











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
México posee una privilegiada diversidad de ecosistemas naturales, integrados por matorrales xerófilos (41 %), bosques templados (24 %) y selvas (23 %) (Conafor, 2009). Las zonas templadas ocupan alrededor de 46 millones de ha, lo que equivale a 23.4 % del territorio nacional. Se distribuyen, principalmente, en cuatro regiones fisiográficas: Sierra Madre Occidental, Sierra Volcánica Transversal, Sierra Madre Oriental y Sierra Madre del Sur (González, 2003). Estos sistemas montañosos son sitios con alta diversidad de plantas, distribuida entre árboles y el sotobosque; en general, los bosques templados reúnen más de la mitad de las especies de pinos y encinos, además de 3 % de los taxa de Quercus del mundo (Rzedowski, 2006; Rodríguez y Myers, 2010; González-Elizondo et al., 2012).
Determinar la estructura florística, vertical y horizontal, la densidad de individuos dentro de un área boscosa permite conocer la naturaleza de la masa forestal, con base en su diversidad biológica, composición florística, así como la abundancia en la distribución espacial, y en los estratos altitudinales de las especies que forman las comunidades vegetales (Aguirre, 2002).
La diversidad es un sinónimo de la riqueza o variedad de especies que se presentan en un ecosistema (Gaines, 1999; Gadow et al., 2007). La estructura arbórea es un elemento clave para evaluar la estabilidad de los bosques, la cual se puede modificar mediante la aplicación de tratamientos silvícolas que cambian la estructura de los rodales o masas forestales y en consecuencia, el bosque (Lähde et al., 1999; del Río et al., 2003; Castellanos-Bolaños et al., 2010). Por lo tanto, la cuantificación de las variables estructurales de los bosques es importante para entender el funcionamiento del ecosistema y con ello contribuir al manejo forestal sustentable (Castellanos-Bolaños et al., 2010).
El conocimiento de la estructura horizontal y vertical es sustancial para planear acciones que favorezcan el desarrollo de los bosques, y que se realizan al seleccionar los árboles que serán extraídos, sin afectar la estructura original (Aguirre-Calderón, 2015). Primordialmente, la estructura horizontal está dada por la distribución dasométrica (área basal y volumen por categoría diamétrica), así como por la abundancia, frecuencia y dominancia. La estructura vertical utiliza diferentes zonas de altura para la detección de cambios en la diversidad arbórea en diferentes estratos del bosque, con el objetivo de aportar información básica sobre la dinámica del rodal (Pretzsch, 2009).
La composición florística describe al número de familias, géneros y especies en un bosque al momento de realizar un inventario. Los elementos que se consideran para lograrlo se enfocan en la diversidad, riqueza de especies y la similitud entre otras (Louman et al., 2001). Los estudios de la vegetación son uno de los principales soportes para la planificación, manejo y conservación de cualquier ecosistema. Por tal motivo, un inventario florístico planificado debe suministrar información de la riqueza específica (diversidad alfa) (Villarreal et al., 2006), en la que se incluye solamente el número de especies, no la abundancia de cada una de ellas.
El uso de índices de diversidad que ponderan la riqueza y la abundancia de especies proporcionan una validez científica para establecer criterios de conservación, ya que estas son con frecuencia utilizadas como indicadores ambientales de la salud del ecosistema (Magurran, 1989). Esos índices se han aplicado para orientar las acciones relacionadas con la biodiversidad de los hábitats, en virtud de que expresan en valores numéricos la información de censos o muestras que contribuyen a fundamentar la toma de decisiones en el manejo forestal (Lübbers, 1997).
Es por ello, que el manejo forestal responsable requiere de una visión de sustentabilidad, el cual se dictamina con principios y criterios que evalúan el mantenimiento de la diversidad y la conservación de la composición florística. Para todo bosque que es aprovechado y esencialmente para aquel que tenga certificación de buen manejo, se deben conocer las características y el comportamiento del estrato arbóreo (Hernández, 2012). Por lo tanto, el objetivo del presente estudio consistió en describir y comparar la diversidad, estructura y composición de la vegetación en bosques templados ubicados en el sur del estado de Nuevo León, México.
