










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.12 no.67 México sep./oct. 2021 Epub 25-Oct-2021
https://doi.org/10.29298/rmcf.v12i67.858
Scientific article
Árboles dañados, crecimiento y fenología después de un huracán en una selva seca en Veracruz

1Instituto de Ecología, A. C., Red de Ecología Funcional. México.
2 Universidad Veracruzana, Instituto de Investigaciones Forestales. México.
Los huracanes son disturbios importantes que afectan el crecimiento y la fenología de los árboles. Durante tres años se registró el crecimiento anual y la fenología mensual de 16 especies arbóreas en una selva seca en Veracruz. En septiembre del 2010, el huracán Karl azotó la región, y un mes después se registró el daño en árboles y se reanudaron las observaciones fenológicas. El daño en árboles fue elevado: 10 % desenraizados, 7 % quebrados, y 2 % doblados. Todos los árboles desenraizados murieron, pero algunos quebrados o doblados rebrotaron (15 % de los árboles etiquetados murieron). El crecimiento diamétrico promedio de árboles sobrevivientes (0.79 cm año-1) fue mayor que el crecimiento pre-huracán (0.68 cm año-1). Para todas las especies juntas, la caída de hojas y producción de hojas y flores no fueron estadísticamente diferentes entre pre- y post-huracán, pero la fructificación fue menor en el año post-huracán. A nivel de especie hubo diferencias en fenología reproductiva entre años. En el año siguiente al huracán, la mayoría no produjeron flores, disminuyeron floración y fructificación (Calyptranthes schiedeana) o no fructificaron (Luehea candida, Maclura tinctoria, Tabebuia chrysantha). Se concluye que debido al daño por el huracán, la mortalidad en árboles fue alta pero muchos árboles se recuperaron; el huracán influyó negativamente en la intensidad de la fenología reproductiva, lo que a su vez puede alterar la estructura de la vegetación, cambiar la composición de especies y afectar las relaciones tróficas y el funcionamiento de toda la comunidad forestal.
Palabras clave: Árboles desenraizados; floración; fructificación; huracán Karl; mortalidad de árboles; rebrotes
As a major disturbance, hurricanes affect growth and phenology of trees. Tree diameters were annually measured for three years, and the phenology of 16 tree species monthly recorded in a seasonally dry tropical forest in Veracruz, Mexico, when on September 2010, Hurricane Karl struck the region. One month later, tree damage was recorded and phenological observations resumed for 12 more months, and diameter measurement for two more years. Tree damage due to the hurricane was high: 10 % were uprooted, 7 % broken and 2 % bent. All trees uprooted died, but some broken or bent trunk trees resprouted (15 % of tagged trees died). Overall, mean diameter growth of trees that survive the hurricane (0.79 cm yr-1) was greater than pre-hurricane growth rate (0.68 cm yr-1). For all the studied species together, leaf fall, leafing, and flowering phenology did not differ between pre- and post-hurricane whereas fruiting was lower for the post-hurricane year. At species level, most species displayed differences in reproductive phenology between pre-hurricane and post-hurricane years. Most species did not flower, lower flowering and fruiting (Calyptranthes schiedeana), or did not fruit (Luehea candida, Maclura tinctoria, Tabebuia chrysantha) the year following the hurricane. In conclusion, due to hurricane damage, tree mortality was high but many trees recovered from damage, and that the hurricane negatively influenced intensity in reproductive phenology, and in turn may alter forest structure, shift species composition, and affect the trophic relationships and functioning of the whole forest community.
Key words: Uprooted trees; flowering; fruiting; Hurricane Karl; tree mortality; sprouting
Introducción
En muchos bosques tropicales estacionalmente secos (SDTF, por su acrónimo en inglés) de la zona Neotropical, los huracanes son eventos de disturbio importantes. Se ha documentado cómo afectan la estructura del bosque, las condiciones ambientales y las interacciones bióticas (Whigham et al., 1991; Zimmerman et al., 1994; Koptur et al., 2002; Van Bloem et al., 2003; Angulo-Sandoval et al., 2004; Bonilla-Moheno, 2012; Jiménez-Rodríguez et al., 2018; Stan y Sánchez-Azofeifa, 2019). Varios autores citan que las tasas de crecimiento de los árboles supervivientes aumentan después de los huracanes (Whigham et al., 1991; Tanner y Bellingham, 2006; Tanner et al., 2014). Este incremento en el crecimiento de las plantas, probablemente, se deba a la precipitación extra estacional que caracteriza a dichos fenómenos meterológicos, lo que es importante para los ecosistemas con agua restringida (Parker et al., 2018).
