











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El banco de semillas del suelo es fundamental en la dinámica vegetal, constituye una importante etapa de la regeneración, pues almacena las especies de plantas que potencialmente se establecerán (De Souza et al., 2006). Los bancos de semillas son dinámicos en espacio y tiempo (Fenner y Thompson, 2005): en espacio, los frutos y las semillas se dispersan cuando caen al suelo o mediante distintos vectores como el viento, agua o los animales (Pijl, 1969), y en el tiempo, las semillas pueden permanecer latentes en el suelo, es decir, retrasan la germinación debido a características físicas o fisiológicas en sus estructuras, o simplemente porque no hay las condiciones ambientales adecuadas para germinar (Baskin y Baskin, 2004). Mediante la dispersión y la lluvia de semillas, estas llegan al suelo, mientras que con la germinación, depredación y la muerte por patógenos, son eliminadas del mismo (Fenner y Thompson, 2005).
Los bancos de semillas del suelo pueden contener las especies presentes en la vegetación o que lo estuvieron antes de un disturbio y en áreas cercanas (Bossuyt y Honnay, 2008), por lo que representan un importante recurso para la restauración y resiliencia de una comunidad vegetal dañada (Shiferaw et al., 2018). Tradicionalmente, en las comunidades perturbadas, la degradación se evalúa solo por el estado de la vegetación en pie, y se olvida que el análisis de las semillas almacenadas en el suelo proporciona información crucial para conocer la dinámica vegetal, interpretar el estado de conservación y determinar el potencial de recuperación de las plantas (Ma et al., 2021).
Las investigaciones sobre las variaciones temporales y espaciales del banco de semillas se abordan estudiando sus características básicas como la densidad, diversidad o riqueza. Un aspecto importante que tiene influencia en la distribución espacial de las semillas es su mecanismo de dispersión, ya que mediante este proceso las poblaciones de plantas crecen, intercambian genes y sobreviven a cambios ambientales (Levin y Muller-Landau, 2000). Por ejemplo, las semillas dispersadas por animales son depositadas con mayor frecuencia cerca de árboles con dispersión similar que aquellos con diferente mecanismo de dispersión (Saatkamp et al., 2013). Esto se debe a que las plantas con frutos atractivos para los animales suelen ser visitadas por ellos para alimentarse, protegerse, anidar o perchar (en el caso de aves), y mientras realizan estas actividades, al defecar depositan las semillas bajo las copas de los árboles (DeMars et al., 2010).
Los árboles aislados, aquellos que permanecen en pie después de un disturbio o intencionalmente se dejan en áreas deforestadas, son ampliamente estudiados y utilizados por su influencia en la dinámica de regeneración y restauración ecológica (Guevara et al., 2005). Además, si estas especies atraen animales frugívoros, suelen actuar como depósitos de semillas, aumentan las interacciones con otras plantas y animales, y con ello se mantiene alta biodiversidad (Camargo et al., 2020). Es probable que lo anterior ocurra en las selvas y bosques con mayor frecuencia, porque hay más especies zoocoras que en otros ecosistemas (Gentry, 1982; Willson et al., 1990), sin embargo, en áreas perturbadas de zonas áridas, la dispersión zoócora y la acumulación de semillas bajo árboles aislados también ocurre (Filazzola et al., 2019) y de su estudio depende si se constituye como una herramienta útil para la recuperación de la vegetación y las interacciones que se pierden con el continuo cambio de uso de suelo.
El Matorral Espinoso Tamaulipeco se localiza en una zona semiárida al Noreste de México, es un tipo de vegetación muy fragmentado, compuesto por una matriz de vegetación inducida (para ganadería o agricultura) con pequeños parches remanentes de vegetación nativa, así como árboles aislados de diferentes especies (Molina-Guerra et al., 2013). En el matorral se reconocen 37 especies que tienen dispersión zoócora, lo que representa un mayor porcentaje de taxones con este tipo de dispersión en comparación con otras zonas semiáridas (Jurado et al., 2001a). No obstante, las relaciones entre las plantas y los dispersores, así como la influencia del tipo de dispersión en la regeneración, han sido escasamente estudiadas en este ecosistema.
