










Services on Demand
Journal
Article
 Spanish (pdf)
Spanish (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
-
 Cited by SciELO
Cited by SciELO -
 Access statistics
Access statistics
Related links
-
 Similars in
SciELO
Similars in
SciELO
Share

 Permalink
PermalinkTherya
On-line version ISSN 2007-3364
Therya vol.2 n.3 La Paz Dec. 2011
https://doi.org/10.12933/therya-11-47
Renovando las medidas para evaluar la diversidad en comunidades ecológicas: El número de especies efectivas de murciélagos en el sureste de Tabasco, México
Rodrigo García-Morales1, Claudia E. Moreno1 y Joaquín Bello-Gutiérrez2†
1 Centro de Investigaciones Biológicas. Universidad Autónoma del Estado de Hidalgo. Carretera Pachuca-Tulancingo Km 4.5, Col. Carboneras. Mineral de la Reforma, Hidalgo, México. CP 42184. Teléfono: 01 (771) 71 72 000, ext. 6665. E-mail: r.garciamorales83@gmail.com (RGM), cmoreno@uaeh.edu.mx (CEM)
2 División Académica de Ciencias Biológicas. Universidad Juárez Autónoma de Tabasco.
Sometido: 4 octubre 2011
Revisado: 14 noviembre 2011
Aceptado: 18 noviembre 2011
Resumen
En muchos estudios se analiza y se compara la diversidad de especies, como una propiedad de la estructura de las comunidades, especialmente para sugerir prioridades para la conservación. En años recientes se han propuesto medidas más robustas que pueden ayudar a lograr de mejor manera este objetivo. En este trabajo se evalúa la diversidad de especies "verdadera" con base en la riqueza de especies y el número de especies efectivas de las comunidades. Como estudio de caso se analiza el cambio de la diversidad de murciélagos filostómidos entre zonas de selva y zonas de vegetación secundaria en el sur del estado de Tabasco, México. Tanto la riqueza como la diversidad resultaron ser mayores en los remanentes de selva. En promedio la vegetación secundaria alberga 2.33 especies menos, y el 79.56% de la diversidad de murciélagos de la selva. Una de las principales ventajas del número de especies efectivas es la facilidad con la cual podemos evaluar directamente la magnitud de los cambios en la diversidad entre comunidades.
Palabras clave: Chiroptera, diversidad verdadera, Parque Estatal La Sierra, Phyllostomidae, riqueza de especies, vegetación secundaria.
Abstract
Many studies analyze and compare species diversity, as a property of community structure, mainly in order to suggest priorities for conservation. Recently, robust measures of diversity have been proposed, and they may be helpful to achieve this objective in a better way. In this work we assess "true" species diversity, using species richness and the effective number of species in ecological communities. As a study case we assess the change of phyllostomid bat diversity between forest and secondary vegetation areas in Southern Tabasco, Mexico. Both species richness and species diversity were higher in forest remnants. In average, secondary vegetation areas harbor 2.33 less species, and 79.56% of bat diversity, than forests. One of the main advantages of using effective species numbers is that they allow for a direct evaluation of the amount of change in diversity between communities.
Key words: Chiroptera, true diversity, Parque Estatal La Sierra, Phyllostomidae, species richness, secondary vegetation.
