










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.13 no.69 México ene./feb. 2022 Epub 09-Mayo-2022
https://doi.org/10.29298/rmcf.v13i69.1093
Scientific article
New species of Monoctenus (Hymenoptera: Diprionidae) attacks Juniperus flaccida Schltdl. (Cupressaceae) in Guerrero, Mexico
1Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Campo Experimental Pabellón. México.
2 Secretaria de Bienestar, Delegación Guerrero. México.
3Comisión Nacional Forestal en Guerrero. México.
Monoctenus is a genus of sawfly of the Diprionidae family that feed on the leaves of Cupressaceae, mainly Juniperus spp. In 2017, Monoctenus sp. was detected, attacking white cedar in 420 ha in Ixcateopan de Cuauhtémoc, Guerrero. In order to know its biology, and to determine whether it was one of the species already recorded for this genus, its phenology was followed during one life cycle, and tests were carried out for molecular identification. In the field, the adults were observed from July to August, while larvae appeared in July-September; the formation of the cocoons started in October, and pupae were formed until April. Only one generation occurs per year, and the species differs from the others in the maculation of the adult face (M. sanchezi) and the length of the rami on the antennae (M. sadadus). Enough information was obtained to consider it as a new species, named Monoctenus cuauhtemoci n. sp. in honor of the Aztec emperor.
Key words Phylogenetic tree; white cedar; phenology; Monoctenus cuauhtemoci n. sp. De Lira; 2021; sawfly; molecular tests
Monoctenus es un género de “mosca sierra” de la familia Diprionidae que se alimenta de las hojas de árboles pertenecientes a la familia Cupressaceae, principalmente Juniperus spp. En 2017, se detectó a Monoctenus sp. atacando a cedro blanco en 420 ha ubicadas en el municipio Ixcateopan de Cuauhtémoc, Guerrero. Para conocer su biología y determinar si correspondía a una de las especies citadas para el género, se siguió la fenología durante un ciclo y se realizaron pruebas para su identificación molecular. En campo, los adultos se observaron de julio-agosto, con la aparición de las larvas en el periodo de julio-septiembre; la formación de los cocones se registró a partir de octubre y las pupas se formaron hasta abril. Solo se presenta una generación por año y difiere de los taxones previamente identificados en la maculación de la cara de los adultos (M. sanchezi) y en la longitud de los ramis en las antenas (M. sadadus). El análisis molecular corroboró la identificación morfológica del material procedente de Ixcateopan de Cuauhtémoc. Se obtuvo suficiente información para considerar a los individuos estudiados como una nueva especie, la cual se denominó: Monoctenus cuauhtemoci n. sp., en honor al emperador azteca.
Palabras clave Árbol filogenético; cedro blanco; fenología; Monoctenus cuauhtemoci n. sp. De Lira; 2021; moscas sierra; pruebas moleculares
Introduction
Conifer "sawflies" (Hymenoptera: Diprionidae) are insects that cause defoliation in trees of the genera Pinus L., Juniperus L., Cupressus L., Abies Mill., Larix Mill., and Pseudotsuga Carrière in temperate forests (Smith, 1988; 1993). This family comprises two subfamilies: Diprioninae and Monocteninae; the former attacks Pinus, while the latter has been cited in association with Juniperus and Cupressus and includes the genus Monoctenus Dahlbom 1835, of which twelve species are known: three in Europe, five in Japan and four in North America (Taeger and Blank, 2008).
In Mexico, until 1974, M. sadadus Smith was recorded in Durango and Veracruz (Smith, 1974). Subsequently, in 2007, in the Sierra de Álvarez in Armadillo de los Infante, San Luis Potosí, (22 °11´53´´ N; 100° 37´03´´ W; 1 840 masl) 20 ha of white cedar were recorded as affected by Monoctenus sp., an infestation that increased to 200 ha in 2008. In 2010, it was determined to be a new species, identified as M. sanchezi Smith (Smith et al., 2010).
In Ixcateopan de Cuauhtémoc, Guerrero (18°32'5.56'' N; 99°47'15.6'' W, 1 741 masl) during 2017 Monoctenus sp. was detected in 460 ha of white cedar (J. flaccida). However, when comparing the adults obtained in that locality with those of San Luis Potosí, external morphological differences were observed, as with M. sadadus. In addition, M. sanchezi and the species under study also differed in phenology.