En el Matorral Espinoso Tamaulipeco se han realizado investigaciones de los bancos de semillas con diversos métodos (Pando-Moreno et al., 2010; Martínez-Adriano et al., 2021; Valdes-Alameda et al., 2021); se conocen los bancos de semillas dentro de fragmentos de vegetación sin perturbación y en los bordes de estos, pero no se tiene información sobre los bancos en sitios destinados a la ganadería, el uso de suelo más común de la región. Por otro lado, debido a la fragmentación del matorral, el papel de los árboles aislados se ha estudiado en el contexto productivo para la ganadería o agroforestería (Sarmiento-Muñoz et al., 2019), mientras que en el ecológico se estudió la remoción de semillas y la producción de frutos para compararla con la producida en la vegetación continua remanente (Jurado et al., 2006; Cuéllar-Rodríguez y Jurado, 2016).
En este trabajo se determinaron las características del banco de semillas, en términos de riqueza y diversidad, bajo individuos aislados de especies arbóreas con diferente síndrome de dispersión (clasificados por las características de sus frutos). Se comparó el banco de semillas del suelo bajo las especies nativas Diospyros texana Scheele y Neltuma laevigata (Humb. & Bonpl. ex Willd.) Britton & Rose (antes Prosopis laevigata (Humb. & Bonpl. ex Willd.) M. C. Johnst.), Yucca filifera Chabaud (de dispersión zoócora), Parkinsonia aculeata L. y Vachellia farnesiana (L.) Wight & Arn. (antes Acacia farnesiana (L.) Willd.) (sin síndrome de dispersión aparente). Además, se exploró la variación temporal del banco de semillas estudiado. Las hipótesis fueron: (1) Los individuos aislados de especies de dispersión zoócora tienen mayor riqueza y densidad de semillas bajo sus copas que las especies con otro tipo de dispersión debido a una mayor deposición de semillas generada por la visita de dispersores que son atraídos a los frutos zoócoros, y (2) La composición de los bancos es variable entre las estaciones debido a que las plantas de la comunidad producen semillas en diferentes estaciones.
Materiales y Métodos
Área de estudio
El área de estudio se ubica dentro de las instalaciones de la Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León, en el municipio Linares, Nuevo León. Está entre las coordenadas 24°48'2.81" N y 99°31'54.51" O al norte, 24°47'50.17" N y 99°32'10.16" O al sur, 24°47'50.80" N y 99°31'39.01" O al este, y 24°47'59.27" N y 99°32'18.12" O al oeste, y tiene una altitud promedio de 379 m. El clima en el área es semicálido subhúmedo con lluvias en verano, descrito como Bs hw(w) según la clasificación de Köppen modificada por García (2004). Las lluvias son irregulares de mayo a junio y entre septiembre a octubre se presentan dos periodos de sequía: uno corto en verano y otro largo en invierno (Foroughbakhch et al., 2013). La temperatura media anual es 23.7 °C y la precipitación media alcanza los 810 mm (Martínez-Adriano et al., 2021). El área tiene 40 ha y consiste en parcelas de pastoreo rotativo de bovinos, con árboles aislados de diferentes especies que corresponden a la vegetación previa al desmonte y que está presente en las zonas aledañas: Matorral Espinoso Tamaulipeco.
La ganadería disminuye significativamente la abundancia, dominancia y diversidad de las comunidades arbóreas y arbustivas de este matorral (Mora et al., 2013) en el cual se han contabilizado alrededor de 160 especies (Foroughbakhch et al., 2013). Aunque no todas se han clasificado por síndrome de dispersión, se registran seis taxones con frutos explosivos (autócoras), 37 con adaptaciones para la dispersión animal (zoócoras), 31 con características de dispersión por el viento (anemócoras) y 43 especies no tienen características claramente asociadas a algún vector de dispersión (no asistidas) (Jurado et al., 2001a).
La fenología de las especies se ha estudiado en un número limitado, pero la tendencia es a que la temporada de dispersión ocurra alrededor de junio y julio (inicio del verano), o de octubre a noviembre (finales de otoño y principios de invierno), algunos taxa presentan frutos en ambas temporadas (García, 1997). En esta región, las semillas de algunas especies pueden germinar por encima de su intervalo de distribución actual, por ello tendrían la capacidad de desplazarse hacia altitudes superiores por efecto del cambio climático (Pérez-Domínguez et al., 2013).
