











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
En México, una de las principales amenazas de la biodiversidad es la introducción de especies exóticas, ya sea de manera intencional o accidental, las cuales desarrollan un comportamiento invasivo y logran así desplazar a las nativas.
El borrego berberisco (Ammotragus lervia Pallas, 1777) fue introducido al norte y centro del país con fines de aprovechamiento. Actualmente, se encuentra en 38 Unidades de Manejo para la Conservación de la Vida Silvestre (UMA); sin embargo, en las serranías de algunas entidades federativas como Sonora, Coahuila, Nuevo León y Chihuahua se reúnen las principales poblaciones ferales (Medellín, 2005).
Una de las formas para saber cómo afectan las especies exóticas a las nativas, es mediante el conocimiento de la similitud y preferencia de la dieta. Al respecto, se han realizado investigaciones entre especies exóticas y nativas (Barrett, 1967; Tapia y Mellink, 1989; Jackley, 1991; Gastelum, 2020; Gastelum-Mendoza et al., 2023), específicamente, en la composición botánica de la dieta del borrego berberisco y del cimarrón (Medellín, 2005; Guerrero-Cárdenas et al., 2016, 2018; Gastelum-Mendoza et al., 2023).
La cobertura vegetal, como un elemento del hábitat que aporta alimento y distintos tipos de cobertura a la fauna silvestre, es un elemento fundamental cuya importancia varía en función de los requerimientos de la especie (Ramírez, 2004; Fulbright y Ortega-Santos, 2006). En el norte de México y suroeste de Estados Unidos de América, el borrego cimarrón (Ovis canadensis Shaw, 1804) habita en áreas con climas áridos y topografía accidentada que se caracterizan por tener cobertura de altura baja, que brinda una adecuada visibilidad y terreno de escape (Tarango et al., 2002; Escobar-Flores et al., 2015). Asimismo, el borrego berberisco en el norte de África está adaptado a un clima árido extremo, en sierras con baja cobertura vegetal (Nowak, 1991).
La composición botánica de la dieta sirve como instrumento para evaluar y conocer la compatibilidad que existe entre la fauna silvestre y su hábitat. El conocimiento de la composición botánica y nutricional de la dieta es esencial para establecer la capacidad de carga en los planes de manejo de las UMA (Villarreal-Espino-Barros et al., 2008). Existe un buen número de estudios sobre las dietas de ambas especies, pero en muy pocos se aborda la competencia entre ambas, y los que existen se han realizado en los Estados Unidos de América (Smith y Krausman, 1988; Etchart et al., 2016).
El borrego berberisco puede competir por los recursos alimenticios con las especies nativas del noreste mexicano que se alimentan de pastos y herbáceas, y por ello ejercen una presión sobre sus poblaciones, lo cual modifica la dinámica poblacional de ambos grupos (plantas y animales), por ejemplo a las especies de cérvidos y al borrego cimarrón, en particular.
El presente estudio tiene como objetivo, en primer lugar, estimar las dietas de ambos borregos, obtener el grado de similitud de su dieta y finalmente, saber si hay competencia por alimento entre las dos especies.
Materiales y Métodos
Área de estudio
El estudio se realizó en el periodo 2021-2023 en la Unidad de Manejo para la Conservación de la Vida Silvestre (UMA) Rancho San Juan, localizada en las coordenadas: 26°49´12.34” latitud norte y 100°58´43.52” longitud oeste (Figura 1), la cual se localiza a 42 kilómetros en línea recta al este de Monclova, Coahuila, México.