Materiales y Métodos
Área de estudio y sitios de muestreo
El trabajo de campo se llevó a cabo durante el invierno del año 2019, en cuatro localidades de bosque mixto de pino-encino, ubicados en el sistema montañoso de la Sierra Madre Oriental, con una distancia aproximada de 5 km entre ellos; tres se localizan en el municipio Galeana y uno en el municipio Aramberri en el estado de Nuevo León, México; entre los 23°43'51'' - 25°26'45'' N y los 99°28'30'' - 100°55'48'' O (Figura 1).
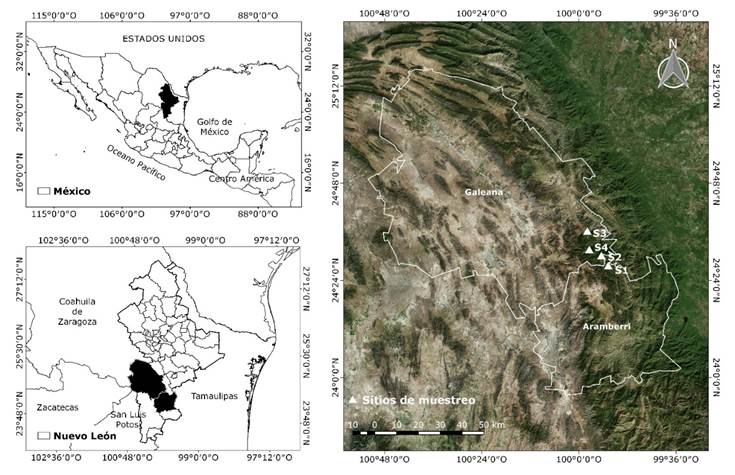
En la región, el clima es templado subhúmedo con lluvias en verano; su régimen pluvial varía entre 600 y 1 200 mm, con temperatura media anual de 14 a 18 ˚C.
Predomina el tipo de suelo Litosol y la vegetación corresponde a bosques de coníferas conformados por asociaciones de pino-encino y encino-pino (Inegi, 2017).
Se establecieron cuatro parcelas de muestreo con una superficie de 2 500 m2 (50 × 50 m), en los parajes Canoas (S1), Agua Blanca (S2), Cañada El Llorón (S3) y El Alamillal (S4); en cada una de ellas se registraron las coordenadas geográficas, sus características fisiográficas (Cuadro 1) y se censaron todos los individuos de las especies arbóreas mayores a 7.5 cm de diámetro normal (diámetro a una altura de 1.3 m sobre el nivel del suelo), conforme a la metodología utilizada en los sitios de investigación forestal y de suelos desarrollado por Corral-Rivas et al. (2009). Se identificó la especie y el número de árbol de cada individuo y se midieron la altura total (h), diámetro normal (d 1.30 ) y diámetro de copa (d copa ).
Cuadro 1 Descripción fisiográfica de los sitios de muestreo.
| Sitios | Paraje | Latitud (N) | Longitud (O) | Altitud (m) | Exposición | Tipo de Vegetación |
|---|---|---|---|---|---|---|
| 1 | Canoas | 24°27'14.62'' | 99°52'44.97'' | 2 665 | Suroeste | Pino-encino |
| 2 | Agua blanca | 24°29'44.81'' | 99°54'26.17'' | 2 442 | Suroeste | Encino-pino |
| 3 | Cañada El Llorón | 24°35'47.76'' | 99°57'55.87'' | 2 133 | Noreste | Encino-pino |
| 4 | El Alamillal | 24°31'12.69'' | 99°57'25.94'' | 2 806 | Noreste | Pino-encino |
Composición Florística
Durante el levantamiento de la información en campo, se recolectó material botánico de todas las especies arbóreas existentes en los sitios de muestreo; además, de cada una se recolectó una porción terminal de una rama, conformada por hojas y la estructura reproductiva. Para su posterior identificación taxonómica, dichos ejemplares se llevaron a la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León. La validez de los nombres científicos se corroboró con base en la plataforma The Plant List (2013).
Coeficiente de similitud
Para determinar la semejanza en la composición de las especies entre las cuatro localidades, se aplicó un análisis con el programa BioDiversity Professional versión 2.0 (McAleece et al., 1997), mediante un algoritmo que permite analizar la similitud de las muestras a través del cálculo del porcentaje con intervalos de 0 a 100 %, cuyo resultado se representa en un dendrograma de similitud-disimilitud de Bray-Curtis (Bray y Curtis, 1957).