En Jamaica, Tanner y Bellingham (2006) observaron que las tasas de crecimiento del diámetro del tallo aumentaron durante la década del huracán Gilberto, y que la tasa de crecimiento se duplicó en la década posterior. En Yucatán, Whigham et al. (1991) también registraron que durante el primer año después de ese huracán, el crecimiento relativo del diámetro de la mayoría de las especies fue mayor que el crecimiento promedio de los años previos a Gilberto. Estos fenómenos también modifican la fenología en los bosques, multiplican la producción de follaje y disminuyen tanto la herbivoría (Koptur et al., 2002; Angulo-Sandoval et al., 2004), como la disponibilidad de alimentos para los vertebrados (Wunderle, 1999; Schaffner et al., 2012; Renton et al., 2018). Además, los patrones de fructificación se desfasan, se reducen o incluso se suprimen por completo después de los huracanes, como se registró en Puerto Rico (Wunderle, 1999); la Península de Yucatán, México (Schaffner et al., 2012); y en la Reserva de la Biosfera Chamela Cuixmala en la costa de Jalisco (Renton et al., 2018).
Las regiones tropicales donde los huracanes solían ser raros están experimentando un incremento de los eventos meteorológicos extremos. El calentamiento global, probablemente, disminuirá la frecuencia de todas las tormentas, pero se prevé que la frecuencia de las tormentas más intensas (huracanes) aumente en el Golfo de México (Biasutti et al., 2012). El 14 y 18 de septiembre de 2010, el huracán Karl (categoría 3) impactó el SDTF del centro de Veracruz, México y provocó intensos desastres en toda la región (Stewart, 2011). Sin embargo, antes de Karl, el último acontecimiento registrado de ese tipo data de 1932, cuando un huracán de categoría 3, sin nombre, azotó la región (Gómez, 2006).
Los objetivos del presente estudio fueron examinar el impacto del huracán Karl en la mortalidad de los árboles y la forma en que estos murieron, así como comparar el crecimiento del diámetro y los patrones fenológicos observados en un bosque tropical estacionalmente seco, antes y después del huracán.
Materiales y Métodos
El estudio se realizó en un bosque tropical estacionalmente seco ubicado en el centro de Veracruz, México (19°16'N, 96°29' O, altitud: 97-117 m, área: 21 ha). La precipitación media anual total es de 932 mm y la mayor parte de la lluvia ocurre entre junio y octubre; la temperatura media anual es de 25 °C. Las especies arbóreas dominantes son Bursera cinerea Engl., B. fagaroides (Kunth) Engl., B. simaruba (L.) Sarg., Calyptranthes schiedeana O. Berg, Heliocarpus donnellsmithii Rose, Stemmadenia pubescens Benth. y Tabebuia chrysantha (Jacq.) G. Nicholson (Williams-Linera y Lorea, 2009). La precipitación y temperaturas mensuales se obtuvieron de la estación meteorológica más cercana, ubicada a 7 km del sitio de estudio.
A lo largo de un transecto de <1 m de ancho que cruzaba el fragmento del bosque, y a 30 m del borde de este, se eligió al azar un árbol cerca del transecto, y se marcó el vecino más cercano de cada árbol consecutivo, hasta tener 165 individuos pertenecientes a 16 especies comunes. Se marcaron entre 3 y 19 individuos por especie (Cuadro 1). El estado fenológico se observó en cada árbol desde el suelo con ayuda de binoculares (Swift, SeaHawk 7x50 No. 753). Se estimó, visualmente, tanto el porcentaje de copa sin hojas, como el correspondiente a la presencia de hojas, flores o frutos; y se definieron las siguientes categorías: 0 (0 %) 1 (1 - 25 %), 2 (26 - 50 %), 3 (51 - 75 %) y 4 (76 - 100 %). Estos valores se promediaron sobre los individuos de cada especie por mes y se utilizaron como Índice Fenológico de Intensidad. Se realizaron observaciones fenológicas mensuales desde octubre de 2007, hasta septiembre de 2010, solo unos días antes de que se presentara el huracán Karl (Williams-Linera y Álvarez-Aquino, 2016). A principios de noviembre de 2010, se reanudaron las observaciones fenológicas mensuales por un año más (octubre de 2011).