According to Smith et al. (2010), adults of M. sanchezi occur from June to the end of July, and larvae occur from July to November; whereas cocoons built by the larvae in order to pupate are observed from November to December, and pupation occurs in June. This study presents the phenology and description of a new species of Monoctenus from Ixcateopan de Cuauhtémoc, Guerrero, Mexico.
Materials and Methods
Area of study of phenology and collection of biological material
Phenology monitoring began in January 2017 and continued until September 2019 in La Barranca, Ixcateopan de Cuauhtémoc, Guerrero (18°32´5.56´´ N; 99°47´15.6´´ W, 1 741 masl), in order to determine when the different developmental stages of Monoctenus occur in J. flaccida.
Fortnightly visits were made at ten points within La Barranca, where the presence of male and female adults differentiated by sexual dimorphism (presence of bipectinate antennae in males) was registered. In addition, the foliage of the cedar trees was checked for larvae, and the soil was removed to search for active (presence of green larvae and pupae) and inactive (with sawfly adult emergence holes, damaged by parasitoids, entomopathogens or rodents) cocoons. It is worth mentioning that no eggs were observed, as it is difficult to determine their presence in the field (González and Sánchez, 2018).
Larvae were collected (September 8, 2017) from the foliage of white cedars, and cocoons, from the ground of affected trees (January 25, 2018 and May 29, 2019). Both were confined in 4.0 L plastic cans and transported in coolers to the Agricultural and Forest Health Laboratory of the Pavilion Experimental Field (Cepab) of INIFAP, in Pabellón de Arteaga, Aguascalientes. The larvae were fed on leaves of their natural host. Cocoons obtained from larval rearing or collected from the soil were placed in 250 mL plastic bottles with organza lids and placed in a Binder 720 KBW (E5.1) temperature-controlled chamber at 26 ±2 ºC, with a photoperiod of 14:10 hours light and dark, respectively. Twice a week, the buds were sprayed with water using a 1.0 L hand sprayer to prevent dehydration. The emergence of adults or parasitoids was monitored three times a week.
Additionally, on August 15, 2019, larvae were collected in Llano Grande, Xichú, Guanajuato (21°16'9.9'' N; 100°03'13.0'' W, 2 100 masl), a locality where the health liaison of the National Forestry Commission (Conafor) was informed of the presence of sawflies affecting 150 ha of J. flaccida, since November 2017.
Groups of larvae from both Ixcateopan de Cuauhtémoc and Xichú were immersed in hot water at 70 °C for three minutes, and then fixed in 70 % alcohol (González et al., 2014) for morphological description and photographing with a cell camera through the objective of a Motic dissecting microscope at 40X magnification. The characteristics for the differentiation of specimens at the larval level were based on the publications of Ross (1955), Yuasa (1922), Wong and Szlabey (1986), and González et al. (2014).
The adults emerged at Cepab were morphologically identified based on the descriptions of Monoctenus spp. adults by Smith et al. (2010) and the terminology of Goulet (1992). The ovipositor lancet of the females was extracted after a cold maceration process. The abdomen was separated from the body and placed in a 4 mL Eppendorf tube with 10 % KOH for 24 h. After the maceration, the abdomen was rinsed with distilled water and dehydrated in 70 % alcohol, and temporary mounts were made in glycerin. The structures were photographed with a Motic BA 200 compound microscope, and a complementary photograph of the lancet was obtained with a Scanning Electron Microscope (SEM, EVO MA15, Zeiss Instrument). No genitalia of male specimens were mounted because there were no specimens for extraction.
A group of five Monoctenus sp. larvae from Ixcateopan de Cuauhtémoc, Guerrero, and two M. sanchezi larvae collected in Armadillo de los Infante, San Luis Potosí, preserved in 70 % alcohol, and six adult females of M. sanchezi from Armadillo de los Infante, San Luis Potosí, with an emergence date of June 19, 2012, and five female Monoctenus sp. larvae from Ixcateopan de Cuauhtémoc, Guerrero, with the emergence date 8 October, 2018, were sent to the National Center for Disciplinary Research in Conservation and Improvement of Forest Ecosystems (Cenid-Comef) in Mexico City for molecular identification.