Selección de especies y muestreo
Las especies seleccionadas fueron mezquite (Neltuma laevigata), chapote blanco (Diospyros texana) y yuca (Yucca filifera) que tienen dispersión zoócora, además de huizache (Vachellia farnesiana) y retama (Parkinsonia aculeata), cuyas características de sus frutos las clasifican como no asistidas, es decir, sus frutos carecen de rasgos evidentes que se relacionen con un síndrome de dispersión (Jurado et al., 2001a). Los criterios para seleccionar 10 individuos de cada especie fueron: (1) No tener ningún otro individuo arbóreo adulto o en fructificación en un radio de 10 m alrededor de él, (2) Un promedio de copa de 6 m (±1.0), con excepción de la yuca que se seleccionó con una copa de 10 m (±1.0), y (3) Estar ubicados al menos a 90 m (±10) de distancia del fragmento de matorral continuo más cercano.
Se recolectaron cuatro muestras por árbol (40 por especie) durante los dos años de estudio para evaluar el banco de semillas bajo las copas de cada árbol seleccionado. El periodo de recolección fue en 2021 y 2022, dos veces al año, en marzo y septiembre (a principios de la primavera y el otoño) que corresponden a las temporadas posteriores a la dispersión de la mayoría de las plantas del matorral y antes de la temporada de germinación.
El sitio de recolecta se determinó a una distancia media entre el fuste y el límite de la copa de cada individuo, cada temporada de muestreo se eligió un sitio diferente. El tamaño de la muestra de suelo fue de 30×30 cm y 3 cm de profundidad (alrededor de 1 600±132 g).
Emergencia e identificación de especies
El banco de semillas se contabilizó mediante el método de emergencia de plántulas, que es el más útil para describir las reservas de toda la comunidad vegetal (Piudo y Cavero, 2005). Las muestras de suelo se colocaron en charolas para germinación en el invernadero de la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León, se mantuvieron bajo la luz natural con malla sombra (50 %) y temperatura de 20 a 25 °C. Los riegos se realizaron cada tercer día para mantener la humedad constante. Todos los días se registraron el número y la especie de todas las plántulas que emergieron durante el experimento en un periodo de cuatro meses (cuando no germinaron nuevas plántulas por 15 días), se realizó la identificación utilizando el manual de plantas vasculares de Texas (Correll y Johnston, 1970). Las plántulas identificadas se retiraron de los recipientes y las no reconocidas se trasplantaron a bolsas de vivero para permitir su crecimiento y posterior identificación. Cuando no fue posible asignar el nombre de la especie se identificó hasta el género. La emergencia de una plántula se consideró como una semilla viable. Una vez identificadas las plántulas, se obtuvo información del origen, hábito y el tipo de dispersión de las especies (Cuadro 1). Se contabilizó el número de plántulas emergidas sin conocer el tiempo que las semillas habían pasado en el suelo.
Cuadro 1 Composición de especies del banco de semillas, hábito de crecimiento, origen y síndrome de dispersión asignado.
