











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
Introducción
La riqueza de especies se ha relacionado con la productividad y producción de la vegetación a través de diferentes enfoques y marcos teóricos para explicar los procesos subyacentes y sus factores (LaRue et al., 2019; Sheil y Bongers, 2020; Wang et al., 2020), aunque con resultados insuficientes para explicar la variabilidad observada (Wang et al., 2019).
Con relación al funcionamiento de los ecosistemas, se ha citado un incremento de la productividad o producción de la vegetación con la diversidad de especies de plantas (Liang et al., 2016). En este sentido, Tilman (1982, 1999) y Tilman et al. (1997), mediante el uso de modelos de competencia de recursos, plantearon que la producción (biomasa) crece con el número de especies que compiten por un solo recurso, por lo que la variación en la producción de biomasa es alta y su varianza se reduce al aumentar el número de taxones. En el caso de competencia por dos recursos (por ejemplo: agua y nutrientes) se observa el mismo patrón; por lo que, si el número de especies es mayor, los recursos se usan totalmente y se presenta un patrón de saturación.
Tilman et al. (1996) mostraron en un experimento sobre la relación productividad/producción-riqueza de especies en un pastizal que la cobertura proyectiva foliar se incrementa con el número de especies, lo que define una relación funcional específica, más allá de la relación biomasa-riqueza de especies.
Vilá et al. (2005) analizaron evidencia experimental en sistemas forestales y plantearon que hay una relación entre el número de taxones y la productividad/producción de los bosques antes del cierre de la cobertura del dosel (etapas iniciales de la sucesión o en bosques con limitaciones de estrés), y después esta se modifica al declinar (si sucede) el bosque. Al inicio de la sucesión, la cobertura de arbustos aumentó con el incremento de la riqueza de árboles, lo cual implicó que no interfirieran con la producción arbórea.
Wang et al. (2020) analizaron la relación productividad/producción-riqueza de especies y determinaron patrones cóncavos, asintóticos, positivos, negativos e irregulares, con un efecto positivo en la productividad. Ónodi et al. (2021) registraron diferentes relaciones antes y después de un incendio, por lo que la influencia de los disturbios en la sucesión vegetal condiciona la estabilidad de los ecosistemas.
La cobertura aérea del follaje de la vegetación tiene un efecto positivo en la diversidad de especies; la intensidad de las perturbaciones ejerce uno negativo en la cobertura aérea y un impacto variable en la riqueza de especies (Sanaei et al., 2018). La cobertura aérea del follaje, medida como disponibilidad de iluminación por radiación, está fuerte y positivamente correlacionada con la riqueza de especies (Dormann et al., 2020).
El objetivo de este trabajo fue analizar la relación de la productividad/producción vegetal con relación a la riqueza de especies para el establecimiento de condiciones necesarias para el incremento de la biomasa aérea y la cobertura proyectiva foliar, además de la relación entre la cobertura proyectiva foliar y el número de especies para los diferentes tipos de vegetación existentes en el Estado de México, producto de un muestreo extensivo de sus diferentes comunidades vegetales.