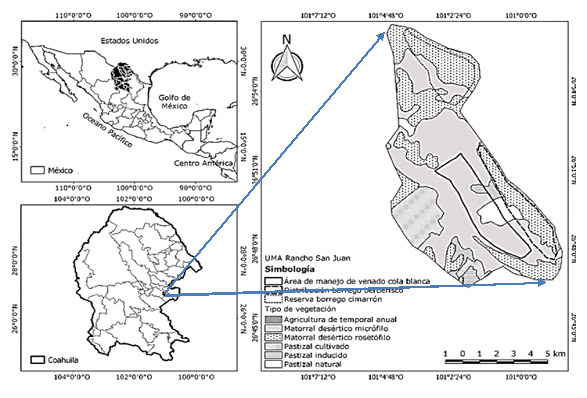
En esta UMA se ubica la sierra Las Hormigas, lugar donde habitan y se desarrollan poblaciones introducidas procedentes de Sonora y Texas de borrego cimarrón y de borrego berberisco, respectivamente en semicautiverio, con las mismas condiciones de hábitat y tipos de vegetación. La UMA tiene una superficie de aproximadamente 4 500 ha y la sierra Las Hormigas tiene una superficie aproximada de 1 200 ha para el berberisco y alrededor de 500 ha para el cimarrón, ambos separados solo por un cerco de 2.8 m de alto, su altitud es de 1 100 msnm en las partes más altas. La vegetación dominante corresponde a un matorral desértico rosetófilo y un chaparral (Gastelum, 2020).
Técnica microhistológica
Para la identificación de las plantas que componen la dieta de los bóvidos, se utilizó la técnica microhistológica (Peña-Neira y Habib de Peña, 1980; Catán et al., 2003) sobre las especies recolectadas del matorral desértico rosetófilo y del chaparral, la cual permite comparar la presencia de residuos vegetales en la materia fecal a través de un catálogo fotográfico de referencia que muestra las estructuras epidérmicas microscópicas de cada planta; con dicha información se generaron los datos que se analizaron estadísticamente.
Los datos en campo se obtuvieron mediante la recolecta de las muestras fecales frescas, aproximadamente 70 por especie de borrego, en cada una de las estaciones del año.
En los análisis estadísticos del presente estudio se utilizó el programa Past 3 (Hammer et al., 2001). A partir de la frecuencia acumulada, se estimó la contribución que tiene cada taxon vegetal en la dieta (Peña-Neira y Habib de Peña, 1980).
Sobre esta base (FA) y el número total de campos analizados (N), se calculó la frecuencia (F) con la siguiente ecuación:
Donde:
F = Frecuencia
FA = Frecuencia acumulada
N = Número total de campos analizados
Una vez conocida la frecuencia, se determinó la densidad relativa (%) y el resultado de este parámetro se utilizó para expresar la composición botánica de la dieta. El cálculo se hizo por estación del año, mediante la siguiente ecuación (Peña-Neira y Habib de Peña, 1980):
Donde:
DRa = Densidad relativa de la especie
D a = Densidad de la especie
Di = Densidad de cada una de las especies
Para conocer la competencia alimentaria existente entre las especies estudiadas se aplicó el Índice de Sørensen y el Índice de Jaccard (Alanís et al., 2020). Para el Índice de Sørensen se usó la fórmula siguiente:
Donde:
I s = Índice de similitud de Sørensen
a y b = Número de especies en las muestras a y b, respectivamente
c = Número de especies compartidas por las dos muestras
El Índice de Jaccard es muy similar al coeficiente de similitud de Sørensen para datos cualitativos; sin embargo, no se relaciona con las especies sino con sus abundancias, su cálculo se hizo con la siguiente ecuación (Alanís et al., 2020):
Donde:
I j = Índice de Jaccard
a = Número de especies en el sitio A
b = Número de especies en el sitio B
c = Número de especies presentes en ambos sitios A y B, es decir, que están compartidas
El intervalo de este Índice varía desde cero (0 % de competencia) cuando no hay especies compartidas, hasta uno (100 % de competencia) si las dos dietas comparten las mismas especies; mide diferencias en la presencia o ausencia de especies.
Índice de Similitud de las Dietas
Para la determinación de la similitud y competencia entre dietas, se utilizó el Índice de Kulcynski (Saiz, 1980), el cual en su inicio se usó para observar el grado de semejanza entre comunidades vegetales, pero también se emplea para conocer la superposición de las dietas. A continuación se presenta la fórmula para su cálculo:
Donde:
IS = Índice de Similitud
W = Porcentaje menor de una determinada planta cuando se comparan sus porcentajes de consumo por dos animales diferentes
(a+b) = Es la suma de estos porcentajes
Resultados
Dieta de las dos especies de borregos
Composición de dieta del borrego berberisco. El primer lugar de consumo anual lo ocuparon las arbustivas con un promedio de 39.87 %, seguidas de los pastos con 28.02 %, las herbáceas con 20.68 %, las especies arbóreas con 6.69 % y finalmente las plantas suculentas con 4.74 % (Cuadro 1).