| Especie | Hábito | Origen | Síndrome de dispersión | |
|---|---|---|---|---|
| Nombre científico | Nombre común | |||
| Acanthaceae | ||||
| Ruellia nudiflora (Engelm. & A. Gray) Urb. | Ruellia | Herbácea | Nativa | Fruto explosivo (Vargas-Mendoza et al., 2015) |
| Amaranthaceae | ||||
| Amaranthus palmeri S. Watson | Herbácea | Nativa | No asistida (Jurado et al., 2001a) | |
| Amaranthus viridis L. | Herbácea | Exótica | Anemócora/hidrócora (SER y RBGK, 2023) | |
| Chenopodium album L. | Quelite | Herbácea | No asistida (Williams, 1963) | |
| Apiaceae | ||||
| Cyclospermum leptophyllum (Pers.) Sprague ex Britton & P. Wilson | Apio silvestre | Herbácea | Nativa | Anemócora (Ogle, 2023) |
| Asparagaceae | ||||
| Yucca filifera Chabaud | Yuca | Arbórea | Nativa | Zoócora (Waitman et al., 2012)* |
| Asteraceae | ||||
| Calyptocarpus vialis Less. | Hierba de caballo | Herbácea | Nativa | No asistida (Valerio y Moreira, 1986) |
| Cirsium texanum Buckley | Cardo | Herbácea | Nativa | Anemócora (SER y RBGK, 2023) |
| Conyza canadensis (L.) Cronquist | Herbácea | Prob. nativa | Anemócora (SER y RBGK, 2023) | |
| Erigeron L. sp. | Herbácea | Anemócora (Pijl, 1969)* | ||
| Helianthus annuus L. | Girasol | Herbácea | Nativa | No asistida |
| Parthenium confertum A. Gray | Herbácea | Nativa | Anemócora (Mao et al., 2019) | |
| Sanvitalia ocymoides DC. | Herbácea | Nativa | Anemócora (Jurado et al., 2001a) | |
| Sonchus oleraceus L. | Lechuguilla común | Herbácea | Exótica | Anemócora (SER y RBGK, 2023) |
| Taraxacum officinale F. H. Wigg. | Diente de león | Herbácea | Anemócora (SER y RBGK, 2023) | |
| Boraginaceae | ||||
| Lithospermum matamorense DC. | Herbácea | Nativa | No asistida (Pijl, 1969)* | |
| Brassicaceae | ||||
| Lepidium virginicum L. | Lenteja de campo | Herbácea | Nativa | Anemócora (Díaz y Ríos, 2017) |
| Cactaceae | ||||
| Opuntia Mill. sp. | Nopal | Arbórea | Nativa | Zoócora (SER y RBGK, 2023) |
| Campanulaceae | ||||
| Lobelia L. sp. | Herbácea | Anemócora (SER y RBGK, 2023) | ||
| Cannabaceae | ||||
| Celtis laevigata Willd. | Palo blanco | Arbórea | Nativa | Zoócora (Jurado et al., 2001a) |
| Celtis pallida Torr. | Granjeno | Arbórea | Nativa | Zoócora (Jurado et al., 2001a) |
| Convolvulaceae | ||||
| Convolvulus arvensis L. | Herbácea | Exótica | No asistida (SER y RBGK, 2023) | |
| Ipomoea L. sp. | Herbácea | No asistida (SER y RBGK, 2023) | ||
| Cordiaceae | ||||
| Cordia boissieri A. DC. | Anacahuita | Arbórea | Nativa | Zoócora (Jurado et al., 2001a) |
| Ebenaceae | ||||
| Diospyros texana Scheele | Chapote blanco | Arbórea | Nativa | Zoócora (Jurado et al., 2001a) |
| Euphorbiaceae | ||||
| Argythamnia humilis (Engelm. & A. Gray) Müll. Arg. | Herbácea | Nativa | No asistida (SER y RBGK, 2023) | |
| Croton ciliatoglandulifer Ortega | Croton | Arbustiva | Nativa | Zoócora (Jurado et al., 20017a) |
| Croton cortesianus Kunth | Croton | Herbácea | Nativa | Zoócora (Jurado et al., 2001a) |
| Croton fruticulosus Engelm. ex Torr. | Croton | Arbustiva | Nativa | Zoócora (Jurado et al., 2001a) |
| Croton incanus Kunth | Croton | Arbustiva | Nativa | Zoócora (Jurado et al., 2001a) |
| Euphorbia prostrata Aiton | Hierba de la golondrina | Herbácea | Nativa | No asistida (Baiges et al., 1991) |
| Tragia ramosa Torr. | Ortiguilla | Herbácea | Nativa | No asistida (Everitt et al, 1999; SER y RBGK, 2023) |
| Fabaceae | ||||
| Desmanthus virgatus (L.) Willd. | Herbácea | Nativa | No asistida (Jurado et al., 2001a) | |
| Galactia texana (Scheele) A. Gray | Herbácea | Nativa | Fruto explosivo (Jurado et al., 2001a) | |
