221.51 AICc), and thus showed evidence for home ranging behaviour over the four-month monitoring period (Figure 2). Only on very short time scales did the animal display random, undirected movement (2.53 hours, CI 0.92, 6.91) indicative of foraging. The OU and BM models followed with ∆AICc 8.11 and 1760.48 respectively.
The estimated home range size was 55.2 km2 (CI 34.09, 81.32) which the animals could cover in 3.66 days (CI 1.70, 7.89; Figure 3). The 95 % MCP home range estimate was less than half that of the 95 % KDE isopleth, while the core area estimate based on the KDE was closer to the 95 % MCP (Table 1).
Table 1 Home range size estimates for a single white-lipped peccary herd in southern Belize across different levels.

*Minimum Convex Polygon.
**Kernel Density Estimate.
Movement. Mean distances covered during fix intervals of six, 12 and 24 hours were 190, 910 and 1181 m respectively, while daily distances covered between subsequent fixes with multi-day fix intervals averaged around 900 m (Table 2). The maximum distance covered during fix intervals of up to one day was 2,380 m. For fix intervals longer than one day, the maximum observed speed was 3,788 m/day. Over all fix intervals, the maximum movement speed calculated was 270 m/h.
Table 2 White-lipped peccary step length statistics for different time intervals between GPS fixes.

*statistics per day.
Habitat preference. Comparing the proportion of land use/land cover types for the relocations with the proportion of these classes in the 100 % MCP area, a χ2-test showed significant difference between use and availability (χ2 =15.133 and P < 0.001 based on 2000 Monte Carlo replicates).
Jacobs D indicated a preference for broadleaf forest and a slight avoidance of shrubland (Table 3). Since no relocations were recorded in savannah and agricultural lands, the D index indicates total avoidance (-1.00). Note that we could not calculate significance levels for preferences of individual land cover classes, because our sample size of one does not allow us to calculate the relevant test statistics.

Discussion
Home range, density and population size. Even though white-lipped peccaries have been considered nomadic or migratory in early studies (Kiltie and Terborgh 1983; Bodmer 1990), the current consensus is that they use well-defined home ranges that are large enough to encompass seasonal or multiannual movements (Fragoso 2004; Keuroghlian et al. 2004; Reyna-Hurtado et al. 2009; de Almeida Jácomo et al. 2013). All our home range estimates fall within the range sizes reported for white-lipped peccaries from areas under various disturbance and rainfall regimes in Brazil, Costa Rica, and Mexico (Fragoso 1998, 2004; Carrillo et al. 2002; Keuroghlian et al. 2004; Reyna-Hurtado et al. 2009; de Almeida Jácomo et al. 2013). However, the time span covered by our estimates is four months for a single herd, whereas the other estimates cover multiple herds, each of which was tracked for a period of three months to more than one year. Despite the short time span, our data covers the end of the 2015 dry season and the beginning and peak of the wet season of the same year, and it remains to be seen whether our home range estimates will shift or expand during the transition into the 2016 dry season, until the collar drops off in May 2016. However, although the semi-variogram indicated home ranging behaviour, it is impossible to say whether our location data adequately represent the entire home range, given the numerous gaps in the data. For example, we suspect that the dense canopy in rugged terrain may more severely affect GPS fix success rate and satellite data transfer than relatively flat terrain, which would cause an underestimate of space-use in the more rugged areas.
Nonetheless, within the home range estimated thus far, our habitat preference results agree with the preference for forested areas that is well-known for the species. Given the low availability of agricultural lands inside the MCP area and the limited number of relocations, the total avoidance of agricultural lands remains uncertain. With about 12.5 % of the home range covered by savannah, it is remarkable that no relocations were observed there, while almost 5 % of the relocations were located in shrubland, which was available at half the rate as savannah. This could be explained by i) shrubland providing better cover or food and ii) the patchy spatial configuration of shrubland throughout the forest making movement through these patches more likely than through the one continuous patch of savannah south of the forested areas. However, we emphasize that due to a sample size of one herd, significance testing of preference for specific habitats was not possible.