Cuadro 1 Principales especies con su porcentaje en la composición de la dieta del borrego berberisco (Ammotragus lervia, Pallas 1777) en cada estación del año en el Rancho San Juan, Monclova, Coahuila, México.
| Especie/Estación | Primavera (%) |
Verano (%) |
Otoño (%) |
Invierno (%) |
|---|---|---|---|---|
| 1. Acacia berlandieri Benth. | 4.97 | 1.85 | ||
| 2. Acacia rigidula Benth. | 12.07 | 10.6 | 9.61 | 18.23 |
| 3. Aloysia macrostachya (Torr.) Moldenke | 16.1 | 3.65 | 3.8 | 8.48 |
| 4. Aristida adscensionis L. | 16.1 | 9.98 | 11.31 | |
| 5. Bothriochloa saccharoides (Sw.) Rydb. | 2.04 | 6.87 | 1.39 | |
| 6. Bouteloua curtipendula (Michx.) Torr. | 1.98 | 4.97 | 4.8 | 3.8 |
| 7. Bouteloua gracilis (Kunth) Lag. ex Griffiths | 11.3 | 14.47 | 21.06 | 8.48 |
| 8. Caesalpinia mexicana A. Gray | 1.31 | 4.97 | 4.8 | 6.34 |
| 9. Castela texana (Torr. & A. Gray) Rose | 10.53 | 2.04 | 3.8 | |
| 10. Chamaecrista greggii (A. Gray) Pollard ex A. Heller | 6.83 | 17.95 | 1.85 | 7.94 |
| 11. Euphorbia antisyphilitica Zucc. | 3.32 | 6.87 | 2.33 | |
| 12. Euphorbia polycarpa Benth. | 8.18 | 6.87 | ||
| 13. Forestiera angustifolia Torr. | 0.65 | 3.11 | 0.46 | |
| 14. Larrea tridentata (DC.) Coville | 1.98 | 6.43 | 4.8 | |
| 15. Leucophyllum frutescens (Berland.) I. M. Johnst. | 0.65 | 0.46 | 0.46 | |
| 16. Medicago sativa L. | 1.31 | 3.31 | 5.3 | |
| 17. Opuntia microdasys (Lehm.) Pfeiff. | 1.98 | 6.35 | 9.59 | |
| 18. Tecoma stans (L.) Juss. ex Kunth | 0.65 | 1.02 | 10.17 | 0.46 |
Las especies más importantes por su consumo anual en la dieta del berberisco y el estrato al que pertenecen fueron: de los pastos, Bouteloua gracilis (Kunth) Lag. ex Griffiths (13.82 %); arbustivas, Acacia rigidula Benth. (12.63 %); entre las herbáceas, Aloysia macrostachya (Torr.) Moldenke (8.01 %); de las suculentas, Opuntia microdasys (Lehm.) Pfeiff. (4.48 %) fue la más importante; Tecoma stans (L.) Juss. ex Kunth (3.08 %) sobresalió en el componente arbóreo.
Los taxa del estrato arbustivo ocuparon, en casi todas las estaciones del año, el primer lugar del material vegetal consumido, solo en el otoño fueron sustituidos por las herbáceas, lo que pudo deberse a que esta es la época de lluvias y en el área las primeras especies en proliferar fueron las herbáceas, mismas que fueron consumidas, principalmente, por los borregos berberiscos.
Composición de dieta del borrego cimarrón. Como se indicó previamente con el borrego berberisco, el primer lugar de consumo anual correspondió a las arbustivas, con un promedio de 39.30 %, seguidas de las arbóreas con 20.55 %, el tercer lugar fue para las suculentas con 17.46 %, para los pastos fue el cuarto lugar con 15.44 % y finalmente las herbáceas con 7.25 % (Cuadro 2).