Índices de diversidad y riqueza
Se estimaron los siguientes índices de diversidad y riqueza:
El índice de diversidad de Shannon-Weaver (H'), que expresa la heterogeneidad de una comunidad sobre la base de dos factores: el número de especies presentes y su abundancia relativa (Shannon, 1948; Castellanos-Bolaños et al., 2008).
Donde:
S = Número de especies presentes
ln = Logaritmo natural
P i = Proporción de los individuos hallados de la especie i; se calcula mediante la relación (n i /N)
n i = Número de individuos de la especie i
N = Número total de individuos
El índice de diversidad de Simpson (D), que estima si una comunidad determinada está compuesta por especies muy abundantes, ya que suma las abundancias de cada una al cuadrado y así, les da importancia a los taxones con alto valor (Lamprecht, 1962).
Donde:
P i = Proporción de las especies i en la comunidad (n i /N)
n i = Número de individuos de la especie i
N = Número total de individuos
El índice de riqueza de Margalef (D Mg ), que determina la biodiversidad de una comunidad con base en la distribución numérica de los individuos de las diferentes especies, en función del número total de individuos existentes en la muestra analizada. Combina el número de especies (S) y el número de individuos (N) (Magurran, 2004).
Donde:
ln = Logaritmo natural (base e)
S = Número total de especies presentes
N = Número total de individuos
El índice de riqueza de Menhinick (D Mn ), que se basa en la relación entre el número de especies y el número total de individuos observados, el cual se incrementa al aumentar el tamaño de la muestra.
Donde:
S = Número de especies
N = Número total de individuos
Categorías diamétricas
Se utilizó la función de Weibull para crear histogramas de distribuciones diamétricas, los cuales se evaluaron con una prueba de bondad de ajuste de X-cuadrada, con el software Minitab versión 16 (Minitab, 2014). Dicha función se usa en la ciencia forestal por su facilidad de aplicación en análisis semejantes de algunos taxa del género Pinus (Bailey y Dell, 1973).
Estructura horizontal
Para evaluar la estructura horizontal de cada especie, se determinó su abundancia, de acuerdo con el número de árboles, su dominancia en función del área de copa, y su frecuencia con base en la presencia en los sitios de muestreo. Los resultados se utilizaron para calcular el Índice de Valor de Importancia (IVI), que adquiere valores porcentuales en una escala de cero a 100 (Müeller-Dombois y Ellenberg, 1974; Mostacedo y Fredericksen, 2000).
La estimación de la abundancia relativa se obtuvo con la siguiente ecuación:
Donde:
A i = Abundancia absoluta de la especie i (N ha-1)
AR i = Abundancia relativa de la especie i respecto a la abundancia total
N i = Número de individuos de la especie i
S = superficie de muestreo (ha)
La dominancia relativa se calculó mediante la siguiente exresión:
Donde:
D i = Dominancia absoluta de la especie i (N ha-1)
DR i = Dominancia relativa de la especie i respecto a la dominancia total
Ab i = Área de la copa de la especie i
S = Superficie de muestreo (ha)
La frecuencia relativa se estimó con la siguiente ecuacion:
Donde:
F i = Frecuencia absoluta (porcentaje de presencia en los sitios de muestreo)
FR i = Frecuencia relativa de la especie i respecto a la suma de las frecuencias
f i = Número de sitios en el que está presente la especie i
NS = Número total de sitios de muestreo
El índice de valor de importancia (IVI) se definió mediante la ecuación (Whittaker, 1972; Moreno, 2001):
Donde:
IVI = Índice de valor de importancia
AR i = Abundancia relativa de la especie i respecto a la abundancia total
DR i = Dominancia relativa de la especie i respecto a la dominancia total
FR i = Frecuencia relativa de la especie i respecto a la frecuencia total
Estructura vertical (A de Pretzsch)
Se interpretó la estructura vertical de las especies por medio del índice de distribución vertical A de Pretzsch (Pretzsch, 1996; del Río et al., 2003), el cual es una modificación del índice de Shannon (Pretzsch, 2009), en el que A tiene valores entre cero y un valor A max ; cuando un valor A = 0 significa que el rodal está constituido por una especie que ocurre en un solo estrato. A max se alcanza cuando la totalidad de las especies están presentes en la misma proporción, tanto en el rodal como en los diferentes estratos (Corral et al., 2005). Por ello, se definieron tres estratos con intervalos de altura máxima del área, en los cuales el árbol más alto representa 100 %, y corresponde al estrato I: 80 - 100 %; al estrato II: 50-80 %; y al estrato III: de 0-50 % (Aguirre, 2002; Jiménez et al., 2008; Pretzsch, 2009). El índice A se calcula mediante la siguiente ecuación:
Donde:
A = Índice de distribución vertical
S = Número de especies presentes
Z = Número de estratos respecto a la altura
P ij = Porcentaje de especies en cada zona, se estima mediante (n i;j /N), n i
j = Número de individuos de la especie i en el estrato j
N = Número total de individuos
Para comparar el índice de Pretzsch fue necesario estandarizarlo, y esto se realizó con el valor de A max, que se obtuvo con la siguiente ecuación:
Para estandarizar el valor de A se aplicó la siguiente expresión.