Specimens in 70 % alcohol were washed three times with sterile distilled water and placed in 1.5 mL Eppendorf tubes; liquid nitrogen was added for maceration, and 500μL of lysis buffer were added. The tube was allowed to stand at room temperature for 30 min and centrifuged for 1.0 min at 12 000 rpm; the supernatant was recovered in a new tube, to which 300 μL of phenol:chloroform 1:1 were added; it was placed in the vortex for one minute and centrifuged at 12 000 rpm for 3 min. The supernatant was transferred to another tube, and 200 μL of 1:1 phenol:chloroform were added. The supernatant was transferred to a new tube, and 300 μL isopropanol and 30 μL ammonium acetate were added, lightly mixed and centrifuged for 5 min at 12 000 rpm; the supernatant was decanted, and the pellet was washed with 50 μL of 70 % ethanol. The pellet was dried at room temperature and resuspended in 50 μL of nuclease-free water. The quality of the DNA was measured with a Thermo Scientific NanoDrop 2000 spectrophotometer. The product was kept at a temperature of ± 4 °C.
The universal primers LCO1490 (5'-GGTCAACAAATCATAAAGATATTATTGG-3') and HCO2198 (5'TAAACTTCAGGGTGACCAAAAAAAATCA-3') (Folmer et al., 1994) were used for the amplification of ribosomal DNA regions, which recognize the Cytochrome Oxidase, Subunit I gene (COI) gene. PCR reactions were performed by adding the following reagents: PCR Buffer (5x: 5.0 µL concentration: quantity, respectively), MgCl2 (25mm: 0.75 µL), dNTP (10mm: 0.25 µL), Primer 1 (10 nmol: 1.0 µL), Primer 2 (10 nmol: 1.0 µL), DNA Taq polymerase (5u µL-1: 0.5 µL), target DNA (20 ng µL-1: 3.0 µL), and 13.5 µL PCR grade water.
Amplification was performed on a Biorad T100 thermal cycler using the following program: 1) initial denaturation 1 cycle of 3.0 min at 95 °C; 2) denaturation 1.0 min at 95 °C; 3) alignment 5 cycles of 30 sec. at 51 °C; 4) extension 1.0 min at 70 °C; 5) denaturation 1.0 min at 94 °C; 6) alignment 35 cycles of 30 s at 51 °C; 7) extension 1.0 min at 72 °C; 8) final extension 1 cycle of 5.0 min at 72 °C, and 9) Storage at 4.0 °C. The amplified fragments were run on 1 % agarose gels with 0.4 μL of ethidium bromide and visualized in an Infinitity-ST5 Vilber Lourmat photodocumenter.
The band of interest was cut for purification with the WIZARD® SV Gel and PCR Clean-Up System kit, following the supplier's protocol (Promega Corporation, 1999). 50 µL of solution 1 were added to the agarose gel preserved in Eppendorf tubes, dissolved at 65 °C. The diluted gel mix was transferred to a minicolumn inserted into a collecting tube and incubated at room temperature for 1.0 min; it was then centrifuged at 14 000 rpm for 1.0 min, and the supernatant in the collection tube was decanted. Subsequently, two washes were performed with 700 µL of solution 2 and centrifuged at 14 000 rpm for 3.0 min, and the supernatant from the collecting tube was decanted. The minicolumn was transferred to a sterile Eppendorf tube and allowed to dry at room temperature. 50 µL of nuclease-free water were added, incubated at room temperature for 5.0 min, and centrifuged at 14 000 rpm for 1.0 min; the minicolumn was discarded, the product was stored at 4 °C, and the nuclease-free water was added. Purified PCR products were sent for sequencing to Macrogen Korea in Seoul, Republic of Korea.
The sequences were assembled in the SeqMan Pro module of the DNASTAR Lasergene Molecular Biology 16 program, and each assembly was subjected to BLAST (Basic Local Alignment Search Tool) homology analysis against the National Center of Biotechnology Information nucleotide database (https://blast.ncbi.nlm.nih.gov/Blast.cgi); they were also analyzed in the BOLD SYSTEMS database.
A phylogenetic analysis was carried out for the genus Monoctenus, including the sequencing assemblies of specimens from the collections and the reference databases resulting from the search in the Gen Bank. A phylogenetic analysis was carried out for the genus Monoctenus, including the sequencing assemblies of specimens from the collections and the reference databases resulting from the search in the Gen Bank (Darriba et al., 2019). Phylogenetic reconstruction was performed with Bayesian inference using Markov Chains Monte Carlo (MCMC), implemented in the BEAST v1.10.4 software (Suchard et al., 2018) with 1 000 000 generations. It is worth mentioning that the GenBank sequence HM114320 corresponding to Sirex noctilio Fabricius, 1793 was used as an outgroup control (Hymenoptera: Siricidae).