Cuadro 1 Especies y familias de árboles en el bosque seco tropical de Veracruz, México.
| Familias/Especies | Acr | WD | No. | UR | BE | BR | Muertos |
|---|---|---|---|---|---|---|---|
| Bignoniaceae | |||||||
| Tabebuia chrysantha (Jacq.) G. Nicholson | Tab | 0.33 | 19 | 0 | 0 | 1** | 0 |
| Bixaceae | |||||||
| Cochlospermum vitifolium (Willd.) Spreng. | Coc | 0.48 | 14 | 0 | 2 | 0 | 2 |
| Burseraceae | |||||||
| Bursera cinerea Engl. | Buc | 0.35 | 11 | 0 | 1 | 0 | 1 |
| Bursera simaruba (L.) Sarg. | Bus | 0.78 | 9 | 0 | 1 | 0 | 1 |
| Convolvulaceae | |||||||
| Ipomoea wolcottiana Rose | Ipo | 0.22 | 11 | 1 | 2 | 1** | 3 |
| Fabaceae | |||||||
| Gliricidia sepium (Jacq.) Kunth ex Walp. | Gli | 0.58 | 3 | 1 | 0 | 0 | 1 |
| Leucaena lanceolata S. Watson | Leu | 0.53 | 7 | 0 | 0 | 0 | 0 |
| Senna atomaria (L.) H. S. Irwin & Barneby | Sen | 0.28 | 4 | 2 | 0 | 0 | 2 |
| Hernandiaceae | |||||||
| Gyrocarpus jatrophifolius Domin | Gyr | 0.16 | 7 | 0 | 0 | 0 | 0 |
| Malvaceae | |||||||
| Ceiba aesculifolia (Kunth) Britt. & Baker f. | Cei | 0.44 | 10 | 0 | 0 | 0 | 0 |
| Guazuma ulmifolia Lam. | Gua | 0.94 | 10 | 0 | 0 | 0 | 0 |
| Heliocarpus donnellsmithii Rose | Hel | 0.91 | 16 | 7 | 3* | 1** | 8 |
| Luehea candida (DC.) Mart. | Lue | 0.76 | 15 | 1 | 1** | 0 | 1 |
| Meliaceae | |||||||
| Trichilia trifolia L. | Tri | 0.80 | 8 | 0 | 0 | 0 | 0 |
| Moraceae | |||||||
| Maclura tinctoria (L.) D. Don ex Steud. | Mac | 1.04 | 10 | 4 | 1** | 1 | 5 |
| Myrtaceae | |||||||
| Calyptranthes schiedeana O. Berg | Cal | 0.80 | 11 | 1 | 0 | 0 | 1 |
| Total | 165 | 17 | 11 | 4 | |||
| Muertos en un año | 17 | 7 | 1 | 25 | |||
*Dos sobrevivieron; **Uno sobrevivió; Acr = Acrónimo; WD = Densidad de la madera (g cm-3; Chave et al., 2006); No. = Número de árboles marcados antes del huracán Karl y número de árboles que fueron arrancados de raíz (UR), doblados (BE), quebrado (BR) y muertos en el año siguiente al huracán.
El diámetro del árbol se midió con una cinta diamétrica (Forestry Suppliers, Inc.) en el periodo de junio - julio de 2007 a 2009; en 2010, después del huracán Karl; junio de 2011 y octubre de 2013. Un mes después, en octubre, los árboles en los cuales se hicieron las observaciones fenológicas fueron clasificados como tronco intacto, desenraizado y doblado o quebrado.
Análisis de datos
Las variables de respuesta (clima, incremento de diámetro y fenología) se probaron con un análisis ANODE para determinar si diferían entre años. ANODE es similar al análisis de varianza, pero la prueba no asume una distribución normal. Las diferencias en el incremento de diámetro antes y después del huracán, y las diferencias en la precipitación anual y las temperaturas máximas y mínimas entre años se analizaron utilizando modelos lineales generalizados (GLM) con distribución normal y función de enlace logarítmico. Las diferencias entre años para cada intensidad de fenofase para todas las especies juntas se probaron usando GLM. Las proporciones de fenofase se ajustaron asumiendo una distribución binomial para la variable de respuesta y la función de enlace logit.