Materiales y Métodos
Campaña de muestreo en el Estado de México
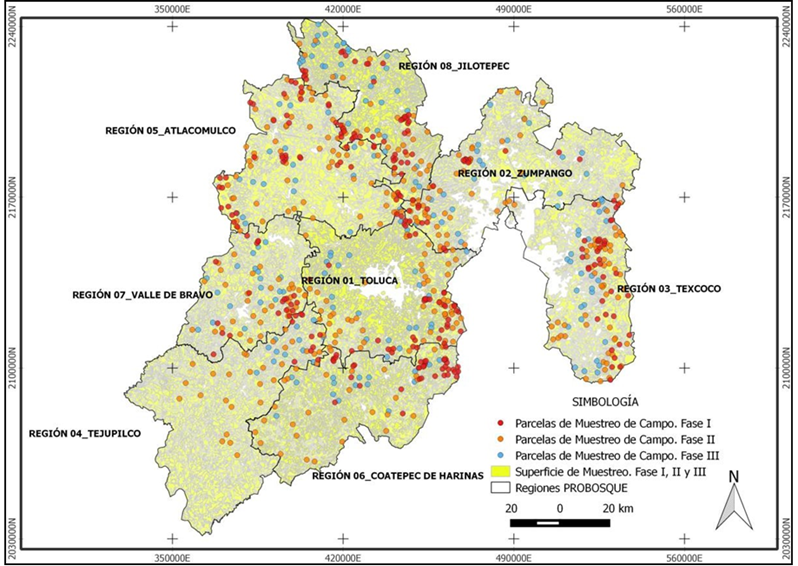
En 2014 se implementó una campaña de campo para realizar el inventario de los almacenes de carbono y de riqueza de plantas vasculares en los diferentes tipos de vegetación del Estado de México. Para optimizar los recursos, se siguió una estrategia de muestreo en cronosecuencias, en la que primero se muestreó la vegetación primaria (no perturbada), después la vegetación perturbada (vegetación secundaria, en sucesión), y por último la vegetación asociada a cambios permanentes del uso del suelo, fundamentalmente, para actividades antropogénicas (Figura 1).

Fuente: Elaboración propia.
Los círculos en color rojo y amarillo representan los sitios de muestreo en bosque de oyamel (Abies spp.) en diferentes estados de sucesión. BA = Bosque de oyamel primario; BA/VSA = Bosque de oyamel con vegetación secundaria arbórea; BA/VSa = Bosque de oyamel con vegetación secundaria arbustiva; BA/VSh = Bosque de oyamel con vegetación secundaria herbácea.
Figura 1 Fases del muestreo con base en la estratificación del uso del suelo y vegetación homologada para el Estado de México.
Los tipos de vegetación y uso del suelo se clasificaron de acuerdo al sistema del Instituto Nacional de Estadística y Geografía (Inegi), incluidas en la FAO FRA 2010 (Paz-Pellat et al., 2019), en el que la vegetación secundaria (VS) tiene tres estados de sucesión dependientes de la dominancia del estrato: (A) arbóreo, (a) arbustivo o (h) herbáceo. Las fases de la sucesión vegetal permiten cuantificar el nivel de perturbación del ecosistema como una medida de su estabilidad.
Distribución de los muestreos
Se establecieron aleatoriamente sitios de muestreo de 1 000 m2. En el Cuadro 1 se presenta el número de sitios medidos en cada fase de la campaña, con un total de 682. El tamaño de la muestra se estimó con base en el protocolo para calcular el número de puntos de muestro usado en los proyectos de Mecanismos de Desarrollo Limpio para Forestación/Reforestación (A/R CMD project activity, por sus siglas en inglés) de la Convención Marco de Naciones Unidas para el Cambio Climático (UNFCCC, por sus siglas en inglés), mediante la herramienta metodológica A/R CDM Tool (2014), desarrollada por Winrock International (Pearson et al., 2005). Los sitios se distribuyeron en función de las regiones en que la Protectora de Bosques del Estado de México (Probosque) divide a la entidad. El uso de la fase de muestreo permite analizar la vegetación por dominancia del estado de sucesión secundaria.
Cuadro 1 Número de parcelas muestreadas en cada región y fase.
| Región Probosque | Fase de muestreo | ||
|---|---|---|---|
| I | II | III | |
| 01 Toluca | 40 | 45 | 16 |
| 02 Zumpango | 22 | 28 | 12 |
| 03 Texcoco | 56 | 65 | 20 |
| 04 Tejupilco | 9 | 44 | 10 |
| 05 Atlacomulco | 42 | 50 | 23 |
| 06 Coatepec Harinas | 23 | 30 | 11 |
| 07 Valle de Bravo | 26 | 27 | 13 |
| 08 Jilotepec | 19 | 35 | 16 |
| Total | 237 | 324 | 121 |
En la Figura 2 se muestra la localización espacial de los sitios medidos por región y por fase de muestreo. La variabilidad espacial de los muestreos refleja la de los tipos de vegetación registrados en cada región. Los tipos particulares de vegetación no se exhiben en la Figura 2, dado que los análisis están planteados en términos generales.