Resultados y Discusión
Composición florística
Se identificaron 10 especies de árboles, pertenecientes a cuatro familias y cinco géneros. Fagaceae reunió la mayor riqueza con cinco especies, seguida de Pinaceae con tres. Estas familias representaron 92.32 % del total registrado en los cuatro sitios de estudio (Cuadro 2). Las restantes fueron Ericaceae y Cupressaceae con un género. Destacaron los taxones de los géneros Quercus (cinco), Pinus (dos); de Abies, Arbutus y Juniperus solamente se identificó un taxón. En relación al número de individuos para los cuatro sitios de estudio, dominaron Fagaceae, con 228 y Pinaceae, con 205; mientras que, las de menor abundancia fueron Ericaceae, con 32 y Cupressaceae, con 4.
Cuadro 2 Especies registradas en los sitios de muestreo, ordenados por familia.
| Familia | Nombre científico | Nombre común |
|---|---|---|
| Pinaceae | Abies vejarii Martínez | Oyamel |
| Pinaceae | Pinus pseudostrobus Lindl. | Pino blanco |
| Pinaceae | Pinus teocote Schiede ex Schltdl. et Cham. | Ocote |
| Fagaceae | Quercus canbyi Trel. | Encino amarillo |
| Fagaceae | Quercus laeta Liebm. | Encino colorado |
| Fagaceae | Quercus mexicana Humb. et Bonpl. | Encino de México |
| Fagaceae | Quercus polymorpha Schltdl. et Cham. | Encino roble |
| Fagaceae | Quercus rugosa Née | Encino blanco |
| Ericaceae | Arbutus xalapensis Kunth | Madroño |
| Cupressaceae | Juniperus flaccida Schltdl. | Enebro |
En la Sierra Madre Oriental destacan los estudios florísticos realizados por García-Arévalo (2008), Valenzuela y Granados (2009), Aragón-Piña et al. (2010), De León et al. (2013), Delgado et al. (2016) y Graciano-Ávila et al. (2017), los cuales coinciden en que Pinaceae y Fagaceae son las familias más abundantes; Valencia (2004), Sánchez-González (2008), González-Elizondo et al. (2012) y Gernandt y Pérez-de la Rosa (2014) argumentaron que la abundancia de estas dos familias se explica por su amplia diversidad de especies en México, pues de Pinus se consignan 49 y de Quercus, 161. Asimismo, Zúñiga et al. (2018) documentan un alto valor de importancia para los géneros Pinus y Quercus, a lo que se debe agregar el gran interés económico de ambas familias en los ecosistemas forestales del país (Semarnat, 2014).
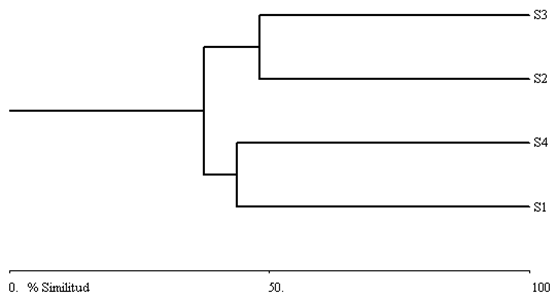
Coeficiente de similitud
El dendrograma derivado de la matriz de similitud-disimilitud de Bray-Curtis (Figura 2) agrupa los sitios de estudio en dos conjuntos bien definidos. El primero está integrado por S2 y S3, que fueron los más similares en la composición de especies, con 48.03 % de similitud y correspondieron a una comunidad vegetal de bosque mixto de encino-pino. El segundo grupo está conformado por S1 y S4, con 43.72 % de similitud, y una comunidad vegetal de pino-encino. Las agrupaciones están influidas por el número de individuos de los taxa con mayor composición florística, debido a que su abundancia resultó muy parecida, a pesar que de que su riqueza fue muy diferente entre los sitios de muestreo.