El modelo para el incremento de diámetro incluyó especies, año (antes y después del huracán) y el término de interacción. En el modelo de clima y fenología solo se consideró el año. Cuando se detectaron diferencias significativas, se emplearon contrastes ortogonales planificados para determinar, si el año posterior al huracán era diferente de cada uno de los tres años anteriores. Los datos se analizaron con JMP, v10.0.0 (SAS Institute, 2012).
Resultados
Clima
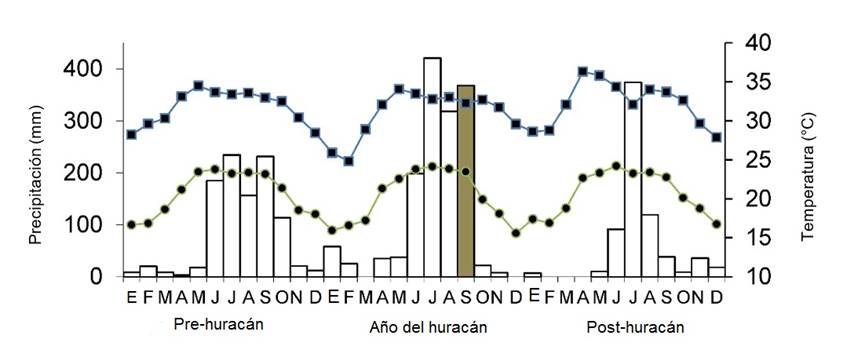
En el año del huracán Karl (2010), la precipitación fue mayor (1 440 mm) que el promedio (932 mm) de la región (Figura 1). Sin embargo, la precipitación (Χ 2 = 1.84, P = 0.60), la temperatura máxima (Χ 2 = 1.81, P = 0.61) y la temperatura mínima (Χ 2 = 0.23, P = 0.97) no difirieron estadísticamente de las variables meteorológicas registradas durante el periodo anterior al huracán (2007 - 2009) y al año posterior (2011) (Figura 1).
Daño y mortalidad de árboles
El huracán Karl defolió todos los árboles, pero los daños variaron entre las especies. Del total de individuos marcados, 32 resultaron dañados; algunos de ellos murieron, mientras que otros rebrotaron de tres a 12 meses después del huracán (Cuadro 1). Entre los árboles muertos, 17 fueron de los desenraizados, siete doblados y uno quebrado. Rebrotaron cinco especies con troncos doblados y quebrados; mientras que, todos los árboles desenraízados murieron (Cuadro 1).
La mortalidad más alta ocurrió en lo taxones con densidad de madera relativamente alta: Heliocarpus donnellsmithii, Maclura tinctoria (L.) D. Don ex Steud. (Cuadro 1), pero otros taxa con madera de densidad baja presentaron un número intermedio de árboles dañados: Ipomea wolcottiana Rose, Senna atomaria (L.) H.S. Irwin & Barneby. Algunas especies de sucesión tardía y madera de alta densidad resultaron parcialmente dañadas y sobrevivieron al impacto del huracán, como Leucaena lanceolata S. Watson y Trichilia trifolia L.; Tabebuia chrysantha tuvo un individuo con un tronco roto que rebrotó (Cuadro 1).
Crecimiento en diámetro
En general, hubo diferencias significativas entre las tasa de crecimiento anterior (0.68 + 0.09 cm año-1) y posterior al huracán (0.79 ± 0.10 cm año-1) (Χ 2 = 5.12, P = 0.024) y entre especies (Χ 2 = 66.36, P <0.0001, Figura 2). Sin embargo, la interacción entre los taxones y el efecto del huracán no fue significativo (Χ 2 = 17.16, P = 0.25). Las tasas de crecimiento del diámetro antes y después del huracán variaron de 0.33 a 1.63 cm año-1 y de 0.30 a 1.89 cm año-1, respectivamente (Figura 2). Luehea candida (DC.) Mart. Registró el mayor crecimiento en diámetro, seguida de Guazuma ulmifolia Lam. y Senna atomaria.

Figura 2 Tasa de crecimiento diametral media registrada anualmente durante los tres años anteriores y los dos años posteriores al huracán Karl. Los acrónimos de las especies se indican en el Cuadro 1.