Results and Discussion
Monoctenus cuauhtemoci n. sp. De Lira, 2021
Distribution. Mexico, Ixcateopan de Cuauhtémoc, Guerrero; 18°3’26.06’’ N; 99°47’19.4’’ W, 2 136 masl.
Host. white cedar Juniperus flaccida Schltdl. growing in the mountains of Cuauhtémoc, Guerrero. A total of 460 ha were affected in 2017 and by 2020, the affected area will increase to 4 320 ha.
Data on its phenology and natural enemies are available at De Lira-Ramos et al. (2019) and González et al. (2019).
Etymology. in honor of the last Aztec emperor, originally from the municipality of Ixcateopan, where the attack by this species of Monoctenus occurred, and the mountain range where the infestations are located is known as the Cuauhtémoc mountain range.
Type Material. Holotype. Dry mounted female, labeled as Mexico, Ixcateopan de Cuauhtémoc, Guerrero. “La Barranca” 18°3’26.06’’ N; 99°47’19.4’’ W, 2 136 masl; Collector E. González, G. of October 8, 2018 (Figures 1A at 1D). Dry mounted male, same data (Figure 2). Deposited in the Forest Insect Collection of Cenid-Comef, INIFAP, Viveros de Coyoacán, Mexico City.

Photographs taken at 2X (A, B and C) and 4X (D).
Figure 1 Female Monoctenus cuauhtemoci n. sp. De Lira, 2021 attacking Juniperus flaccida Schltdl. in Ixcateopan de Cuauhtémoc, Guerrero, Mexico. Dorsal (A), ventral (B), lateral (C); head (D).

Photomicrographs taken at 10X (A and C) and 40X (B and D). Glycerin mounting.
Figure 2 Lancet of the ovipositor of the female Monoctenus cuauhtemoci n. sp. de Lira, 2021; complete lancet (A) and close-up of the apex (B) and Monoctenus sanchezi; complete lancet (C), and close-up of the apex (D).
Paratypes. Three females with the same collection data. Deposited in the forest insect collection of Cepab, belonging to INIFAP, at km 32.5 of the Aguascalientes-Zacatecas highway, Pabellón de Arteaga, Aguascalientes, Mexico.
Larvae. Collection of sawfly larvae preserved in 70 % alcohol with label GROM2. Forestry Training Center (Cefofor) of Conafor. Ciudad Guzmán, Jalisco, Mexico; and collection of sawfly larvae in 70 % alcohol with GROM2 label from Cepab, under INIFAP, at km 32.5 of the Aguascalientes-Zacatecas highway. Pabellón de Arteaga, Aguascalientes, Mexico.
Description. Female, most of the body is black; part of the head and the pro- and mesothorax segments are orange (Figure 1A); black antennae formed by 15 antennomeres, 2 to 9 have large rami, but do not reach twice the length of the base as in M. sadadus (Figure 1C). The length of the scape is not greater than its width, and that of the third antennal segment is more than twice the width. The ocellar area has a black spot that joins with two semicircular spots that reach the middle and upper inner portion of the inner orbits of the compound eyes, encompassing the antennal sockets (Figure 1D). On the outer sides of the antennal sockets there is an orange area that interrupts the stain on the sides of the sockets and on the lower inner portion of the compound eyes, clypeus, and labrum. The prescutum, tegula, and middle and lateral lobes of the mesoscutum are of an orange hue, and the mesopleura is black. Black legs with lighter femur apex and tibia base on the middle and hind legs (Figure 1B). In the front legs, the tibia is yellowish brown in its entirety, and the femur has a yellowish apical third. The wings are dark, with light brown from the base to the mid-wing, and the rest is hyaline; the stigma and the veins are dark brown. The abdomen in dorsal view is shiny black, while laterally only the first segment has a clear portion surrounding the spiracle (Figures 1A and 1B).
In the lancets of the ovipositor of the females of M. cuauhtemoci n. sp. and M. sanchezi differences are observed (Figures 2A and 2C), both species exhibit nine rings and have the basal teeth in the first two rounded; however, rings 3-6 in M. sanchezi are observed to be curved by the greater preponderance of the basal tooth, while they are straighter in M. cuauhtemoci n. sp.). Rings 7-9 at the tip of the lancet in both rings show the basal tooth in forward and backward projection; ring 9 in M. cuauhtemoci n. sp. exhibits five well-defined teeth, and the teeth are not marked in M. sanchezi (Figures 2B and 2D).