| Mimosa monancistra Benth. | Uña de gato | Arbustiva | Nativa | No asistida (Jurado et al., 2001a) |
| Neltuma laevigata (Humb. & Bonpl. ex Willd.) Britton & Rose | Mezquite | Arbórea | Nativa | Zoócora (Jurado et al., 2001a) |
| Parkinsonia aculeata L. | Retama | Arbórea | Nativa | Hidrócora (SER y RBGK, 2023) |
| Vachellia farnesiana (L.) Wight & Arn. | Huizache | Arbórea | Nativa | No asistida (Jurado et al., 2001a) |
| Lamiaceae | ||||
| Teucrium cubense Jacq. | Herbácea | Nativa | No asistida (Jurado et al., 2001a) | |
| Malvaceae | ||||
| Malvastrum coromandelianum (L.) Garcke | Malva | Herbácea | Prob. nativa | No asistida (Jurado et al., 2001a) |
| Sida rhombifolia L. | Herbácea | Zoócora (Mori y Brown, 1998) | ||
| Namaceae | ||||
| Nama jamaicensis L. | Herbácea | Nativa | No asistida (Pijl, 1969)* | |
| Onagraceae | ||||
| Oenothera speciosa Nutt. | Hierba del golpe | Herbácea | Nativa | Anemócora (SER y RBGK, 2023) |
| Oxalidaceae | ||||
| Oxalis L. sp. | Herbácea | Nativa | Fruto explosivo (Unison y ASU, 2023) | |
| Plantaginaceae | ||||
| Plantago lanceolata L. | Plantago | Herbácea | Exótica | Anemócora (SER y RBGK, 2023) |
| Portulacaceae | ||||
| Portulaca L. sp. | Verdolaga | Herbácea | Exótica | No asistida (SER y RBGK, 2023) |
| Rhamnaceae | ||||
| Condalia hookeri M. C. Johnst. | Brasil | Arbórea | Nativa | Zoócora (Jurado et al., 2001a) |
| Rubiaceae | ||||
| Diodia teres Walter | Herbácea | Nativa | No asistida (SER y RBGK, 2023) | |
| Randia rhagocarpa Standl. | Cruceto | Arbórea | Nativa | Zoócora (Jurado et al., 2001a) |
| Rutaceae | ||||
| Zanthoxylum fagara (L.) Sarg. | Colima | Arbórea | Nativa | Zoócora (Jurado et al., 2001a) |
| Sapotaceae | ||||
| Sideroxylon celastrinum (Kunth) T. D. Penn. | Coma | Arbórea | Nativa | Zoócora (Jurado et al., 2001 a ) |
| Solanaceae | ||||
| Capsicum annuum L. var. glabriusculum (Dunal) Heiser & Pickersgill | Chile piquín | Arbustiva | Nativa | Zoócora (Jurado et al., 2001a) |
| Physalis viscosa L. | Tomatillo | Arbustiva | Nativa | Zoócora (Jurado et al., 2001a) |
| Urticaceae | ||||
| Urtica chamaedryoides Pursh | Ortiguilla | Herbácea | Nativa | No asistida (Pijl, 1969)* |
| Verbenaceae | ||||
| Glandularia bipinnatifida (Nutt.) Nutt. | Alfombrilla | Herbácea | Nativa | Anemócora (Pijl, 1969)* |
| Lantana L. sp. | Lantana | Arbustiva | Nativa | Zoócora (SER y RBGK, 2023) |
| Verbena canescens Kunth | Verbena | Herbácea | Nativa | Anemócora (Jurado et al., 2001a) |
| Violaceae | ||||
| Hybanthus verticillatus (Ortega) Baill. | Arbustiva | No asistida (Pijl, 1969)* | ||
* Observación personal basada en la cita; Prob. Nativa = Probablemente nativa.
Análisis estadístico
Con los datos obtenidos se calcularon la riqueza y densidad por metro cuadrado de plántulas (plántulas m2), así como la riqueza y densidad de especies herbáceas y arbóreas. La densidad está dada en metros cuadrados para facilitar su contraste con otros trabajos. Cada parámetro fue comparado entre los dos años de estudio, las estaciones en que se realizó la recolecta (primavera y otoño) y entre las especies de árboles aislados.
Dado que los datos no cumplieron con el supuesto de normalidad (P<0.01), se hizo una comparación de medianas de la riqueza y densidad con la prueba no paramétrica Kruskall-Wallis y el método de corrección de Bonferroni (para comparación entre especies), también se utilizó la prueba U de Mann-Whitney (para comparación entre estaciones y años) (Zar, 1999). Los análisis se efectuaron con en el programa SPSS statistics versión 25.0 (IBM, 2022).