Fenología
A nivel de la comunidad, el estadio sin hojas, la producción de follaje y la floración evidenciaron patrones anuales cíclicos (Figura 3a, b, c). Se presenta la fenología de los 3 años previos al huracán (Figura 3) para determinar, si los datos registrados después del huracán corresponden con la variación de un año a otro. Durante la estación seca se observa la falta de hojas, y la presencia de estas, en todos los árboles, en los meses de julio - septiembre (Figura 3a, b). La caída de las hojas, su producción y la floración fueron estadísticamente similares entre los años anteriores y posterior al huracán, pero el pico de la floración cambió después del huracán al periodo de abril - mayo (Figura 3c). La producción de frutos fue mucho menor al año siguiente del huracán (Figura 3d).

Figura 3 Fenolía a nivel comunidad registrada durante los tres años previos al huracán. El año 4 comienza en noviembre de 2010, justo después de la entrada del huracán a Veracruz, México.
A nivel de especies, los patrones fenológicos vegetativos se mantuvieron similares antes y después del huracán. En contraste, la intensidad de las fenofases reproductivas sí fueron afectadas por el huracán (Figuras 3 y 4). Para la mayoría de los taxones (12), la intensidad de la floración disminuyó en el año posterior al huracán, y tres especies no florecieron (Calyptranthes schiedeana, Luehea candida y Tabebuia chrysantha); mientras que en, Bursera simaruba y Guazuma ulmifolia (Figura 4a) la intensidad de la floración fue mayor después del huracán. En el año siguiente al huracán, la fructificación fue nula, en cuatro especies y menor para la mayoría de los taxa, o bien el pico de fructificación se desplazó (Leucaena lanceolata y Senna atomaria); solo Gliricidia sepium (Jacq.) Kunth ex Walp. registró una fructificación superior después del huracán (Figura 4b).
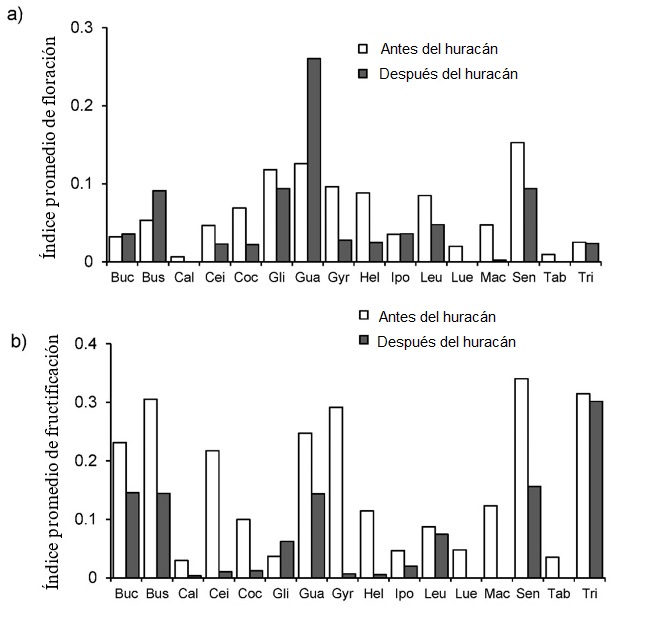
Figura 4 Fenología promedio mensual de floración (a) y fructificación (b) registrada en un bosque seco tropical durante tres años antes y un año después del huracán Karl. Los acrónimos de las especies se indican en el Cuadro 1.
Discusión
Durante el año posterior al huracán, 15 % de los individuos arbóreos perecieron (25 de los 165); cifra superior al intervalo registrado de 7 a 14 % después de un evento de ese tipo en bosques neotropicales (Brokaw y Walker, 1991; Zimmermann et al., 1994; Bonilla-Moheno, 2012; Stan y Sánchez-Azofeifa, 2019). En el área de estudio, la mayoría de los árboles murieron desenraízados, seguidos de aquéllos con el tronco doblado o quebrado, pero la mortalidad no se relacionó con la densidad de la madera. En otros bosques secos donde los huracanes son frecuentes, pocos ejemplares murieron arrancados de raíz, pero los quebrados presentaron la mortalidad más alta (Whigham et al., 1991; Zimmerman et al., 1994; Bonilla-Moheno, 2012).
Algunos autores señalan que las especies de sucesión temprana son más susceptibles a quebrarse que las de sucesión tardía (Zimmermann et al., 1994; Ross et al., 2001). En general, el daño a los árboles ocurre en los ejemplares más altos, ya que podrían ser más vulnerables a daños mecánicos (Jiménez-Rodríguez et al., 2018); por ello, los más grandes registraron más muerte por desenraice y quiebre que los pequeños (Bonilla-Moheno, 2012; García y Siliceo-Cantero, 2019).