Males (Figure 3) with a length of 8 mm, completely dark. Black head with yellow setae, dark brown labrum, jaws and palps. Black antennae with 22 antennomeres, with small rami from the second to the last antennomere. Forelegs with dark brown coxa, trochanter and femur, and with yellowish brown tibia. The second pair of legs with the coxa, trochanter, and femur are dark brown, and the tibia is dark brown in the middle part, with the basal and anterior part of a yellowish-brown hue. The third pair of legs with dark brown coxa, trochanter, and femur, and light brown tibia and first tarsomere in the anterior part, and dark brown basal part. The tegula is dark brown, almost black. The median and lateral lobes of the mesoscutellum are black without spots.

Under field conditions, adults were observed from July 18 to August 24 (Figure 4), reducing their presence until September 4, one month later than that reported for M. sanchezi in San Luis Potosí. In total, 187 females and only two males were observed, which coincides with the data from San Luis Potosí, where a low presence of males was observed. With the dissection of the cocoons, it was determined that the change from larvae to pupae occurred from April 27, with the highest presence from July 27 to August 10, which coincides with the emergence of adults and with the decrease of green larvae (diapause) in the cocoons; this represents two months earlier than that recorded for M. sanchezi (Smith et al., 2010).
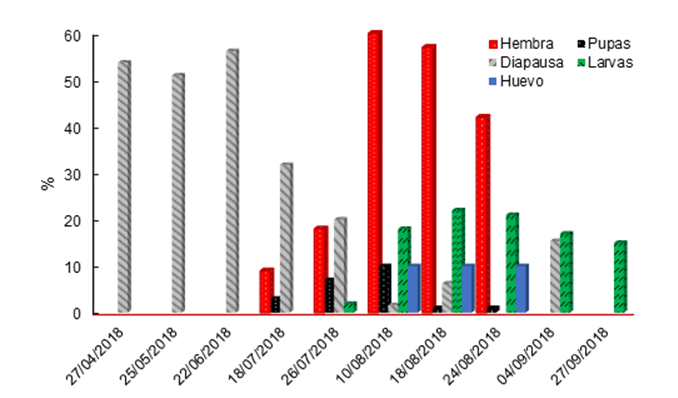
Hembra = Female; Diapausa = Diapause; Huevo = Eggs; Pupas = Pupae; Lavas = Larvae.
Figure 4 Phenology of Monoctenus cuauhtemoci n. sp. De Lira, 2021, in Ixcateopan de Cuauhtémoc, Guerrero, Mexico. Cycle 2018.
Larvae were observed after July 18, with the highest population density on August 10, and were present until September 24. Larval mortality ―the effect of the combat treatment applied on Aug. 25, 2018― was observed after Sept. 4, i.e. 10 days after application. The formation of cocoons was observed after October, and within the cocoons, the larvae remained in diapause (in cocoons in the soil), with higher proportion in November and December; in this condition, they remained until April 9, 2019. This indicates that pupae are formed from the end of April, as was observed in 2018. The data allow inferring that there is only one generation per year, as in San Luis Potosí, although with differences in the time of appearance (Smith et al., 2010). The detection of eggs occurred in August, a stage of development that is difficult to observe with the naked eye, since training is required to identify them.
Of the factors affecting the population of the cocoons in the soil, the most important is mortality caused by parasitoids, followed by predation by rodents and by parasitoid flies of the families Bombyliidae and Tachinidae. Of the entomopathogenic fungi, Beauveria bassiana (Bals.) Vuillemin was the most prevalent, compared to Metarhizium anisopliae (Metsch.) Sorokin (Figure 5).
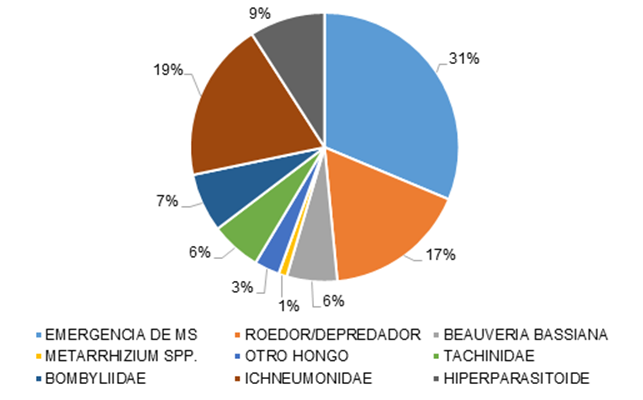
Emergencia = Emergence; Roedor/depredador = Rodent/predator; Otro hongo = Other fungus; Hiperpasitoide = Hyperparasitoid.