Índices de diversidad y riqueza
Los valores más altos de los índices de diversidad de Shannon-Weaver y de Simpson, y el índice de riqueza de Margalef se registraron en el S4, porque presenta el mayor número de especies; sin embargo, el valor más alto del índice de riqueza de Menhinick se determinó en el S3. Los valores de menor diversidad de Shannon-Weaver y Simpson se registraron en el S1, pero los índices de riqueza de Margalef y Menhinick en el S2, representada por cuatro especies (Figura 3).
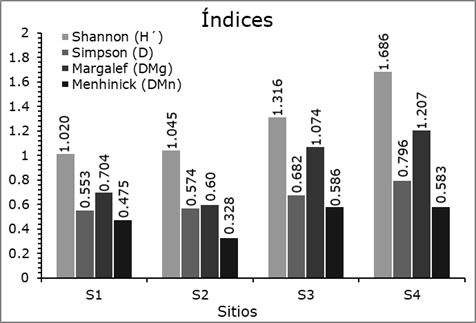
Al comparar los resultados de este estudio con los valores de diversidad y riqueza en otras regiones del país, se observa que el valor del índice de Shannon-Weaver se ubica dentro de los intervalos registrados por Solís et al. (2006) de H´=0.72 y 1.21; así como con los de Návar-Cháidez y González-Elizondo (2009) de H´=0.53 a 1.33 en bosques templados de Durango; Hernández-Salas et al. (2013) de H´=1.23 en un bosque templado de pino-encino en Chihuahua; López-Hernández et al. (2017) de H´=1.37 en bosques templados de Puebla; Domínguez et al. (2018) de H´=0.93 a 1.51 en cuatro sitios de la SMO; Buendía-Rodríguez et al. (2019) de H´=0.83 a 1.78 en 10 sitios del noreste de México.
De acuerdo al índice de Margalef, algunos autores como Hernández-Salas et al. (2013) refieren valores de D Mg =0.90 a 1.04 en bosques templados del noreste de México; López-Hernández et al. (2017) de D Mg =1.35 en bosques templados de Puebla. De igual manera para el índice de Menhinick,Návar-Cháidez y González-Elizondo (2009) y Ríos-Saucedo et al. (2019) registraron valores superiores de D Mn =0.82 y 0.95 en bosques templado de Durango, y valores de índices de riqueza de Simpson de D=0.73. Con base en los resultados de los índices de diversidad y riqueza de especies en los sitios de estudio, se clasifican como de diversidad baja a media.
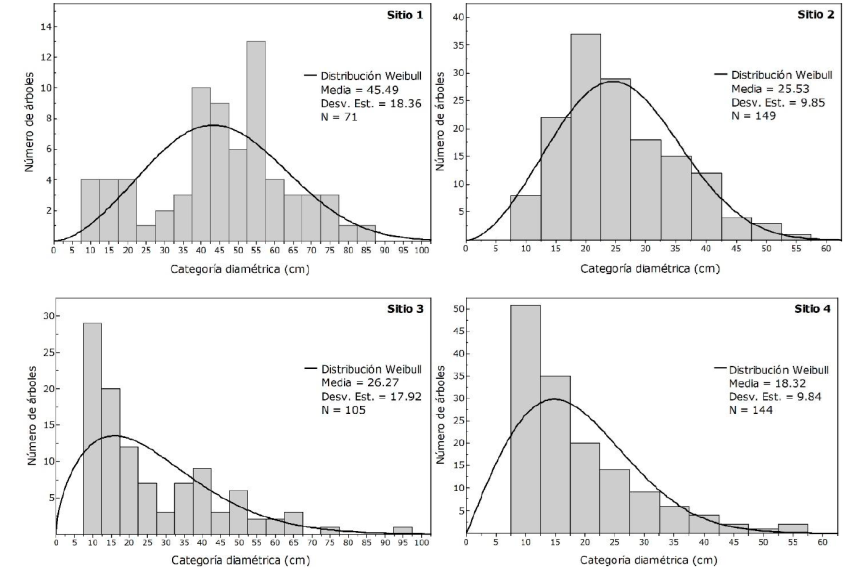
Categorías diamétricas
Las distribuciones diamétricas indican masas irregulares, pues reflejan tanto tendencias de J invertida, como curvas de distribución normal. En este estudio, se presentan bosques cuya dominancia es de árboles jóvenes con diámetros de 10 a 30 cm, valores cercanos a los registrados por Návar-Cháidez (2010) y Delgado et al. (2016) para bosques templados de Nuevo León y Durango. Se observaron individuos con diámetros mayores a 40 cm, en los cuales conforme aumenta el diámetro, la estructura de los sitios va decreciendo en densidad; lo que refleja un flujo adecuado de regeneración, con una reserva de individuos jóvenes que en un futuro podrían reemplazar a los mayores diámetros, de modo que se garantiza la supervivencia del ecosistema. Por lo tanto, es factible realizar aclareos, ya que hay individuos que han alcanzado su máximo desarrollo (Figura 4).