Cuadro 2 Principales especies en la composición de la dieta del borrego cimarrón (Ovis canadensis, Shaw 1804) durante las cuatro estaciones del año en el Rancho San Juan, Monclova, Coahuila, México.
| Especie/Estación | Primavera (%) |
Verano (%) |
Otoño (%) |
Invierno (%) |
|---|---|---|---|---|
| 1. Acacia berlandieri Benth. | 1.77 | 7.43 | 8.97 | 4.62 |
| 2. Acacia rigidula Benth. | 22.36 | 12.22 | 21.22 | 16.66 |
| 3. Acourtia runcinata (D. Don) B. L. Turner | 2.35 | - | 3.05 | 0.75 |
| 4. Agave L. sp. | 0.59 | 4.65 | 10.37 | 2.28 |
| 5. Aloysia macrostachya (Torr.) Moldenke | 0.59 | 4.65 | - | 1.51 |
| 6. Bouteloua adscensionis L. | 1.77 | 8.76 | - | 6.23 |
| 7. Bouteloua curtipendula (Michx.) Torr. | 8.74 | 3.86 | 8.29 | 10.4 |
| 8. Bouteloua gracilis (Kunth) Lag. ex Griffiths | 0.59 | 1.52 | 1.81 | 0.75 |
| 9. Castela texana (Torr. & A. Gray) Rose | 1.17 | 1.52 | 0.6 | 1.51 |
| 10. Croton incanus Kunth | 0.59 | 4.65 | 6.93 | 2.28 |
| 11. Dasylirion berlandieri S. Watson | 3.59 | 0.76 | 1.2 | 3.83 |
| 12. Euphorbia antisyphilitica Zucc. | 4.21 | 5.46 | 0.6 | 2.28 |
| 13. Forestiera angustifolia Torr. | 1.17 | 3.05 | - | - |
| 14. Gochnatia hypoleuca (DC.) A. Gray | - | 4.65 | 5.88 | 10.4 |
| 15. Guaiacum angustifolium Engelm. | 2.97 | 2.29 | - | 0.75 |
| 16. Larrea tridentata (DC.) Coville | - | 2.29 | 2.41 | 2.28 |
| 17. Leucophyllum frutescens (Berland.) I. M. Johnst. | 2.97 | 1.52 | 1.81 | 4.62 |
| 18. Opuntia engelmannii Salm-Dyck ex Engelm. | - | 6.27 | - | 1.51 |
| 19. Opuntia leptocaulis DC. | 2.97 | 4.65 | 0.6 | 4.62 |
| 20. Opuntia microdasys (Lehm.) Pfeiff. | 4.84 | 11.34 | 5.88 | 3.03 |
| 21. Tecoma stans (L.) Juss. ex Kunth | 21.52 | 4.65 | 12.53 | 12.14 |
Para el borrego cimarrón, las principales especies consumidas anualmente y su estrato correspondiente fueron: Acacia rigidula (18.12 %) perteneciente a las arbustivas, Tecoma stans (12.71 %) a las arbóreas, Bouteloua curtipendula (Michx.) Torr. (7.82 %) a los pastos, Opuntia microdasys (6.27 %) fue la más importante de las suculentas y Croton incanus Kunth (3.61 %) del estrato herbáceo.
Este borrego también mostró una mayor preferencia por las arbustivas, las cuales ocuparon, en casi todas las estaciones del año, el primer lugar del material vegetal consumido; sin embargo, las arbóreas tuvieron un lugar importante en su dieta.
El berberisco mostró una mayor preferencia por Bouteloua gracilis y Acacia rigidula, mientras que el cimarrón por Acacia rigidula y Tecoma stans. El resto de las especies, mientras el borrego berberisco las consumió en altas cantidades; el cimarrón lo hizo en bajas cantidades, de tal forma que hay una competencia menor entre ellas (Figura 2).

Comparación y similitud de dietas entre borregos
Índice de similitud de dietas (índices de Sørensen y Jaccard). Los resultados para el Índice de Sørensen mostraron que la competencia mayor entre ambas especies se presentó en invierno con 65 % y que el valor más bajo de competencia fue en otoño con 41 %. Esto lo confirma el Índice de Jaccard para la época invernal con un resultado de 48 % de las plantas registradas en el área de estudio. La estación del año con el menor grado de competencia fue el otoño con 26 %. Mediante este índice se determinó que la competencia es baja entre ambas especies, con base en los valores de referencia.