Figure 5 Mortality Factors affecting the population of Monoctenus cuauhtemoci n. sp. de Lira 2021, in Ixcateopan de Cuauhtémoc, Guerrero.
During emergence under controlled conditions at Cepab, the most common parasitoids (Figure 6) were those of the family Mantispidae, followed by the Tachinidae, with 85 and 28 individuals, respectively. Although the emergence of only one specimen of Exenterus sp. (Hymenoptera: Ichneumonidae) was recorded, in the field it is common to observe emergence holes caused by parasitoids of the Ichneumonidae family (Ordaz et al., 2013). This is the first time that the family Mantispidae (Insecta: Neuroptera) is cited as a parasitoid of sawflies in Mexico, although the family has been documented in the literature as a parasitoid of hymenopterans (Triplehorn and Johnson, 2005).

Machos = Males; Hembras = Females; No. de individuos = Number of individuals.
Figure 6 Mortality factors affecting the population of Monoctenus cuauhtemoci n. sp. De Lira 2021, (A) and emergence of female sawflies, Monoctenus cuauhtemoci n. sp., under controlled conditions (B).
The larvae of the genus Monoctenus are olive green; their head is light brown with dark longitudinal stripes, and they feed solitarily. In addition, the light brown head has a black spot on the crown suture; the antennae consist of three segments, the last one being conical in shape; the left mandible has more than three teeth; the thoracic legs are dark brown and have seven rings on each one of the abdominal segments. In dorsal view they have a dark longitudinal stripe in the middle portion of the back, with two light stripes on the sides and a dark subdorsal stripe (Figure 7). The difference between M. sanchezi and Monoctenus cuauhtemoci of Ixcateopan is observed in the first pair of thoracic legs, which in M. sanchezi have a completely whitish trochanter, while in Monoctenus cuauhtemoci, the basal portion is dark brown and only the apical portion is whitish.

Photos: Adriana R. Gijón.
Photographs taken at 1X (A, B, D and E) and 2X (C and F).
Figure 7 Appearance of larvae of M. sanchezi Smith (A, B and C) and M. cuauhtemoci n. sp. De Lira, 2021 (D, E and F). Dorsal (A and D), lateral (B and E) and approach to thoracic legs (C and F).
Regarding molecular identification, for the genus Monoctenus there are 34 sequences in the GENBANK corresponding to COI, of which 60 % correspond to Monoctenus juniperi Linneaeus, 1758, and Monoctenus obscuratus Hartig, 1837, both present in Europe; while the rest are genus-specific.
Molecular studies showed that the PCR product band for both larval and adult Monoctenus was 740 bp of the mitochondrial cytochrome C oxidase Subunit I gene.
For this genus, a tree with two well-defined clades was obtained (Figure 8). The first one groups the specimens collected in this study, while the second one corresponds to the sequences of the bases recorded in GENBANK for the European species. In the first clade, M. sanchezi is closely related to specimen GTOM1_35 with 100 % support, which was morphologically identified as M. sanchezi; and the Monoctenus specimens from Ixcateopan de Cuauhtémoc, Guerrero, both larvae (GROM2_2) and adults (GROM2_AD) are classified as a different species.

Discussion
Larvae were obtained from two species of Monocotenus: 1) Sierra de Álvarez in San Luis Potosí and 2) Sierra de Cuauhtémoc in Guerrero. Both are very similar, but differ in the trochanters of the thoracic forelegs, which in M. sanchezi are completely whitish, while in the larvae from Ixcateopan, the trochanter has a dark brown basal portion and a whitish apical portion. At the adult level, maculation on the face of M. sanchezi is very light, covering only the ocellar area and two small circular spots near the inner orbit of the eyes; in M. sadadus Smith and M. cuauhtemoci n. sp., the spot is very large and covers a large part of the face, although they differ in size: in M. cuauhtemoci n. sp. the spots below the ocelli touch the inner orbits of the compound eyes, and in M. sadadus they are smaller. Also, in M. cuauhtemoci n. sp. the spot is interrupted on the sides of the antennal sockets, which is not the case in M. sadadus. Another difference between them is the length of the rami in antennomeres 2 to 9, which in M. sadadus are twice the length of the base, and are shorter in M. cuauhtemoci n. sp. The morphological differences between the populations of Sierra de Álvarez, San Luis Potosí and Xichú, Guanajuato (identified as M. sanchezi) with respect to the population of Sierra de Cuauhtémoc were corroborated by molecular analysis and differences in the lancet of the female ovipositor based on the preponderance of the basal tooth in rings 3 to 6 and the number of teeth in ring nine.