Estructura horizontal
Se registraron 469 árboles. A S4 le correspondió el mayor número de familias, con cuatro; seguido de S3, con tres; S1 y S2, con dos. En S3 se identificaron seis especies y en S4 siete, que son las cifras más altas; quizás por su exposición noreste, ya que propicia condiciones de temperatura y humedad favorables, pues los bosques con esta exposición presentan menor déficit hídrico debido a la baja temperatura y evapotranspiración durante el día (López-Gómez et al., 2012). S1 y S2 siguen una tendencia inferior, con cuatro familias y una exposición suroeste. En todos los sitios, al menos hubo un taxon del género Pinus y Quercus. Las especies más escasas, una por cada sitio, fueron Abies vejarii Martínez, Juniperus flaccida Schltdl., Quercus canbyi Trel. y Quercus laeta Liebm.
Q. mexicana Humb. et Bonpl fue la mejor representada en el S2, con 89 árboles (59.73 % del total); seguido de S1, con 44 árboles de P. teocote Schiede ex Schltdl. et Cham (61.97 % del total), P. pseudostrobus Lindl., con 37 individuos (35.24 % del total) en el S3; Q. canbyi con 35 árboles (24.3 % del total) en el S4. En menor proporción Q. mexicana y Q. rugosa Née con tres ejemplares (4.23 y 2.08 % del total) en S1 y S4. El sitio con el número más alto de individuos fue S2 (149), seguido de S4 (144), S3 (105) y S1 (71). Las mayores coberturas correspondieron al S2, S4 y S3 con valores de 2 853 m2 ha-1, 2 806 m2 ha-1 y 2 779.4 m2 ha-1, respectivamente; mientras que, el sitio de menor cobertura correspondió al S1 con 2 215.02 m2 ha-1. Las especies que tuvieron la mayor cobertura fueron Q. mexicana, con 3 189.44 m2 ha-1 y P. teocote, con 3 038.56 m2 ha-1 y la de menor cobertura, J. flaccida con 77.14 m2 ha-1.
Las especies más abundantes fueron Q. mexicana, P. teocote y P. pseudostrobus; en tanto que la menor se registró para J. flaccida. Las especies dominantes fueron Q. mexicana, P. pseudostrobus y P. teocote; J. flaccida tuvo el valor más bajo; y P. teocote fue la más frecuente (Cuadro 3).
Cuadro 3 Parámetros estructurales de especies por sitio de muestreo.