La alta mortalidad y el grado de desenraice se relacionaría con el hecho de que Karl fue el primer huracán que llegó a esta región en más de 60 años (Gómez, 2006). Es posible que haya sido el primero en mucho tiempo que dañara los árboles más vulnerables o expuestos. Bonilla-Moheno (2012) refiere que los impactos de los huracanes subsecuentes son menores, comparados con el primero; lo que sugiere que los bosques secos tienen un alto grado de recuperación, en función del nivel de los huracanes subsiguientes.
El crecimiento en diámetro de los árboles después de Karl fue mayor al promedio registrado previamente; esto coincide con lo citado en la literatura (Tanner y Bellingham, 2006; Tanner et al., 2014). Sin embargo, en el presente estudio la gran precipitación observada durante los dos meses anteriores al huracán, también tendría un efecto positivo en el crecimiento del diámetro. Aparentemente, el cambio en la tasa de crecimiento depende de la especie; lo cual se corroboró con la obtención de diferencias significativas entre los taxones evaluados. Tanner et al. (2014) indicaron una variación entre especies, y que el aumento fue superior en algunas que son demandantes de luz (por ejemplo, Alchornea latifolia Sw.). Whigham et al. (1991) afirmaron que las altas tasas de crecimiento relativo pueden reflejar una mayor disponibilidad de nutrientes, ya que la tormenta depositó grandes cantidades de P, K, Ca, Mg y Mn en el suelo del bosque.
La fenología del bosque se estudió durante los tres años previos al huracán (Williams-Linera y Álvarez-Aquino, 2016). Los patrones de producción de hojas se mantuvieron similares antes y después del huracán; pero a nivel de la comunidad, la defoliación causada por Karl afectó el tiempo de estacionalidad de la caída de las hojas y la comunidad permaneció sin hojas durante un mes. La floración fue afectada, porque antes del huracán se apreciaban picos importantes en todos los niveles, pero después de este no hubo estacionalidad. Se han documentado cambios en los picos de floración en el sur de Florida, donde el huracán Andrew la retrasó en un arbusto tropical durante dos meses, aunque una fuerte defoliación también podría relacionarse con una alta floración (Pascarella, 1998).
La fructificación fue la fase fenológica más afectada en el año siguiente al huracán. En Puerto Rico, Wunderle (1999) documentó que el efecto del huracán Hugo se tradujo en una menor producción de frutos y en una asincronía, ya que los árboles tardaron cuatro meses en que el número de especies en fructificación alcanzara un nivel equivalente al mínimo anual normal. La producción de frutos en la Península de Yucatán fue cuatro veces mayor durante la estación seca antes que después de los huracanes (Schaffner et al., 2012). En la Reserva de la Biosfera Chamela-Cuixmala, las flores y frutos se redujeron durante el primer año después de los huracanes Jova y Patricia, y el bosque recuperó los ciclos fenológicos en el segundo año posterior a esos eventos (Renton et al., 2018).
El impacto de los huracanes en la fenología puede tener un fuerte efecto en las relaciones tróficas. En algunos estudios se consigna que el bosque compensa el daño aumentando la producción de hojas, además hay niveles más bajos de herbivoría, probablemente porque el disturbio elimina las poblaciones de insectos herbívoros; lo cual permite que las plantas se recuperen (Koptur et al., 2002; Angulo-Sandoval et al., 2004). Asimismo, la escasez de frutos afecta el comportamiento de otros herbívoros, como los monos araña (Schaffner et al., 2012) y de especies amenazadas como los loros (Wunderle, 1999; Renton et al., 2018).
Los efectos de los huracanes a largo plazo provocan cambios en la composición de las especies (Tanner et al., 2014); en el bosque estudiado la composición puede afectarse, ya que dos especies que no florecieron ni fructificaron fueron Calyptranthes schiedeana y Tabebuia chrysantha, la primer endémica de Veracruz; y la segunda amenazada, según la norma oficial mexicana, NOM-059-SEMARNAT, (Semarnat, 2010).