Lo anterior puede explicarse por el hecho de que la época de lluvias ocurre en el otoño para el noreste de México, por lo que se presentó mayor producción de biomasa en el área de estudio, como lo son las especies de herbáceas y pastos, mientras que en el verano, se registró baja presencia de taxones de este tipo de estrato, lo que provocó que los borregos tuvieran más consumo y preferencia sobre ejemplares de arbustos y árboles, ya que fueron las plantas que se mantuvieron presentes y con mayor disponibilidad en el hábitat; ello dio lugar a mayor competencia por su consumo entre ambos mamíferos.
Similitud alimentaria de acuerdo con el Índice de Kuczynski
A continuación, se muestra la lista de especies en la dieta de ambos borregos, así como en la estación cuando se registró su consumo y la similitud (Cuadro 3).
Cuadro 3 Porcentajes de similitud en la dieta de algunas plantas entre el borrego berberisco (Ammotragus lervia, Pallas 1777) y el borrego cimarrón (Ovis canadensis, Shaw 1804) en el Rancho San Juan, Monclova, Coahuila, México.
| Especie/Estación | Primavera % |
Verano % |
Otoño % |
Invierno % |
|---|---|---|---|---|
| 1. Acacia berlandieri Benth. | - | 80 | 34 | - |
| 2. Acacia rigidula Benth | - | 93 | 62 | 96 |
| 3. Aloysia macrostachya (Torr.) Moldenke | 7 | 88 | - | 30 |
| 4. Aristida adscensionis L. | 20 | 93 | - | 71 |
| 5. Bouteloua curtipendula (Michx.) Torr. | 37 | 87 | 73 | 54 |
| 6. Bouteloua gracilis (Kunth) Lag. ex Griffiths | 10 | 19 | 16 | 16 |
| 5. Castela texana (Torr. & A. Gray) Rose | 20 | 85 | - | 57 |
| 6. Chamaecrista greggii (A. Gray) Pollard ex A. Heller | - | 8 | - | 32 |
| 7. Euphorbia antisyphilitica Zucc. | 88 | - | 16 | 99 |
| 8. Forestiera angustifolia Torr. | 71 | 99 | - | - |
| 9. Gochnatia hypoleuca (DC.) A. Gray | - | - | - | 37 |
| 10. Larrea tridentata (DC.) Coville | - | 53 | 67 | - |
| 11. Leucophyllum frutescens (Berland.) I. M. Johnst. | 36 | - | 41 | 18 |
| 12. Opuntia microdasys (Lehm.) Pfeiff. | 82 | - | 96 | 48 |
| 13. Tecoma stans (L.) Juss. ex Kunth | 6 | 36 | 90 | 7 |
| 14. Ziziphus obtusifolia (Hook. ex Torr. & A. Gray) A. Gray | 71 | - | - | 90 |
| Media por estación (%) | 40.76 | 64.98 | 54.97 | 51.08 |
Para evaluar dicha competencia se aplicó el Índice de Kuczynski, y se determinó la similitud entre dietas por especie y por estación; posteriormente, se evaluó el porcentaje de similitud para cada una de ellas, así como el porcentaje anual.
Los taxones con mayor similitud en la dieta entre ambos borregos en las diferentes estaciones del año fueron Euphorbia antisyphilitica Zucc. para primavera, además registró el más alto porcentaje de similitud, con un valor de 88 %; Forestiera angustifolia Torr. con 99 % en verano; Opuntia microdasys con 96 % en otoño; y Acacia rigidula con 96 % en invierno. Estas fueron las especies más representativas en cada una de las estaciones con base en su porcentaje de similitud en consumo (Cuadro 3). Se obtuvo una similitud anual entre dietas de ambos borregos con un valor de 52.95 %, lo que representa una competencia de media a alta.
Con base en los resultados para el Índice de Kuczynski, se determinó que el porcentaje más alto de competencia por estación del año ocurrió en verano, con un valor de 64.98 %; y el porcentaje más bajo, se presentó en primavera, con un valor de 40.76 %. El taxón por el que compitieron ambas especies de borrego durante todo el año fue el chaparro prieto (Acacia rigidula), ya que para el berberisco ocupó el segundo lugar y para el cimarrón el primer lugar de su dieta (Figura 2). El porcentaje promedio anual de competencia fue de 52.95 %; por lo tanto, se determinó que el grado de competencia anual entre ambas especies se cataloga de media a alta.