Due to its robustness with sparse data, we consider the semi-variogram estimate the most accurate for our data set. Using the home range size estimate (and confidence intervals) of this approach and our estimated number of individuals in the herd (n = 60), we would obtain a density of 1.09 (CI 0.74, 1.76) ind/km2. If we i) extrapolate these home range and density estimates to other herds in the Maya Mountains (as delineated by Briggs et al. 2013), and ii) assume that all the broad-leaf forest habitats in the Maya Mountains are suitable for white-lipped peccaries, we estimate 99 (CI 67, 160) herds roaming the area, and a population size of 5,917 individuals (CI 4017, 9582). Clearly, the assumptions for these estimates are untested and the estimates are therefore associated with very high uncertainty; we include them here in the discussion for indicative purposes only. Our density estimate of 1.09 ind/km2 is in the lower half of the range of estimates from the Argentinian Gran Chaco (1.02 ind/km2 Altrichter 2005), the Brazilian Cerrado (2.99 ind/km2 Desbiez et al. 2010) and southern Brazil (4.5 ind/km2 Keuroghlian et al. 2004), but still considerably higher than in the Calakmul Biosphere Reserve in Mexico (0.43 ind/km2 Reyna-Hurtado et al. 2009), which is geographically closest to our study area. However, all of these areas are drier and/or less forested than our study area, which is expected to influence group size and density. For example, the high density in southern Brazil might be due to a crowding effect in the fragmented forests. We expect to have overestimated population size, because not all broad-leaf forests in the Maya Mountains are expected to be used by the species, and hunting pressure in the herd's home range is expected to be lower than in most other parts of the area. Nevertheless, the estimates suggest that the Maya Mountains can potentially harbour a large number of while-lipped peccaries, warranting greater research emphasis and conservation incentives on the species in this region.
Collar performance. The numerous reports of successfully deployed GPS collars in different environmental settings convincingly illustrate the potential that this equipment holds for obtaining large quantities of high quality data in wildlife research (Edenius 1996; Eriksen et al. 2011; Martins et al. 2011; Harju et al. 2013; Krofel et al. 2013). However, the use of GPS collars often comes with unforeseen technical or data quality issues (Sager-Fradkin et al. 2007; Matthews et al. 2013). In particular, studies from tropical forests report imprecise locations or low GPS fix success rate (Phillips et al. 1998; Blake et al. 2001; Lizcano and Cavelier 2004; Coelho et al. 2007; Barlow 2009; Hwang et al. 2010). While we anticipated that the dense forest and steep terrain in our study area would pose a challenge for any telemetry-based research, we opted for GPS collars because i) we wanted to obtain detailed information on movement patterns to parametrize a mathematical model for simulating potential movement paths of peccaries in Belize, and ii) the cost of obtaining a limited number of GPS collars was estimated less than the personnel and equipment costs for running a large camera trap grid or transect network in the inaccessible area. Unfortunately, the fix success rate of 11.6 % was much lower than expected from the literature, even for the given environmental conditions. However, collar malfunction cannot be excluded as a possible explanation. For logistical reasons, we could not perform collar tests in the field prior to deployment, but we stress that this should in fact be standard practice. The poor performance of one collar cannot be used to draw general conclusions about the viability of GPS collars in rugged tropical environments, but we suggest that a review on the usefulness of GPS collars specifically in such environments is needed to help researchers determine a priori the potential in their area. Nonetheless, we are confident that the data gathered during this study outperforms data collection using regular VHF collars in our area and for our highly mobile target species, both in terms of quantity and quality of locations obtained.
Movement. Movement data of high temporal resolution has been, and remains, difficult to obtain for white-lipped peccaries, and hence movement distances of white-lipped peccaries have mostly been described qualitatively in terms of home range behaviour, migration and/or nomadism (Sowls 1997). In this study, the herd of the tagged white-lipped peccary seemed to move about 1 km per day on average, but did at times travel at 270 m/h and covered up to 3,788 m per day, which enabled the herd to traverse its home range in two to eight days. Due to our frequent sampling gaps however, our estimates likely underestimate the mobility of the species in our study area. For comparison, Fragoso (1998) reported distances from 1200 to 2600 m between VHF relocations of unknown time intervals, while Reyna-Hurtado et al. (2009) mentioned a travel speed of up to 3,000 m/h, traversing seasonal home ranges in one-day time intervals.