Conclusiones
Debido a los daños causados por el huracán Karl, la mortalidad de los árboles fue alta, pero muchos ejemplares se recuperaron, probablemente, porque este bosque no había sido muy afectado por huracanes recientes y a su alta resiliencia. Se necesita realizar más investigación para comprender mejor cómo el aumento previsto de huracanes intensos podría influir en la estructura forestal y la fenología en SDTF de la región. Los resultados sugieren que aunque los patrones fenológicos vegetativos pre y poshuracán fueron similares, Karl claramente tuvo un efecto negativo en la intensidad de la fenología reproductiva; lo que a su vez, puede cambiar la composición de las especies y tener un impacto en las relaciones tróficas y el funcionamiento de la comunidad forestal.
Agradecimientos
Los autores agradecen a Marichu Peralta por su valiosa ayuda en el campo y a Don Miguel Morales por el permiso para realizar esta investigación en su predio.
REFERENCIAS
Angulo-Sandoval, P., H. Fernández M. and J. K. Zimmerman. 2004. Changes in patterns of understory leaf phenology and herbivory following hurricane damage. Biotropica 36(1):60-67. Doi: 10.1111/j.1744-7429.2004.tb00296.x. [ Links ]
Biasutti, M., A. H. Sobel, S. J. Camargo and T. T. Creyts. 2012. Projected changes in the physical climate of the Gulf Coast and Caribbean. Climatic Change 112:819-845. Doi: 10.1007/s10584-011-0254-y. [ Links ]
Bonilla-Moheno, M. 2012. Damage and recovery forest structure and composition after two subsequent hurricanes in the Yucatan Peninsula. Caribbean Journal of Science 46(2-3):240-248. doi:10.18475/cjos.v46i2.a12. [ Links ]
Brokaw, N. V. L. and L. R. Walker. 1991. Summary of the effects of Caribbean hurricanes on vegetation. Biotropica 23(4):442-447. Doi:10.2307/2388264. [ Links ]
Chave, J. , H. C. Muller L., T. R. Baker, T. A. Easdale, H. ter Steege and C. O. Webb. 2006. Regional and phylogenetic variation of wood density across 2456 Neotropical tree species. Ecological Applications 16(6):2356-2367. Doi: 10.1890/1051-0761(2006)016[2356:RAPVOW]2.0.CO;2. [ Links ]
García, A. and H. H. Siliceo-Cantero. 2019. Huracán Jova: efecto de un fenómeno meteorológico severo sobre lagartijas insulares en el occidente de México. Revista Mexicana de Biodiversidad 90: e902880. Doi: 10.22201/ib.20078706e.2019.90.2880. [ Links ]
Gómez R., M. 2006. Trayectorias históricas de los ciclones tropicales que impactaron el estado de Veracruz de 1930 al 2005. Scripta Nova. Revista Electrónica de Geografía y Ciencias Sociales. Universidad de Barcelona 10(218): 15. http://www.ub.es/geocrit/sn/sn-218-15.htm (4 de septiembre de 2020). [ Links ]
Jiménez-Rodríguez, D. L., M. Y. Álvarez-Añorve, M. Pineda-Cortes, J. I. Flores-Puerto, J. Benítez-Malvido, K. Oyama and L. D. Ávila-Cabadilla. 2018. Structural and functional traits predict short term response of tropical dry forests to a high intensity hurricane. Forest Ecology and Management 426:101-114. Doi:10.1016/j.foreco.2018.04.009. [ Links ]
Koptur, S., M. C. Rodríguez, S. F. Oberbauer, C. Weekley and A. Herndon. 2002. Herbivore-free time? Damage to new leaves of woody plants after Hurricane Andrew. Biotropica 34(4):547-554. Doi: 10.1111/j.1744-7429.2002.tb00573.x. [ Links ]
Parker, G., A. Martínez Y., J. C. Álvarez Y., M. Maass and S. Araiza. 2018. Effects of hurricane disturbance on a tropical dry forest canopy in western Mexico. Forest Ecology and Management 426:39-52. Doi: 10.1016/j.foreco.2017.11.037. [ Links ]
Pascarella, J. B. 1998. Hurricane disturbance, plant-animal interactions, and the reproductive success of a tropical shrub. Biotropica 30(3):416-424. Doi: 10.1111/j.1744-7429.1998.tb00075.x. [ Links ]
Renton, K., A. Salinas M., R. Rueda H. and L. D. Vázquez R. 2018. Differential resilience to extreme climate events of tree phenology and cavity resources in tropical dry forest: Cascading effects on a threatened species. Forest Ecology and Management 426:164-175. Doi: 10.1016/j.foreco.2017.10.012. [ Links ]