Registro de especies
Se consideraron todas las especies de plantas vasculares presentes en cada sitio de muestreo. Se realizó un registro fotográfico de los taxa y solo se colectaron mediante los métodos estándar aquellos que estuvieron en floración o fructificación (Lot y Chiang, 1986). La identificación taxonómica se realizó en gabinete, y se llevó a cabo con ayuda de literatura especializada (Lot y Chiang, 1986).
Se analizó la composición florística (especies, géneros y familias) de los sitios muestreados. Para integrar el catálogo de especies se usó como referencia el sistema de clasificación taxonómica de angiospermas Angiosperm Phylogeny Group IV (2016). En el caso de la nomenclatura de gimnospermas y pteridofitas, se consultó el proyecto de flora mundial Tropicos TM del Herbario del Jardín Botánico de Missouri (Missouri Botanical Garden, 2018), el cual contiene las actualizaciones más recientes de la información taxonómica referente a este tipo de plantas. Los registros de los taxones se efectuaron con la información de campo y se integraron en bases de datos para su análisis.
Cobertura proyectiva foliar
La cobertura proyectiva foliar (CPF) se evaluó con el método desarrollado por Salas-Aguilar et al. (2017) para la estimación de la fracción de la cubierta forestal con el uso de fotografía digital: las imágenes se tomaron con una cámara Canon ® Eos Rebeld T5 configurada con la máxima resolución espacial (17 915 904 pixeles). El lente de la cámara se ajustó a un ISO de 200, la apertura y la exposición se utilizaron en modo automático. Para la captura de la fotografía, el lente se niveló hacia arriba a 1.5 m de altura con respecto al suelo, en los radios de 17.84, 11.28 y 5.65 m, respectivamente, y en el sentido de las manecillas del reloj: de norte a sur, de noreste a suroeste, de este a oeste y de sureste a noroeste (Figura 3).

Equilibrio de la vegetación
Specht (1972) planteó un modelo de equilibrio de la vegetación con la atmósfera y el agua en el perfil del suelo:
Donde:
aprovechable por la vegetación
El valor del agua almacenada en el suelo (Smax) es el agua entre el valor de capacidad de campo y el punto de marchitamiento permanente. La precipitación (P) en exceso de Smax se pierde como drenaje subterráneo (D). El balance hídrico (Ecuación 1) se realiza a escala de parcela; sus valores corresponden a datos mensuales y sus unidades son cm.
Specht (1972) planteó que la vegetación clímax perenne siempre verde aprovecha la totalidad del agua disponible a través del balance del agua en el suelo y la demanda atmosférica, modulada por el coeficiente evaporativo k, de tal forma que
Donde:
El coeficiente evaporativo k integra los efectos de la geometría aérea de la vegetación (foliar en su mayoría) y la resistencia (estomática en su mayoría y la capa límite de la hoja) al movimiento del agua en las plantas (Specht, 1972). Un valor de k grande implica una comunidad de plantas densamente estructurada (baja resistencia).
El coeficiente k se aproxima por un ajuste evolutivo dentro de la comunidad de plantas a un valor máximo que se aproxime, pero nunca es igual a cero. El supuesto es que la comunidad clímax (en equilibrio atmósfera-vegetación-suelo) utiliza completamente la humedad del suelo disponible para el crecimiento de las plantas (acumulación de biomasa), de tal forma que las raíces deben explorar el suelo para usar toda el agua disponible en cualquier parte del perfil edáfico.
En los trabajos de Specht, la CPF de la comunidad de plantas se divide en un estrato superior o ES (cobertura proyectiva del follaje=CPF ES ) e inferior o EI (cobertura proyectiva del follaje=CPF EI ). Para comunidades dominadas por especies arbóreas, la distinción de estratos es una altura de dos metros (Specht y Morgan, 1981; Specht y Specht, 1993; Specht y Tyler, 2010). En el caso de matorrales, pastizales y cultivos, para la separación entre el estrato superior e inferior se utilizan alturas que reflejen la partición de recursos en la comunidad (Specht y Morgan, 1981).