Resultados
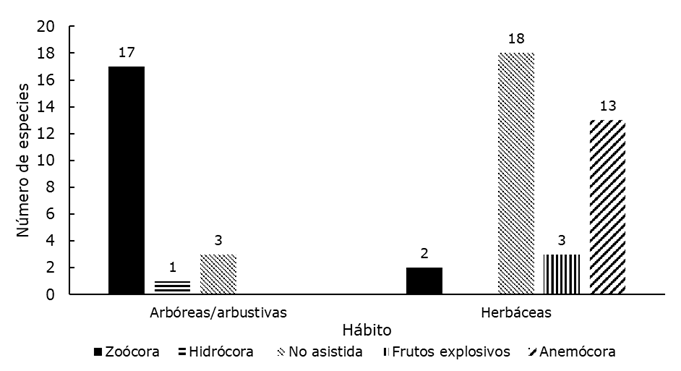
Un total 57 especies pertenecientes a 29 familias se identificaron en el banco de semillas (Cuadro 1): 21 fueron arbóreas o arbustivas y 36 herbáceas (Figura 1). Del total de especies, 21 se clasificaron con dispersión no asistida, 19 como zoócoras, 14 anemócoras, tres con frutos explosivos o autócoras y una hidrócora o dispersada mediante el agua (Jurado et al., 2001a). La mayoría de los taxones (45) se consideraron nativas o probablemente nativas, y cinco como exóticas. Los taxa más abundantes durante los dos años de estudio fueron Malvastrum coromandelianum (L.) Garcke (754 plántulas en total) y Nama jamaicensis L. (616 plántulas en total), ambas herbáceas. Las especies arbóreas más abundantes fueron Neltuma laevigata (54 plántulas en total) y Yucca filifera (49 plántulas en total), aunque la mayoría bajo la planta Madre, y las más abundantes bajo una especie diferente fueron Opuntia Mill. sp. (44 plántulas) y Zanthoxylum fagara (L.) Sarg. (39 plántulas). Las familias mejor representadas fueron Asteraceae (9) y Euphorbiaceae (7), seguidas de Fabaceae (6).
Con base en el análisis del banco de semillas del suelo bajo cada especie de árbol aislado, se determinó que la riqueza y densidad de plántulas fueron similares entre los dos años de estudio (Riqueza N. laevigata: U=186, Z=-0.38, P=0.71; V. farnesiana: U=194, Z=-0.79, P=0.88; D. texana: U=134, Z=-1.79, P=0.76; Y. filifera: U=135, Z=-1.78, P=0.81; P. aculeata: U=192, Z=0.22, P=0.84; Densidad N. laevigata: U=197, Z=-0.08, P=0.97; V. farnesiana: U=180, Z=-0.58, P=0.60; D. texana: U=147, Z=-1.43, P=0.15; Y. filifera: U=184, Z=-0.42, P=0.67, P. aculeata: U=147, Z=-1.43, P=0.15).
La comparación entre estaciones de recolecta resultó en una mayor riqueza y densidad de plántulas en el banco correspondiente al otoño bajo todas las especies (Riqueza N. laevigata: U=49, Z=-4.12, P<0.01; V. farnesiana: U=86, Z=-3.10, P=0.002; D. texana: U=80, Z=-3.27, P=0.001; Y. filifera: U=60, Z=-3.82, P<0.01; P. aculeata: U=64, Z=-3.77, P<0.01; Densidad N. laevigata: U=40, Z=-4.13, P<0.01; V. farnesiana: U=80, Z=-3.24, P=0.001; D. texana: U=132, Z=-1.82, P=0.06; Y. filifera: U=74, Z=-3.42, P<0.01; P. aculeata: U=63, Z=-3.71, P<0.01).