Introducción
Medir la diversidad de especies es un objetivo frecuente en distintas disciplinas biológicas, como la ecología, la biogeografía o la biología de la conservación. Esta medición es necesaria, por ejemplo, para evaluar cuánto cambia la diversidad entre dos o más comunidades (diferentes tipos de hábitat, distintos momentos de tiempo, o a través de gradientes ambientales naturales o antrópicos). La evaluación de la diversidad de especies ha adquirido mayor relevancia en los últimos años debido a su posible relación con el funcionamiento de los ecosistemas, y por su modificación como resultado de actividades humanas (Maclaurin y Sterelny 2008). Además, los cambios en la magnitud de la diversidad pueden servir para justificar acciones de protección de los ecosistemas. Desde el enfoque de la ecología, la diversidad de especies es una propiedad relacionada con la estructura de las comunidades, que puede definirse como el recíproco de un promedio de las abundancias relativas de las especies (Hill 1973). El valor de este recíproco es el número máximo posible de especies que podrían coexistir en una comunidad, si todas ellas tuvieran la misma abundancia, es decir, el número efectivo de especies en la comunidad (Hill 1973; Jost 2006; Tuomisto 2010a, b; 2011; Moreno y Rodríguez 2011; Moreno et al. 2011). La variable más frecuentemente utilizada para medir abundancia de las especies es el número de individuos, pero también se puede emplear la biomasa, el porcentaje de cobertura, o cualquier otra variable cuantitativa de la importancia relativa de las especies en la comunidad. Si todas las especies de una comunidad tuvieran exactamente la misma abundancia, entonces la diversidad sería máxima, y equivaldría al número de especies. Jost (2006) acuño el término diversidad verdadera (true diversity) para referirse de forma particular a medidas matemáticamente robustas que se ajustan a este concepto biológico. Una de las ventajas de expresar la diversidad de una comunidad en números de especies efectivas, es que esta medida permite comparar directamente la magnitud de la diferencia en la diversidad de dos o más comunidades, lo cual no es posible con índices tradicionales de diversidad (Jost 2006, 2007, 2010).
Los murciélagos son un grupo abundante y muy diverso, principalmente en las regiones tropicales, donde llegan a representar localmente hasta el 50% de las especies de mamíferos presentes (Patterson et al. 2003). La familia Phyllostomidae incluye un total de 161 especies, lo que la ubica como una de las familias de mamíferos más diversa en los trópicos (Simmons 2005). Los murciélagos filostómidos han sido utilizados como grupo focal en la evaluación del impacto de la fragmentación y modificación del hábitat sobre las comunidades ecológicas, debido a que las especies responden de manera diferente a las perturbaciones (Galindo-González 2004). Sin embargo, es poco lo que se sabe sobre la ecología de las comunidades de murciélagos en diferentes etapas sucesionales después del abandono de las actividades antrópicas. Este tipo de información puede ser relevante para evaluar el valor que pueden tener las zonas de vegetación secundaria para la conservación de la diversidad de murciélagos, así como el papel que pueden tener los murciélagos en la regeneración de la vegetación de los distintos estadios sucesionales. El objetivo de este trabajo es evaluar cómo cambia la diversidad de murciélagos filostómidos entre zonas de selva y zonas de vegetación secundaria en el sur del estado de Tabasco, México, utilizando números de especies efectivas.
Material y métodos
Área de estudio
El Parque Estatal La Sierra (PELS) se encuentra ubicado al sureste del estado de Tabasco, en los municipios de Tacotalpa y Teapa (92° 38’ y 92° 58’ de longitud oeste, y 17° 25’ y 17° 35’ de latitud norte). Cuenta con una superficie aproximada de 15,000 ha. Está conformado por tres sierras: Madrigal, Poana y Tapijulapa. El PELS presenta una topografía accidentada, con pendientes pronunciadas y altitudes que van desde los 50 a 1,000 msnm. El clima presente es cálido húmedo con lluvias todo el año, con una temperatura media anual superior a los 22 °C y una precipitación que oscila entre los 3,000 y 4,000 mm anuales (SEDESPA 2004).