El rasgo estructural más importante de una comunidad de plantas perennes siempre verdes (y caducifolias) es la distribución y orientación del área foliar del tope al fondo de la comunidad, en relación al componente directo y difuso de la radiación. Estos atributos estructurales, junto con la resistencia al flujo del agua determinan el valor de k (Specht, 1972). A partir de esas consideraciones, Specht (1972; 2000) planteó una relación empírica entre k (mm-1) y CPF:
Donde:
CPF ES = Cobertura proyectiva del follaje del estrato superior
CPF EI = Cobertura proyectiva del follaje del estrato inferior
CPF T = Cobertura proyectiva del follaje total
n = Número de datos
R 2 = Coeficiente de determinación
Specht et al. (1991) y Specht y Specht (1993) desarrollaron relaciones entre el coeficiente evaporativo k (cm-1) y la diversidad de especies en comunidades clímax:
Donde:
Ln( ) = Función logaritmo
S T = Riqueza total de especies
S ES = Riqueza de especies del estrato superior
n = Número de datos
R 2 = Coeficiente de determinación
La relación entre la riqueza de especies del estrato inferior (S EI ) es lineal e inversamente proporcional a CPF ES (Specht y Morgan, 1981; Specht et al., 2006). Este patrón se observa en la sucesión pírica de una comunidad vegetal (Specht y Jones, 1971) (Figura 4), lo que muestra que cuando el bosque tiende a su madurez, la diversidad de especies del EI disminuye y la del ES aumenta.
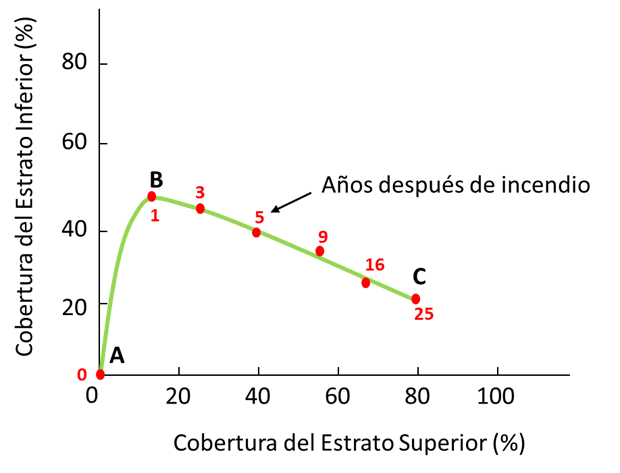
Fuente: Adaptado de Specht y Jones (1971).
Figura 4 Esquematización de la evolución de las coberturas proyectivas foliares en una sucesión pírica.
En el caso del número de especies y la composición de la comunidad de plantas, la trayectoria de sucesión muestra un incremento en la riqueza de especies en el proceso hacia una comunidad clímax. Esta es solo una de las posibles trayectorias, dado que la riqueza y composición de especies tiene un número prácticamente infinito de combinaciones para lograr la condición de equilibrio (clímax). En términos generales, el modelo de Specht plantea que el agua en el perfil del suelo, acoplado a la interacción atmosfera-vegetación es la principal limitante para el desarrollo de las comunidades vegetales.
La relación de la cobertura del follaje entre el estrato superior e inferior, también se ha observado en comunidades vegetales maduras en Estados Unidos de América (Tausch et al., 1981; Tausch y Tueller, 1990).
A partir de la relación lineal entre k y CPF y la relación exponencial entre el número de especies y el coeficiente evaporativo k, se espera una relación exponencial (no lineal) entre la CPF y el número de taxa, que se incrementa con la perturbación en la vegetación, y se refleja en el estado de la sucesión vegetal. Lo anterior aplica, particularmente, para el caso de las comunidades climax.
Procesamiento de datos
Para analizar la relación entre la CPF con la S de plantas vasculares (variables de análisis) se siguió el procedimiento que se describe a continuación:
a) Primero se realizaron los análisis de las relaciones cobertura proyectiva foliar y riqueza de especies para cada región individual (regionalización de Probosque).
b) Por región, para cada tipo de vegetación general y uso del suelo, se identificó el sitio con el mayor número de especies y se le asoció la correspondiente CPF medida en campo.
c) Se seleccionaron todos los sitios de las regiones de Probosque, los cuales se agruparon por tipos de vegetación: bosques/selvas (con sus estados de sucesión), matorrales, pastizales y cultivos.