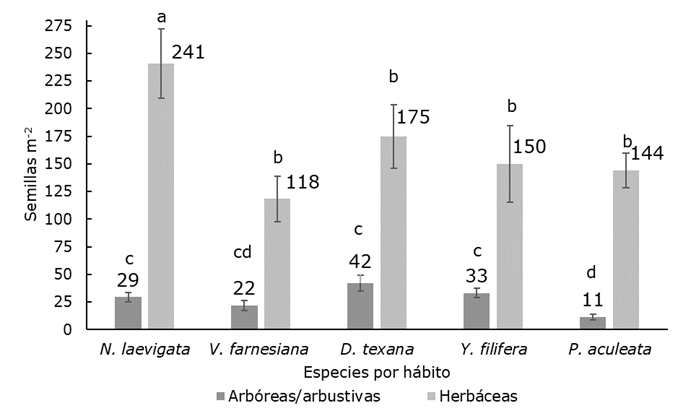
Los taxones de dispersión zoócora (N. laevigata y D. texana) tuvieron bajo sus copas más riqueza en promedio que el resto de las especies (H=21.14; g. l.=4; P<0.001). Además, la densidad de plántulas fue más grande bajo N. laevigata que para V. farnesiana y P. aculeata, especies con semillas no asistidas, incluso fue mayor bajo N. laevigata con respecto a Y. filifera que también es zoócora (H=23.44; g. l.=4; P<0.001) (Figura 2).

Los datos de densidad están redondeados al número entero más cercano. Las barras de error representan la desviación estándar.
Figura 2 Densidad promedio de plántulas bajo cada especie.
En general, se obtuvo más riqueza y densidad de herbáceas que de arbóreas bajo todas las especies (Riqueza: U=3 765, Z=-3.11, P=0.002; Densidad: U=4 075, Z=-2.31, P=0.02). Emergieron más plántulas de arbóreas en promedio bajo D. texana y Y. filifera (dos especies, H=29.93, g. l.=4, P<0.001) y de herbáceas bajo N. laevigata (cinco especies, H=13.96, g. l.=4, P=0.007). La densidad más alta de plántulas de especies arbóreas se contabilizó bajo N. laevigata, D. texana y Y. filifera, mientras que la densidad de herbáceas fue mayor bajo N. laevigata (H=22.47, g. l.=4, P<0.001; H=16.97, g. l.=4, P=0.002, respectivamente) (Figura 3).
Discusión
En este estudio se obtuvieron resultados de una mayor riqueza de especies en el banco de semillas que en cualquier otro realizado en el Matorral Espinoso Tamaulipeco, se identificaron 27 especies más que lo registrado por Martínez-Adriano et al. (2021), quienes citan 30 especies en los bancos de semillas presentes en fragmentos remanentes del Matorral Espinoso Tamaulipeco. Asimismo, la riqueza fue superior a la estimada en un área de matorral denso (33 especies) (Pando-Moreno et al., 2010). Por otro lado, las densidades promedio de plántulas m2 bajo N. laevigata y V. farnesiana se ubican entre las registradas en el área densa (1 900 semillas m2) y en un área erosionada (62 semillas m2), ambas con Matorral Espinoso Tamaulipeco (Pando-Moreno et al., 2010).
No hubo variabilidad anual en la densidad y riqueza del banco de semillas, pero si una variación estacional, ya que hubo más especies y densidad en el banco de semillas recolectado en otoño. En otras regiones semiáridas se registran variaciones en diferentes años en la riqueza de especies, abundancia y densidad de semillas en el suelo debido a variaciones por las condiciones climáticas extremas (Quevedo-Robledo et al., 2010; Dreber y Esler, 2011; da Silva et al., 2013). La producción de frutos puede cambiar en función de factores ambientales y recursos que son variables en el tiempo, en consecuencia, se observan cambios temporales en los bancos de semillas (Quevedo-Robledo et al., 2010), aunque algunas plantas son muy constantes y mantienen sus patrones fenológicos o responden a la variabilidad de recursos con un desplazamiento de algunos días en sus fases fenológicas (Gordo y Sanz, 2009). Es posible que una ligera variación en la producción de los frutos no sea suficiente para que se refleje en la composición del banco de semillas en estudios a corto plazo.
Por otro lado, en las plantas zoócoras, la abundancia de los consumidores de frutos puede disminuir o aumentar la densidad de semillas depositadas (García y Martínez, 2012), sin embargo, la información sobre estas interacciones en la región es escasa o nula. Además, las especies en una comunidad suelen tener diferentes estrategias de germinación, y con el método utilizado en esta investigación, dichas estrategias influyen directamente en los resultados, por lo que la variación estacional registrada podría estar sujeta al cumplimiento o no de las condiciones necesarias de germinación. Para las plantas del matorral, Jurado et al. (2001b) indicaron que algunas de las especies estudiadas germinaron más en otoño, lo que concuerda con los resultados que aquí se documentan. Es probable que la investigación del banco de semillas realizada con otros métodos como el de tamizado, ayuden a confirmar si hay diferencias en el número de semillas o las variaciones estacionales detectadas se deban a las adaptaciones de germinación de la comunidad. El análisis del banco de semillas debe contemplar esta variación estacional, ya que el muestreo en una sola temporada estaría aportando información incompleta para la adecuada evaluación de las áreas perturbadas o la determinación de la riqueza vegetal potencial.