En la región se encuentra una gran variedad de tipos de vegetación, que van desde la vegetación hidrófita, como popales, tulares, selva baja espinosa de tinto y canacoitales, hasta una amplia superficie ocupada por pastizales y cultivos, que han sustituido a la selva alta perennifolia original de la región (SEDESPA 2004). De manera general se observan tres tipos principales de comunidades vegetales dentro del PELS. El primero es la selva mediana subperennifolia, que se encuentra restringida a las partes accidentas del relieve, en alturas que van de los 100 a 1,000 msnm. Se encuentra constituida por un estrato arbóreo de 20 a 30 m de altura. Las especies predominantes en este tipo de vegetación son el huapaque (Dialium guianense), zapote mamey (Pouteria zapota), chicozapote (Manilkara zapota), cachimbo (Platymiscium yucatanum) y la bellota de montaña (Sterculia mexicana). Dentro del estrato medio, con alturas de 13 a 20 mts, se encuentran especies como el ramón (Brosimum alicastrum), el cedrillo (Guarea bijuga) y el jobo (Spondias mombim). Entre los árboles de 4 a 13 m de altura están el botoncillo (Rinorea guatemalensis), la cascarilla (Croton glabellus), el gogo (Salacia ellipatica) y el popiste (Blepharidium mexicanum; SEDESPA 2004).
La segunda comunidad es la vegetación secundaria, que se deriva de la perturbación de la selva por actividades humanas tales como la extracción forestal y las actividades agrícolas y ganaderas. Se localiza en terrenos planos o semiplanos de baja pendiente. La estructura arbórea va de los cuatro a los 12 m de altura, con especies como el guate (Chlocospermum vitifolium), guarumo (Cecropia obtusifolia), dama de noche (Cestrum noctumun), momo (Piper auritum), majagua (Hampea integerrima) y platanillo (Heliconia bihai, H. latispata).
En el estrato inferior se encuentran especies de herbáceas, trepadoras y algunos arbustos, como la hoja de murciélago (Pasiflora coriacea), papaya chica (Carica mexicana), bola de venado (Thevethia ahouai; SEDESPA 2004). Finalmente, la vegetación inducida está caracterizada por cultivos de maíz, frijol, plantaciones de café y cacao principalmente. El cultivo de arboles maderables ocupa una pequeña superficie del PELS. Los cultivos se encuentran distribuidos en las zonas medias y altas de las serranías. Las partes bajas son ocupadas por pastos para la cría de ganado vacuno (SEDESPA 2004).
Para este trabajo se seleccionaron tres sitios para el monitoreo de las comunidades de murciélagos, en cada una de las tres sierras que conforman el PELS (nueve sitios en total). En cada sitio se consideraron dos tipos de vegetación para los muestreos: selva mediana y vegetación secundaria. En cada uno de los sitios de muestreo los murciélagos fueron capturados con el uso de seis redes de niebla (12 x 3 m). Las cuales se dividieron en los dos tipos de vegetación seleccionados. En cada tipo de vegetación se muestrearon los murciélagos durante tres noches consecutivas, por un periodo de 6 horas a partir del anochecer. Los muestreos se realizaron de octubre de 2004 a noviembre de 2005, acumulando un total de nueve noches de muestreo por cada tipo de vegetación en cada sitio. La identificación de los murciélagos se hizo por medio de la clave de campo de Medellín et al. (1997).
Análisis de los datos
El número de especies efectivas se obtiene con la fórmula:
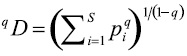
donde: qD es la diversidad verdadera (Jost 2006), pi es la abundancia relativa (abundancia proporcional) de la iésima especie, S es el número de especies, y q es el orden de la diversidad y define la sensibilidad del índice a las abundancias relativas de las especies (Jost 2006, 2007; Tuomisto 2010a, b, 2011). El valor del parámetro q determina qué tanto influyen las especies comunes o las especies raras en la medida de la diversidad, y puede tomar cualquier valor que el usuario estime apropiado (Hill 1973).