| Sitio | Especie | No. Ind |
Densidad | Frecuencia | Área de copa | Área Basal | IVI | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| (N ha-1) | Dr (%) | F | Fr (%) | (m2ha-1) | ACr (%) | (m2 ha-1) | ABr (%) | (%) | |||
| 1 | P. pseudostrobus | 9 | 36 | 12.68 | 0.8 | 26.67 | 369.2 | 16.67 | 10.97 | 20.47 | 18.67 |
| P. teocote | 44 | 176 | 61.97 | 0.8 | 26.67 | 1 401.56 | 63.28 | 37.91 | 70.75 | 50.64 | |
| Q. mexicana | 3 | 12 | 4.23 | 0.6 | 20 | 82.4 | 3.72 | 1.30 | 2.42 | 9.32 | |
| Q. rugosa | 15 | 60 | 21.13 | 0.8 | 26.67 | 361.86 | 16.34 | 3.41 | 6.36 | 21.38 | |
| Total | 71 | 284 | 100 | 3 | 100 | 2 215.02 | 100 | 53.59 | 100 | 100 | |
| 2 | A. vejarii | 28 | 112 | 18.79 | 0.2 | 9.09 | 525.02 | 18.40 | 4.43 | 12.63 | 15.43 |
| P. teocote | 27 | 108 | 18.12 | 0.8 | 36.36 | 559.60 | 19.61 | 11.38 | 32.49 | 24.70 | |
| Q. mexicana | 89 | 356 | 59.73 | 0.6 | 27.27 | 1 678.62 | 58.83 | 18.59 | 53.05 | 48.61 | |
| Q. polymorpha | 5 | 20 | 3.36 | 0.6 | 27.27 | 89.96 | 3.15 | 0.64 | 1.83 | 11.26 | |
| Total | 149 | 596 | 100 | 2.2 | 100 | 2 853 | 100 | 35.04 | 100 | 100 | |
| 3 | Q. mexicana | 43 | 172 | 40.95 | 0.6 | 15 | 1 428.42 | 51.39 | 21.53 | 64.74 | 35.78 |
| Q. polymorpha | 2 | 8 | 1.90 | 0.6 | 15 | 76.20 | 2.74 | 0.66 | 1.99 | 6.55 | |
| P. pseudostrobus | 37 | 148 | 35.24 | 0.8 | 20 | 771.60 | 27.76 | 6.77 | 20.34 | 27.67 | |
| Q. rugosa | 5 | 20 | 4.76 | 0.8 | 20 | 166.66 | 6 | 1.78 | 5.36 | 10.25 | |
| P. teocote | 16 | 64 | 15.24 | 0.8 | 20 | 303.6 | 10.92 | 2.48 | 7.44 | 15.39 | |
| A. xalapensis | 2 | 8 | 1.90 | 0.4 | 10 | 32.92 | 1.18 | 0.04 | 0.13 | 4.36 | |
| Total | 105 | 420 | 100 | 4 | 100 | 2 779.4 | 100 | 33.26 | 100 | 100 | |
| 4 | A. xalapensis | 30 | 120 | 20.8 | 0.4 | 11.76 | 515.20 | 18.36 | 1.81 | 9.2 | 16.99 |
| J. flaccida | 4 | 16 | 2.78 | 0.2 | 5.88 | 77.14 | 2.75 | 0.23 | 1.2 | 3.80 | |
| P. pseudostrobus | 9 | 36 | 6.25 | 0.8 | 23.53 | 189.66 | 6.76 | 1.48 | 7.6 | 12.18 | |
| P. teocote | 35 | 140 | 24.3 | 0.8 | 23.53 | 773.80 | 27.58 | 8.18 | 42 | 25.14 | |
| Q. canbyi | 35 | 140 | 24.3 | 0.2 | 5.88 | 590.24 | 21.03 | 2.51 | 13 | 17.07 | |
| Q. laeta | 28 | 112 | 19.4 | 0.2 | 5.88 | 589 | 20.99 | 4.87 | 25 | 15.44 | |
| Q. rugosa | 3 | 12 | 2.08 | 0.8 | 23.53 | 71.10 | 2.53 | 0.47 | 2.4 | 9.38 | |
| Total | 144 | 576 | 100 | 0.4 | 100 | 2 806 | 100 | 19.55 | 100 | 100 | |
No. Ind = Número de individuos; Dr = Densidad relativa; Fr = Frecuencia relativa; ACr = Área de Copa relativa; ABr = Área Basal; IVI = Índice de Valor de Importancia (Dr+Fr+ABr) /3.
Se calculó un mayor valor de importancia (IVI) para Quercus en los sitios S2, S3 y S4; en el caso de Pinus en el S1. Los taxa de mayor importancia de los sitios muestreados fueron P. teocote, P. pseudostrobus, Q. rugosa y Q. mexicana; comportamiento esperado, porque son bosques mixtos. Guzmán (2009) señaló que cuando un taxón alcanza el valor de importancia destacado, desempeña una clara dominancia ecológica; y es común que unas cuantas especies, con valores de importancia más altos, representen las poblaciones más significativas de los ecosistemas.