Resultados
En la Figura 5 se muestran los resultados obtenidos para el caso de bosques/selvas y sus estados de sucesión. La relación entre S y la CPF del estrato superior se ajustó a un modelo lineal:
Donde:
S = Riqueza de especies
A = Constante empírica
B = Constante empírica
CPF ES = Cobertura proyectiva foliar del estrato superior
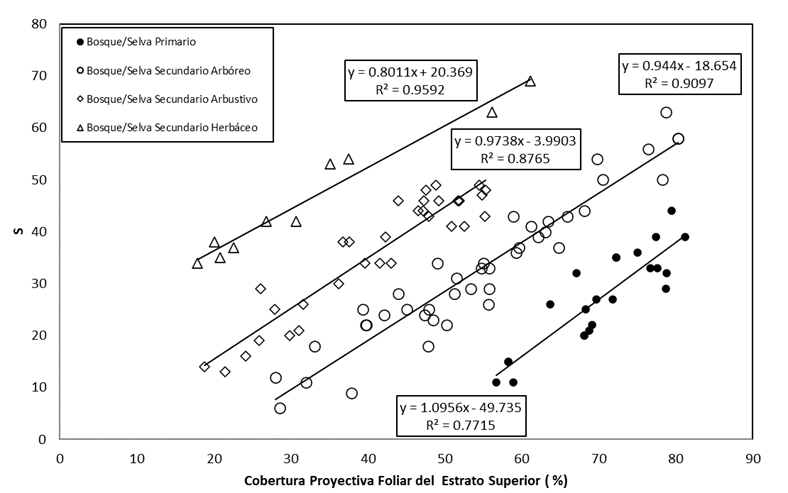
El número de parcelas de muestreo representadas corresponde a 108, aunque no se muestran ocho valores extremos en la parte superior y cuatro valores extremos en la parte inferior, ya que estuvieron fuera de los límites analizados y tenían patrones con pocos datos experimentales.
Dado que se esperaba una relación no lineal entre CPF ES -k, relaciones (3) y (4), en la Figura 6 se observan las relaciones que se establecieron mediante modelos potenciales.
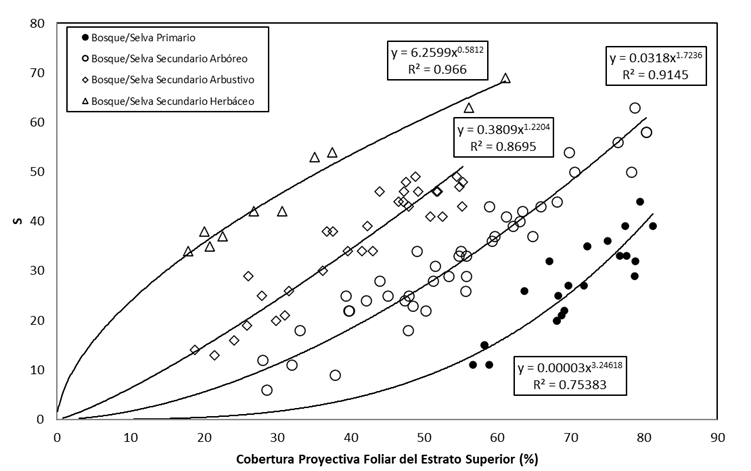
Al realizar un análisis con el modelo potencial S=A(CPF ES ) B , se observó que los parametros ln(A) y B están relacionados entre sí, ya que se interceptan en un punto común (0,1). En la Figura 7 se presenta la relación entre los parámetros de los modelos potenciales ajustados en la Figura 6; la línea recta pasa por el punto (0,1), dado que ln(A=1)=0 en el caso de B=1. Así, la relación entre los parámetros de los modelos potenciales requiere solo de conocer un dato para establecer las relaciones de los diferentes estados de la sucesión de bosques/selvas.