Para este trabajo consideramos dos medidas de diversidad verdadera. La primera medida es la diversidad de orden cero (0D), cuyo valor equivale simplemente a la riqueza de especies (0D = S), pues de esta manera la fórmula de la diversidad verdadera es insensible a la abundancia relativa de las especies. La segunda medida es la diversidad verdadera de orden 1 (1D), en la cual todas las especies son consideradas en el valor de diversidad, ponderadas proporcionalmente según su abundancia en la comunidad (Hill 1973; Jost 2006, 2007; Tuomisto 2010a, b, 2011; Moreno et al. 2011). El cálculo directo de la fórmula de diversidad verdadera con valor de q = 1 no es posible, pero se puede obtener sustituyéndolo por valores cercanos a 1 (0.9999 o 1.0001), o bien, - obteniendo el exponencial de índice de entropía de Shannon (Jost 2006):

Para este trabajo, las dos medidas (riqueza de especies y 1D) se calcularon para cada tipo de vegetación (selva y vegetación secundaria) en los nueve sitios de muestreo.
Resultados
Se registró un total de 1,448 individuos de 17 géneros y 27 especies de murciélagos filostómidos en el PELS. Las especies más abundantes fueron Carollia sowelli y Sturnira lilium, con 532 y 173 individuos, respectivamente. Las especies Chiroderma salvini, Glossophaga commissarisi, Lophostoma brasiliense y Trachops cirrhosus sólo registraron un individuo para cada una. El 48% de las especies pertenecen a la subfamilia Stenodermatinae. Las subfamilias menos representadas son Desmodontinae y Carolliinae (Tabla 1).
En todos los sitios, excepto en uno de la sierra de Tapijulapa, la riqueza de especies fue mayor en zonas de selva que en zonas con vegetación secundaria (Fig. 1). Los mismos sitios fueron los que registraron los valores más altos, o más bajos, de riqueza en los dos tipos de vegetación. La mayor riqueza se registró en un sitio de la Sierra de Tapijulapa (19 especies en la selva y 18 en la vegetación secundaria), mientras que el menor número de especies se registró en un sitio de Madrigal y un sitio de Poana (7 especies en la selva de ambos sitios, 5 y 4 en la vegetación secundaria, Fig. 1a). La mayor diferencia entre la zona de selva y de vegetación secundaria (6 especies) se registró en el sitio 2 de Madrigal (Fig. 1a). En promedio, los sitios de vegetación secundaria tienen 2.34 especies menos que los sitios de selva (Fig. 1b).
La diversidad de especies sigue la misma tendencia: en todos los sitios (excepto uno de la sierra de Poana), el número efectivo de especies es mayor en las selvas que en las zonas con vegetación secundaria (Fig. 2). La mayor diversidad en las zonas de selvas se registró en un sitio de Madrigal (9.12 especies efectivas) y para las zonas de vegetación secundaria en un sitio de Poana (7.11 especies efectivas), mientras que la menor diversidad de ambos tipos de vegetación ocurrió en un sitio de Madrigal (4.49 y 3.00 especies efectivas en selva y en vegetación secundaria, respectivamente, Fig. 2a). Al igual que con la riqueza de especies, la mayor diferencia en la diversidad entre la zona de selva y de vegetación secundaria (4.34 especies efectivas) se registró en el sitio 2 de Madrigal (Fig. 2a). Esta diferencia máxima implica que en la selva hay casi el doble de la diversidad que hay en la vegetación secundaria. En promedio, las zonas de selva tienen 1.26 veces más diversidad que las zonas con vegetación secundaria. Lo que equivale a decir que la vegetación secundaria alberga, en promedio, el 79.56% de la diversidad de murciélagos filostómidos que hay en las zonas de selva del PELS (Fig. 2b).