Discusión
Las especies más importantes con respecto a sus características ecológicas en el área de distribución del borrego berberisco, en orden de importancia, fueron las del estrato arbustivo y el herbáceo. Los valores de los índices mostraron que existe una gran cantidad de ellas en el área con diferentes grados de dominancia, ya que todas registraron distintos niveles de abundancia, y solo una parte fueron las más representativas.
La variabilidad o disponibilidad de las especies pudo afectarse por las épocas de lluvia y sequía. Esto coincide con lo descrito por Guerrero-Cárdenas et al. (2016) sobre la composición y selección de la dieta del borrego cimarrón, cuando analizaron los patrones de uso, así como la disponibilidad de las especies vegetales presentes, estacionalmente, en el área; se identificaron 63 especies en 2010 y 50 en 2011. Las arbustivas fueron las formas de vida dominantes; las cifras más altas de abundancia de especies en el área se registraron en otoño de 2010 e invierno de 2011.
En el presente estudio, las arbustivas ocuparon el primer lugar con casi 40 % de la dieta del borrego berberisco; por lo que esta especie de borrego es más ramoneadora que apacentadora en el norte de México, en comparación con el borrego berberisco en el Parque Nacional Bou Hedma de Túnez, en donde tiene una clara preferencia por los pastos (67 % de la dieta anual), seguida de arbustivas (17 %) y herbáceas (16 %), lo que identifica a este ungulado como un apacentador (Ben y Nouira, 2015). En la región de Edwards Plateau de Texas, su dieta anual consistió en 61 % de pastos, 21 % de arbustivas y 18 % de herbáceas; lo que confirma su carácter de apacentador en los Estados Unidos de América (Ramsey y Anderegg, 1972).
Las arbustivas en todas las estaciones del año fueron las más consumidas, con porcentajes significativos para ambos borregos; ello se asemeja con lo propuesto por Gastelum-Mendoza et. al. (2023), en el sentido de que los arbustos son la forma de vida más importante en la estructura de la vegetación del hábitat del borrego cimarrón en el noreste de México. Guerrero-Cárdenas et al. (2016) también asumen que las arbustivas son las más consumidas, ya que constituyen 62.1 % de su dieta. Según Hanley (1982), sus grandes mandíbulas y molares, así como su gran volumen rumino-reticular son adaptaciones para el consumo de pastos con alto contenido de celulosa. Sin embargo, Gastelum (2020) observó para el hábitat del borrego cimarrón del desierto en el noreste de México, dominado por comunidades arbustivas con una cobertura de pastos muy escasa, que su dieta se compone por arbustos, subarbustos, herbáceas y árboles, y en menor proporción de pastos (Miller y Gaud, 1989; Gastelum, 2015, 2020; Gastelum-Mendoza et al., 2023).
Es importante subrayar que las arbustivas ocuparon el primer lugar en la dieta de las dos especies de borrego estudiadas, que coincide con lo propuesto por Medellín (2005), quien señala que la dieta del borrego berberisco está compuesta, principalmente, por arbustos pequeños, pastos y plantas herbáceas, y que es capaz de levantarse sobre sus patas traseras para ramonear el follaje de los árboles de tamaño reducido.
Conclusiones
Las dos especies de ungulados silvestres estudiadas presentan una composición muy similar en su dieta anual, pues ambos borregos tienen una preferencia por especies de los estratos arbustivo y arbóreo.
De acuerdo con el análisis de competencia alimentaria, el borrego berberisco y el cimarrón muestran un nivel de competencia de media a alta.
Las estaciones en las que registran más competencia alimentaria son invierno y otoño, con base en los índices de Sørensen y Jaccard. Ambos taxa de borregos compiten por recursos alimentarios de acuerdo con su disponibilidad en el hábitat estudiado.
Las dos especies de borrego evidencian una preferencia por especies arbustivas y arbóreas, pero su grado de competencia no es lo suficientemente alto para afectar una especie a la otra; el grado de selección y las plantas elegidas para su consumo no están determinadas por las plantas; sin embargo, la competencia se presenta solo cuando ambos borregos disponen de los mismos recursos y estos son limitados o reducidos dentro de su hábitat.