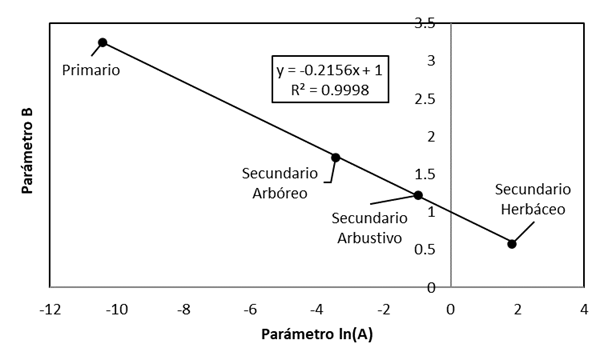
Figura 7 Relacion entre los parámetos de los modelos potenciales para bosques/selvas y sus estados de sucesión.
Para la vegetación diferente a bosque/selva, el número de parcelas muestreadas fue limitado. En la Figura 8 se muestra la relación entre CPF EI y S en el caso de matorrales.

Es las Figuras 9 y 10 se exhiben las relaciones entre la CPF EI y S para pastizales y cultivos agrícolas, respectivamente.


En la Figura 11 se observa la relación entre los parámetros de los modelos potenciales ajustados para la relación de S con la CPF EI para los matorrales, pastizales y cultivos agrícolas que, de manera similar a los bosques/selvas, se ajustaron a una línea recta que pasa por el punto (0,1).

Discusión
La relación entre la riqueza de especies y la cobertura proyectiva foliar para comunidades clímax ha sido determinada por Specht y Morgan (1981) usando gradientes sin disturbios. En el presente estudio, esta relación tuvo patrones congruentes con los planteamientos empíricos de Specht y Specht (1993), particularmente, en función de los estados de la sucesión de bosques/selvas. Cabe señalar que los estudios sobre el particular, prácticamente, se restringe a ecosistemas de clima mediterráneo; hay algunos trabajos en la literatura referentes a bosques templados (Tausch et al., 1981; Tausch y Tueller, 1990) en secuencias de sucesión vegetal. De acuerdo a estas revisiones, la aplicación de la relación CPF-S a otro tipo de ambientes, como los bosques templados y subtropicales del Estado de México, extiende los desarrollos definidos previamente y permite establecer patrones generales de la relación entre CPF ES (k) con la riqueza de especies (S), tal como se demuestra en la investigación aquí documentada.
La relación entre cobertura aérea y riqueza de especies puede determinarse para vegetación clímax no perturbada (Sánchez-Sánchez et al., 2021), aunque su generalización hacia estados de sucesión o perturbación es una tarea pendiente.
El criterio de utilizar el número máximo de especies para caracterizar un tipo de vegetación y uso del suelo establece una condición más orientada al equilibrio, aún en estados de sucesión, ya que considera vegetación cuya relación de cobertura con la riqueza de especies es óptima. Se espera una alta variabilidad observada entre los datos experimentales con relación al ajuste de los modelos potenciales, dado que los criterios de definición de los estados de sucesión son cualitativos (dominancia de estratos) y varían durante el tiempo de la sucesión vegetal, además de estar sujetos a cierto nivel de caracterización subjetiva de la sucesión.
La dinámica de la evolución de la relación entre la cobertura y el número de especies es resultado del balance entre los estratos superior e inferior (Figura 4) del follaje de la vegetación.
La relación entre CPF-S refleja el balance hídrico de los ecosistemas y las trayectorias de la sucesión forestal, así como de otros tipos de vegetación. Aunque no se midieron variables relacionadas con el balance hídrico, los resultados soportan indirectamente su dinámica.
Conclusiones
Los modelos que explican la relación entre la cobertura proyectiva foliar del estrato superior e inferior de la vegetación con relación a la riqueza de especies de plantas vasculares propuestos en este trabajo, son congruentes con las relaciones citadas en la literatura.
La relación entre los parámetros de los modelos potenciales permite simplificar las estimaciones, ya que se requiere solo de un valor para su parametrización, dadas las relaciones entre los parámetros. La relación entre los dos parámetros es consecuencia de que un número de curvas, transformadas linealmente, se interceptan en un punto común.