Discusión
Estos resultados indican que las comunidades de murciélagos son menos ricas y menos diversas en estadios sucesionales, que en los remanentes de selva en la región. Sin embargo, mantienen el 80% de la comunidad de murciélagos filostómidos del PELS. Las áreas con vegetación secundaria podrían funcionar como zonas de amortiguamiento para las especies a nivel de paisaje. Por ejemplo, en una zona cercana al PELS, se ha demostrado que la vegetación secundaria mantuvo valores de riqueza y diversidad de murciélagos similares a la selva (Castro-Luna et al. 2007). Esta situación se presenta en otros grupos biológicos, por ejemplo, para las comunidades de plantas la riqueza y diversidad de especies es mayor en zonas de vegetación secundaria que en zonas de selva, posiblemente por la heterogeneidad espacial de los diferentes estadios regenerativos después del abandono de las actividades humanas (Castillo-Campos et al. 2008). Sin embargo, además de la alta diversidad de plantas, se ha documentado también una elevada tasa de cambio en la composición florística entre sitios (Trejo y Dirzo 2002; Trejo 2005) y particularmente entre zonas de selva y zonas de vegetación secundaria (Castillo-Campos et al. 2008). Esto podría sugerir que para los murciélagos de la familia Phyllostomidae, que en su mayoría se alimentan de frutos, néctar y polen, al parecer una estructura compleja de las comunidades no depende tanto del número de especies de plantas, sino de la composición florística. La composición florística de los remanentes de selva del sur de Tabasco incluye especies que proveen una gama de recursos más amplia para los murciélagos filostómidos, que la que podría ofrecer la vegetación secundaria. Por ejemplo, especies como Brosimum alicastrum, Spondias mombim son características de los remanentes de selva y son consumidas por los murciélagos.
A nivel local, en algunos sitios puntuales la variación en la diversidad de murciélagos entre la zona de selva y la zona de vegetación secundaria es diferente según se incluya o no información de la abundancia proporcional de las especies. La riqueza de especies (0D) es el extremo de un continuo de medidas de diversidad (Moreno y Rodríguez 2011). En las medidas de diversidad con valores positivos del parámetro q (0.5D, 1D, 2D, etc.) se incluye, además del número de especies, las abundancias proporcionales de las especies, dando cada vez menos peso a las especies raras conforme aumenta el valor de q (Hill 1973; Jost 2006, 2007; Tuomisto 2010a, b, 2011; Moreno et al. 2011). Sin embargo, a nivel regional, la tendencia de cambio en las comunidades de murciélagos filostómidos entre zonas de selva y zonas de vegetación secundaria es similar, tanto con la riqueza de especies como con 1D.
El número de especies efectivas es el número de especies que tendría una comunidad virtual en la que todas las especies fueran igualmente comunes, conservando la abundancia relativa promedio de la comunidad estudiada (Jost 2006). El concepto de esta unidad de medición de la diversidad es mucho más fácil de entender en términos biológicos, que unidades como los bits o los nats del índice de entropía de Shannon (Moreno et al. 2011). Pero más aún, una ventaja importante del número de especies efectivas es que se puede evaluar directamente la magnitud de cambio entre comunidades. Por ejemplo, en estos resultados, en el sitio dos de la sierra de Madrigal la diferencia entre 9.12 y 4.78 especies efectivas significa que la selva tiene 1.91 veces más diversidad que la vegetación secundaria. Lo que podría implicar que en la vegetación secundaria hay una pérdida del 47.59% de la diversidad de murciélagos de la selva. Estas relaciones directas con el índice de entropía de Shannon no son posibles porque este índice no tiene un comportamiento lineal y daría un valor distorsionado de la diferencia real: una entropía de 2.21 y 1.56 nats señala equivocadamente que en la vegetación secundaria se ha perdido sólo el 29.23% de la diversidad de murciélagos de la selva, si dicha diversidad se midiera con el índice de entropía de Shannon. Por ello, la evaluación de la diversidad a través del número de especies efectivas constituye un método mucho más valioso que otros índices tradicionalmente utilizados como medidas de diversidad, especialmente para la comparación entre comunidades, que constituye un objetivo de trabajo muy frecuente en ecología y biología de la conservación.