Ross, M. S., M. Carrington, L. J. Flynn and P. L. Ruiz. 2001. Forest succession in tropical hardwood hammocks of the Florida Keys: Effects of direct mortality from Hurricane Andrew. Biotropica 33(1):23-33. Doi: 10.1111/j.1744-7429.2001.tb00154.x. [ Links ]
SAS Institute (2012). JMP. v10.0.0. SAS Institute. Cary, NC, USA. n/p. [ Links ]
Schaffner, C. M., L. Rebecchini, G. Ramos F., L. G. Vick and F. Aureli. 2012. Spider monkeys (Ateles geoffroyi yucatenensis) cope with the negative consequences of hurricanes through changes in diet, activity budget, and fission-fusion dynamics. International Journal of Primatology 33:922-936. Doi: 10.1007/s10764-012-9621-4. [ Links ]
Secretaría del Medio Ambiente y Recursos Naturales (Semarnat).2010. Norma Oficial Mexicana NOM-059-SEMARNAT-2010. Especies nativas de México de flora y fauna silvestres. Categorías de riesgo y especificaciones para su inclusion, exclusion o cambio - Lista de especies en riesgo. Diario Oficial de la Federación. México, D.F., méxico. 78 p. [ Links ]
Stan, K. and A. Sánchez-Azofeifa. 2019. Tropical dry forest diversity, climatic response, and resilience in a changing climate. Forests 10(5):443. Doi: 10.3390/f10050443. [ Links ]
Stewart, S. R. 2011. Tropical cyclone report. Hurricane Karl (AL132010, 14-18 September 2010). https://origin.www.nhc.noaa.gov/data/tcr/AL132010_Karl.pdf (30 de agosto de 2020). [ Links ]
Tanner, E. V. J. and P. J. Bellingham. 2006. Less diverse forest is more resistant to hurricane disturbance: evidence from montane rain forests in Jamaica. Journal of Ecology 94(5):1003-1010. Doi: 10.1111/j.1365-2745.2006.01149.x. [ Links ]
Tanner, E. V. J., F. Rodríguez S., J. R. Healey, R. J. Holdaway and P. J. Bellingham 2014. Long-term hurricane damage effects on tropical forest tree growth and mortality. Ecology 95(10):2974-2983. Doi: 10.1890/13-1801.1. [ Links ]
Van Bloem, S. J., P. G. Murphy and A. E. Lugo. 2003. Subtropical dry forest trees with no apparent damage sprout following a hurricane. Tropical Ecology 44(2):137-145. https://tropecol.com/pdf/open/PDF_44_2/44201.pdf (5 de septiembre de 2020). [ Links ]
Whigham, D. F., I. Olmsted, E. Cabrera C. and M. E. Harmon. 1991. The impact of Hurricane Gilbert on trees, litterfall, and woody debris in a dry tropical forest in the Northeastern Yucatan Peninsula. Biotropica 23(4):434-441. Doi: 10.2307/2388263. [ Links ]
Williams-Linera, G. and F. Lorea. 2009. Tree species diversity driven by environmental and anthropogenic factors in tropical dry forest fragments of central Veracruz, Mexico. Biodiversity and Conservation 18:3269-3293. Doi: 10.1007/s10531-009-9641-3. [ Links ]
Williams-Linera, G. and C. Álvarez-Aquino. 2016. Vegetative and reproductive tree phenology of ecological groups in a tropical dry forest in central Veracruz, Mexico. Botanical Sciences 94(4):745-756. Doi: 10.17129/botsci.682. [ Links ]
Wunderle, J. R. 1999. Pre- and post-hurricane fruit availability: implications for Puerto Rican parrots in the Luquillo Mountains. Caribbean Journal of Science 35(3-4):249-264. https://www.fs.usda.gov/treesearch/pubs/30302 (4 de septiembre de 2020). [ Links ]
Zimmerman, J. K., E. M. Everham, R. B. Waide, D. J. Lodge, C. M. Taylor and N. V. L. Brokaw. 1994. Responses of tree species to hurricane winds in subtropical wet forest in Puerto Rico: implications for tropical tree life histories. Journal of Ecology 82(4):911-922. Doi: 10.2307/2261454. [ Links ]
Recibido: 13 de Septiembre de 2020; Aprobado: 15 de Marzo de 2021
 This is an open-access article distributed under the terms of the
Creative Commons Attribution License
This is an open-access article distributed under the terms of the
Creative Commons Attribution License