Based on observations elsewhere in Central and South America (Carrillo et al. 2002; Tobler et al. 2009; Moreira-Ramírez et al. 2015), we expected white-lipped peccary activity to peak during mid-morning and mid-afternoon. However, from our movement data (i. e. speed and distance measures), we were unable to detect any clear diurnal activity patterns (see Appendix 1). We attribute this to the poor fix success rate of the collar. The fix success rate is lowest during the most active periods of the day (Figure 4), leading to a lack of information on movement distance and speed during these periods. This suggests that the behaviour of the animal is potentially an important factor influencing the success of our GPS collar, in line with previous findings (D'Eon and Delparte 2005). The downward trend in fix success rate from the onset of the high activity period also suggests that time since the last successful fix is inversely correlated to the probability of obtaining a successful subsequent fix. The time lag between fix attempts has been found to negatively affect fix success rate in previous studies (Moen et al. 2001; Cain et al. 2005).

Figure 4 Fix success rate (%) was very low in general, but especially so during expected high activity peaks. * relative activity was calculated as the mean relative activity from the datasets presented in Carrillo et al. (2002), Tobler et al. (2009) and Moreira-Ramírez et al. (2015).
Notes on behavior. During the first six months, the animals visited the trap sites a total of five times with an average of 44.4 (± 51.9 SD) days in between. In the next two months, visit frequency increased, with an average of 11 (± 11.9 SD) days between subsequent visits. Note that the coefficient of variation in both cases is greater than one, indicating the unpredictability in time of the trap site visits. This visit frequency was generally lower than in Manu National Park in the upper Amazon basin in Peru, where white-lipped peccaries passed by the same observation site every 4.25 days in the dry season, and every 14.5 days in the wet season (Kiltie and Terborgh 1983). It is unclear whether this difference in visit frequency would be due to differences in food availability, wariness of human presence, or other factors. Other researchers have found similar time spans for the animals to increase trap visit frequency (three months to one year; C. Richard-Hansen pers. comm.).
White-lipped peccaries have been reported in some cases to engage in aggressive behaviour towards a threat (Sowls 1997), but we observed no direct attacks from the animals inside or outside the corral. Four of the trapped animals escaped upon our arrival as they managed to jump the fence using each other's backs as steps, while the remaining eight animals stood their ground and displayed a circular defence formation in the centre of the corral, in agreement with observations by Nietschmann (1972). Once the selected animal was isolated in the handling compartment out of view of the others, it was secured in place and let itself be handled without resistance.
Conclusions and recommendations
To our knowledge, white-lipped peccaries have so far been fitted with VHF-based radio-telemetry collars only, and no studies using GPS-collared white-lipped peccaries have been published. However such studies are being conducted in French Guyana, the Brazilian Pantanal and Mexico (C. Richard-Hansen pers. comm., A. Keuroghlian and R. Reyna-Hurtado, pers. comm.).
Our study illustrates some important challenges faced in trapping white-lipped peccaries and the use of GPS collars in dense tropical forests on rugged terrain. First, the time between the installation of the traps and the trapping success was considerably longer than anticipated. Second, collar performance was very poor in our area, presumably due to animal behaviour, canopy cover and terrain conditions. Recent implementations of the quick fix pseudo-ranging (QFP) system in terrestrial applications could improve the performance (Tomkiewicz et al. 2010). Third, even though we found that collaring without sedation is possible, we have no information on the animal's stress levels while being handled other than the observation that the animal let itself be handled easily once isolated. Nogueira et al. (2015) simulated a hunting situation on captive white-lipped peccaries using a similar trapping set-up as was used in this study. They found that the psychological negative effect (i.e. increased wariness) of the trapping event lasted no longer than eight days. However, the approach also has the disadvantage of not allowing for collecting body measurements or blood samples. For these reasons, we recommend using sedation when possible.
Despite the challenges, the preliminary results provide a much-needed glimpse of the spatial ecology of white-lipped peccaries in Belize. We present notes on behaviour, measurements of maximum and mean daily movement distances, habitat preference and home range estimates. We cautiously provide density and population size estimates, but future studies should focus on estimating peccary densities directly (e. g. via camera-trapping or genetic mark-recapture), assess variability of herd size in Belize, and collar a greater number of individuals. We note that the relocations of our herd are spread over BNR and all five surrounding protected areas, highlighting the necessity to coordinate management of these areas so as to avoid spill-over effects of disturbances from one area to the other. Improving the management of, and knowledge about, the white-lipped peccary populations in the Maya Mountains is crucial for their survival in Belize as well as for monitoring and maintaining the connectivity of the area to neighbouring forests of La Selva Maya and the Calakmul area (Reyna-Hurtado et al. 2009; Moreira-Ramírez et al. 2015).