Con el propósito de dar énfasis en la evaluación directa de la magnitud de cambio en la diversidad entre comunidades, en este trabajo no se ha incluido una evaluación de la significancia estadística de los cambios en la diversidad. Para muchos objetivos de investigación, manejo y conservación, puede ser interesante saber si la diversidad de una comunidad es significativamente distinta de la diversidad de otra comunidad en un sentido estadístico. Pero casi siempre resulta más importante saber cuánto cambia la diversidad, cuánto se pierde, o cuánto se gana. Sin embargo, la significancia estadística entre números efectivos de especies se puede realizar con pruebas de hipótesis o mediante intervalos de confianza siguiendo los métodos descritos en Moreno et al. (2011).
Es importante resaltar que el uso de medidas verdaderas de diversidad constituye un marco conceptual y metodológico integrador, con muchas más aplicaciones en biología. Por ejemplo, el número efectivo de especies permite dividir la diversidad regional (diversidad gamma) en dos componentes independientes: la diversidad alfa de cada unidad de muestreo, y la diversidad beta o cambio en la composición de especies entre unidades (Jost 2007, 2010; Baselga 2010). El componente beta de la diversidad verdadera puede también aplicarse como una medida matemáticamente consistente para describir la diferenciación genética entre poblaciones (Jost 2008). Finalmente, es de gran relevancia también que este marco general pueda extenderse a otras facetas del concepto amplio de la biodiversidad. Las medidas de diversidad utilizadas para los murciélagos del sur de Tabasco en este trabajo son neutrales en el sentido de que asumen que todas las especies son idénticas. Recientemente Chao et al. (2010) han derivado un grupo de medidas de diversidad verdadera que toman en cuenta tanto la abundancia de las especies como sus relaciones evolutivas (sus diferencias taxonómicas o filogenéticas). Una integración de conocimiento sobre la diversidad de especies, la diversidad evolutiva y/o la diversidad de funciones ecológicas, sin duda mejorará nuestro conocimiento de la estructura de las comunidades de murciélagos y sus cambios a consecuencia de las actividades humanas, y podría generará mejores propuestas para su conservación.
Agradecimientos
E. J. Gordillo-Chávez, G. Ávila, C. Martínez, M. Méndez, R. Carrera, L. M. López por su entusiasta participación en el trabajo de campo. A los guías que nos acompañaron durante los muestreos. A las autoridades civiles de cada uno de los sitios de trabajo. Al Proyecto Aprovechamiento potencial de mamíferos por comunidades asentadas en áreas protegidas de Tabasco Clave: TAB-2003 C02-11255 del cual se derivó la tesis de licenciatura del primer autor. Los análisis de datos y la redacción de este trabajo se realizaron con el apoyo del proyecto CONACYT Ciencia Básica 84127.