Conclusions
Monoctenus cuauhtemoci n. sp. is a new species detected in the Sierra de Cuauhtémoc that differs from M. sanchezi mainly in the maculation of the face and in the length of the rami of the antennomeres. The Sierra de Cuauhtémoc population is more aggressive than the Sierra de Álvarez, San Luis Potosí and Xichú, Guanajuato populations (identified as M. sanchezi), with up to 200 larvae per tree, compared to 30 in the latter localities.
Acknowledgements
We would like to thank the Town Hall (H. Ayuntamiento) of Ixcateopan de Cuauhtémoc, Guerrero, and the councilors of Ecology Víctor Leyva Guerrero and María Dolores Bustamante Cirilo for the facilities provided; to the M.C. Brenda Torres Huerta for the photographs in Figures 1 A, B and C; 7 A, B, D. We are grateful also for the economic support of the Conacyt-Conafor Sectorial fund, through project CONAFOR 2017 CO2 no. 291304.
REFERENCES
Darriba, D., D. Posada, A. M. Kozlov, A. Stamatakis, B. Morel and T. Flouri. 2019. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Molecular Biology and Evolution 37(1):291-294. Doi: https://doi.org/10.1093/molbev/msz189. [ Links ]
De Lira-Ramos, K. V., E. González-Gaona, E. G. Piza-Nuñez, Y. E. Rodríguez-Cruz y J. C. Gómez-Núñez. 2019. Fenología de Monoctenus spp. (Hymenoptera: Diprionidae) y sus agentes de control natural en Ixcateopan de Cuauhtémoc, Guerrero. In: Memorias del XLII Congreso Nacional de Control Biológico. Sociedad Mexicana de Control Biológico. Veracruz, Ver., México. pp 95‒97. [ Links ]
Folmer, O., M. Black, W. Hoeh, R. Lutz and R. Vrijenhoek. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3(5): 294-299 Molecular Marine Biology and Biotechnology 3(5): 294-299 https://www.researchgate.net/publication/15316743 . (7 de junio de 2020). [ Links ]
González G., E., F. Bonilla T., S. Quiñonez B., G. Sánchez M. F. Tafoya, R., M. P. España L. y S. Robles U. 2014. Guía para la identificación de moscas sierra de la familia Diprionidae presentes en el Centro Norte de México. Publicación Especial 41. INIFAP-CIRNOC-CEPAB. Pabellón de Arteaga, Ags., México. 36 p. [ Links ]
González G., E. y M. Sánchez G. 2018. Identificación y manejo de moscas sierra de la familia Diprionidae presentes en el centro norte de México. Folleto técnico. Conacyt-Conafor. Guadalajara, Jal., México. 122 p. [ Links ]
González, G. E., N. E. Piza, R. K. De Lira, J. C. Gómez, Y. E. Rodríguez y M. G. Sánchez. 2019. Fenología de la mosca sierra (Monoctenus sp.) del cedro blanco (Juniperus flaccida) en Ixcateopan de Cuauhtémoc, Guerrero. INIFAP. Reuniones Nacionales de Investigación e Innovación Pecuaria Agrícola Forestal y Acuícola Pesquera. In: X Reunión Nacional Forestal. Tuxtla Gutiérrez, Chis., México. pp. 221‒222. [ Links ]
Goulet, H. 1992.The genera and subgenera of the sawflies of Canada and Alaska. Hymenoptera: Symphyta. The Insects and Arachnids of Canada, Part 20. Center for Land and Biological Resources Research. Research Branch Agriculture Canada. Publication 1876. Ottawa, Canada. 235 p. [ Links ]
Ordaz S., S., G. Gallegos, M., S. R. Sánchez, P. , M. Flores, D., O. García y M., E. Cerna, C. 2013. Familias de parasitoides de larvas de Monoctenus sanchezi Smith (Hymenoptera: Diprionidae) en la sierra de Álvarez, San Luis Potosí. Soc. Mex. Entomol.: 341-345. http://www.socmexent.org/entomologia/revista/2013/CB/341-345.pdf (7 de junio de 2020). [ Links ]