Los resultados muestran que las plantas zoócoras tuvieron más especies y densidad de semillas bajo sus copas, lo que concuerda con la hipótesis planteada. Resultados similares se han descrito para otras zonas áridas, semiáridas y bosques secos (Warnock et al., 2007; Hadinezhad et al., 2021), además de ecosistemas tropicales (Camargo et al., 2020), mediterráneos (González‐Varo et al., 2017) y templados (Rodríguez‐Pérez et al., 2014).
La mayoría de las semillas de las especies arbóreas identificadas tienen características de dispersión zoócora y están bajo una especie arbórea zoócora (D. texana), por lo que es probable que la fauna dispersora, entre otros factores, esté moldeando la comunidad arbórea como sucede en otros ecosistemas (Warnock et al., 2007; Hadinezhad et al., 2021). Adicionalmente, en el matorral la remoción de semillas es más lenta bajo los árboles aislados que en el matorral denso (Jurado et al., 2006), lo que también puede influir en la cantidad de especies identificadas. Jurado et al. (2006) describieron que la remoción fue más lenta, especialmente, en árboles aislados de N. laevigata, la cual es una de las especies con mayor riqueza y densidad de semillas en el Matorral Espinoso Tamaulipeco estudiado. Es posible que el mismo proceso de remoción determine la composición de semillas en individuos aislados de chapote, que fue similar a la del mezquite. Lo anterior parece indicar que en esta región semiárida, la elección de las especies de árboles para dejar en pie o reforestar influye en la capacidad de las semillas de llegar a sitios perturbados y permanecer en el banco de semillas.
En el banco de semillas bajo N. laevigata se obtuvo la mayor cantidad de especies herbáceas, con una densidad alta y dispersión anemócora o no asistida. Asimismo, las herbáceas superaron a las arbóreas bajo todas las especies. Lo anterior se relaciona con el hecho de que las herbáceas suelen producir muchas semillas pequeñas, mientras que las arbóreas producen menos semillas, pero más grandes (Moles et al., 2004; Moles et al., 2005), además son más seleccionadas por los depredadores (Dylewski et al., 2020), lo que pudo influir en la menor densidad registrada. Otro factor es que en general, las herbáceas tienen mayor porcentaje de especies con latencia en comparación con las leñosas en los ecosistemas con condiciones extremas como aquellos con heladas o sequías, como es el Matorral Espinoso Tamaulipeco. La proporción entre las especies con latencia o sin ella no difiere de manera significativa (Jurado y Flores, 2005).
La información generada es importante porque representa la riqueza potencial que se obtendría en una trayectoria de regeneración natural. Con la evaluación de los bancos de semillas y de la vegetación en pie, es posible interpretar si las zonas que parecen degradadas recuperarán la estructura y diversidad de vegetación anterior al disturbio o tendrán un estado alternativo (Ma et al., 2021). En el presente estudio se determinó una alta densidad y riqueza de especies nativas, además de que se infiere que ocurre un ingreso de semillas de áreas cercanas, lo que sugiere que el área estudiada está en un estado apto para recuperar su vegetación a través de la regeneración natural una vez que dejen de presentarse los disturbios.
Conclusiones
Los árboles aislados de las especies nativas Diospyros texana, Neltuma laevigata, Yucca filifera, Parkinsonia aculeata y Vachellia farnesiana del Matorral Espinoso Tamaulipeco, son reservorios de una alta riqueza y diversidad de semillas. Los bancos de semillas conformados bajo especies con dispersión zoócora mantienen mayor riqueza y densidad de semillas que los integrados bajo taxones con otros síndromes de dispersión. El número de especies y la densidad de semillas germinables tienen una variación estacional, ya que los parámetros evaluados son mayores en otoño que en primavera.