Referencias
Baselga, A. 2010. Multiplicative partition of true diversity yields independent alpha and beta components; additive partition does not. Ecology 91:1974–1981. [ Links ]
Castillo-Campos, G., G. Halffter, y C. E. Moreno. 2008. Primary and secondary vegetation patches as contributors to floristic diversity in a tropical deciduous forest landscape. Biodiversity and Conservation 17:1701–1714. [ Links ]
Castro-Luna, A., V. J. Sosa, y G. Castillo-Campo. 2007. Bat diversity and abundance associated with the degree of secondary succession in a tropical forest mosaic in south-eastern Mexico. Animal Conservation 10: 219-228. [ Links ]
Chao A., Ch-H. Chiu, y L. Jost. 2010. Phylogenetic diversity measures based on Hill numbers. Philosophical Transactions of the Royal Society of London B 365:3599–3609. [ Links ]
Galindo-González, J. 2004. Clasificación de los murciélagos de la región de los Tuxtlas, Veracruz, respecto a su respuesta a la fragmentación del hábitat. Acta Zoológica Mexicana 20:239–243. [ Links ]
Hill, M. O. 1973. Diversity and evenness: a unifying notation and its consequences. Ecology 54:427–432. [ Links ]
Jost, L. 2006. Entropy and diversity. Oikos 113:363–375. [ Links ]
Jost, L. 2007. Partitioning diversity into independent alpha and beta components. Ecology 88:2427–2439. [ Links ]
Jost, L. 2008. GST and its relatives do not measure differentiation. Molecular Ecology 17:4015–4026. [ Links ]
Jost, L., P. DeVries, T. Walla, H. Greeney, A. Chao, y C. Ricotta. 2010. Partitioning diversity for conservation analyses. Diversity and Distributions 16:65–76. [ Links ]
Maclaurin, J.,y K. Sterelny. 2008. What is biodiversity? The University of Chicago Press, Chicago, Illinois. [ Links ]
Medellín, R. A., H. T. Arita, y O. Sánchez-Hernández. 1997. Identificación de los Murciélagos de México: clave de campo. Publicaciones Especiales No. 2, Asociación Mexicana de Mastozoología, A.C. México, Distrito Federal. [ Links ]
Moreno, C. E., y P. Rodríguez. 2011. Commentary: Do we have a consistent terminology for species diversity? Back to basics and toward a unifying framework. Oecologia 167:889-892. [ Links ]
Moreno, C. E., F. Barragán, E. Pineda, y N. P. Pavón. 2011. Reanalizando la diversidad alfa: alternativas para interpretar y comparar información sobre comunidades ecológicas. Revista Mexicana de Biodiversidad 82:1249-1261. [ Links ]
Patterson, B. D., M. R. Willig, y R. Stevens. 2003. Trophic strategies, niche partitioning and patterns of ecological organization. Pp. 536–579 in Bat Ecology (Kunz, T. H., y M. B. Fenton, eds.). University of Chicago Press, Chicago, Illinois. [ Links ]
SEDESPA (Secretaria de Desarrollo Social y Protección Ambiental-Gobierno del estado de Tabasco). 2004. Programa rector único de gestión, Programa de manejo del Parque Estatal de la Sierra de Tabasco. (Sierra de Poana, sierra de Tapijulapa y sierra Madrigal). Universidad Juárez Autónoma de Tabasco, División Académica de Ciencias Biológicas, Gobierno del estado de Tabasco y Secretaría de Desarrollo Social y Protección Ambiental. Villahermosa, Tabasco. [ Links ]
Simmons, N. B. 2005. Orden Chiroptera. Pp. 312-529. In Mammal Species of the World: a taxonomic and geographic reference (Wilson D. E. y D. M. Reeder, eds.). Johns Hopkins University press, Baltimore, Maryland. [ Links ]
Trejo, I., y R. Dirzo. 2002. Floristic diversity of Mexican seasonally dry tropical forests. Biodiversity and Conservation 11:2063–2048. [ Links ]
Trejo, I. 2005. Análisis de la diversidad de la selva baja caducifolia en México. Pp. 111–122 in Sobre diversidad biológica: el significado de las diversidades alfa, beta y gamma (Halffter, G., J. Soberón, P. Kolef, y A. Melic, eds.). M3M: Monografías Tercer Milenio, vol. 4. Sociedad Entomológica Aragonesa, Zaragoza, España. [ Links ]
Tuomisto, H. 2010a. A diversity of beta diversities: straightening up a concept gone awry. Part 1. Defining beta diversity as a function of alpha and gamma diversity. Ecography 33:2–22. [ Links ]
Tuomisto, H. 2010b. A consistent terminology for quantifying species diversity? Yes, it does exist. Oecologia 164:853–860. [ Links ]
Tuomisto, H. 2011. Commentary: do we have a consistent terminology for species diversity? Yes, if we choose to use it. Oecologia 167:903-911. [ Links ]
Notas
Editor asociado: Miguel Briones.
Diseño gráfico editorial: Gerardo Hernández.