Promega Corporation. 1999. Wizard® SV Gel and PCR Clean-Up System. Promega Corporation. Technical Bolletin. 13 13 https://worldwide.promega.com/products/nucleic-acid-extraction/clean-up-and-concentration/wizard-sv-gel-and-pcr-clean-up-system/ (7 de junio de 2020). [ Links ]
Ross, H. H. 1955. The taxonomy and evolution of the sawfly genus Neodiprion. Forest Science 1(3): 196-209. https://www.cabdirect.org/cabdirect/abstract/19580500510 (7 de junio de 2020). [ Links ]
Sievers, F. and D. Higgins, G. 2014. Clustal Omega, Accurate Alignment of Very Large Numbers of Sequences. In: Russell, D. (ed.) Multiple Sequence Alignment Methods. Methods in Molecular Biology (Methods and Protocols) Vol. 1079. Humana Press. Totowa, NJ, USA. pp. 105-116. [ Links ]
Smith, D. R. 1974. Conifer sawflies, Diprionidae: Key to North American genera, checklist of world species, and new species from Mexico (Hymenoptera). Proceedings of the Entomological Society of Washington 76(4): 409-418. https://www.researchgate.net/profile/David-Smith-38/publication/260798565_Conifer_sawflies_Diprionidae_Key_to_North_American_genera_checklist_of_world_species_and_new_species_from_Mexico_Hymenoptera/links/5b1f1cb4a6fdcc69745bff2d/Conifer-sawflies-Diprionidae-Key-to-North-American-genera-checklist-of-world-species-and-new-species-from-Mexico-Hymenoptera.pdf . (7 de junio de 2020). [ Links ]
Smith, D. R. 1988. A synopsis of the sawflies (Hymenoptera: Symphyta) of America south of the United States: Introduction Xyelidae, Pamphiliidae, Cimbicidae, Diprionidae, Xyphydriidae, Siricidae, Orussidae, Cephidae. Systematic Entomology 13: 205-261. Doi: https://doi.org/10.1111/j.1365-3113. 1988.tb00242.x. [ Links ]
Smith, D. R. 1993. Systematics, life history and distribution of sawflies. In: Wagner, M. R. and K.F Raffa (eds.) Sawfly life history adaptations to wood plants. Academic Press. San Diego, CA, USA. pp. 3-32. [ Links ]
Smith, R. D., G. Sánchez M. and S. Ordaz S. 2010. A new Monoctenus (Hymenoptera: Diprionidae) damaging Juniperus flaccida (Cupressaceae) in San Luis Potosí, México. Proceedings of the Entomological Society of Washington 112: 444-450. Doi: https://doi.org/10.4289 / 0013-8797.112.3.444. [ Links ]
Suchard, M. A., P. Lemey, G. Baele, D. L. Ayres, A. J. Drummond and A. J. Rambaut. 2018. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10 Virus Evolution 4(1):vey016. Doi:10.1093/ve/vey016. [ Links ]
Taeger, A. and S. M. Blank. 2008. ECatSym -Electronic World Catalog of Symphyta (Insecta, Hymenoptera). Program version 3.9, data version 34 (05.09.2008). Digital Entomological Information. Müncheberg, Germany. https://www.sdei.de/ecatsym/ (7 de junio de 2020). [ Links ]
Triplehorn, C. A. and N. F. Johnson. 2005. Borror and Delong´s Introduction to the study of Insects. Seventh Edition. Thomson Brooks/Cole. Belmont, CA. USA. 864 p. [ Links ]
Wong, H. R. and D. L. Szlabey.1986. Larvae of the North American Genera of Diprionidae (Hymenoptera: Symphyta). Canadian Entomologist 118(6): 577-587. Doi: https://doi.org/10.4039/Ent118577-6. [ Links ]
Yuasa, H. 1922. A classification of the larvae of the Tenthredinoidea. Illinois BiologicalMonographs 7(4):325-490. http://hdl.handle.net/2142/9057 (20 de junio de 2020). [ Links ]
Received: March 08, 2021; Accepted: December 03, 2021
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