Por el contrario, cuando la competencia en el sitio está repartida en varias especies, ninguna de ellas supera 50 % del valor de importancia. En general, los sitios muestreados se localizan en áreas bajo manejo forestal que no han sido intervenidos hasta el momento, y que incluyen especies de interés económico; por lo que, si se les aplican los tratamientos silvícolas correspondientes, se favorecería su densidad, frecuencia, cobertura y área basal. No obstante, las dominancias genéricas hacen que en un sitio se observen bosques de pino-encino o encino-pino, lo que concuerda con lo documentado en el Ordenamiento Ecológico del estado de Durango para los bosques templados de la zona (Semarnat, 2007).
El área basal alcanzó el valor más alto en el S1 con 53.59 m2·ha-1, lo que puede explicarse por los diámetros superiores de P. teocote, y en menor proporción, en el S4 con 19.55 m2·ha-1, integrado de igual manera en su mayoría por P. teocote (Cuadro 3).
El estudio comparativo entre sitios permitió detectar variaciones estructurales y de composición que ocurren dentro de un mismo ecosistema. Aunque en términos generales, una misma especie de conífera o de latifoliada puede desarrollarse en diversos grados de pendiente u orientación, otras tienen preferencias por topoformas en particular (cerros, cañón, valle, loma, etc.) (Martínez-Antúnez et al., 2013).
Estructura vertical (A de Pretzsch)
La estructura vertical de los sitios muestreados se distribuye en tres estratos, de los cuales el superior (I: > 80 % de la altura máxima) fue dominado por P. teocote, mismo que está presente en S1, S2 y S4 con las alturas más destacadas de 26 a 30 m, y para el S3 fue P. pseudostrobus con 22.93 m. Las alturas más bajas en el S1 correspondieron a A. vejarii; en el S2 y S3, a Q. mexicana y Q. rugosa en el S3. Los diámetros en este nivel variaron de 41.13 a 94.67 m. En el estrato II (> 50 %-80 %) domina P. teocote en los cuatro sitios de muestreo, con alturas de 17.90 a 24 m. Sin embargo, otras especies comienzan a aparecer como P. pseudostrobus en S1, S3 y S4; A. vejarii en el S2 y algunas especies de Quercus: Q. cambyi y Q. laeta en el S4, Q. mexicana en S1, S2 y S3; Q. polymorpha Schltdl. et Cham en S2 y S3; y Q. rugosa en S1 y S3. Cabe señalar que Quercus es un género tolerante a la sombra. Los diámetros en este nivel fueron de 31.07 a 66 m. El estrato III (hasta 50 %), del S4 presentó la diversidad más alta de especies, con los valores más grandes de altura P. teocote, pero Q. rugosa y Q. mexicana registraron los mayores diámetros en el S1 y S3 (Cuadro 4).
Cuadro 4 Valores del índice de distribución vertical de Pretzsch.
| Sitio | Estrato | Especie | No. Ind. | D 1.30 (cm) | H (m) | Pretzsch | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min |
|
SD | Max | Min |
|
SD | A | A max | Arel% | ||||
| 1 | I | P. pseudostrobus | 7 | 74.87 | 55.30 | 65.37 | 7.83 | 29.30 | 25.91 | 27.88 | 1.42 | 1.02 | 2.48 | 41.07 |
| P. teocote | 24 | 84.60 | 42.43 | 57.68 | 11.43 | 30.00 | 24.60 | 26.74 | 1.85 | |||||
| II | P. pseudostrobus | 2 | 55.43 | 39.53 | 47.48 | 11.24 | 23.60 | 23.00 | 23.30 | 0.42 | ||||
| P. teocote | 18 | 59.47 | 31.17 | 44.84 | 7.74 | 24.00 | 16.00 | 22.07 | 1.84 | |||||
| Q. mexicana | 1 | 45.17 | 45.17 | 45.17 | - | 18.00 | 18.00 | 18.00 | - | |||||
| Q. rugosa | 2 | 59.67 | 32.80 | 46.23 | 19.00 | 19.00 | 17.80 | 18.40 | 0.85 | |||||
| III | P. teocote | 2 | 18.37 | 16.03 | 17.20 | 1.65 | 13.80 | 10.10 | 11.95 | 2.62 | ||||
| Q. mexicana | 2 | 40.53 | 21.10 | 30.82 | 13.74 | 11.00 | 9.00 | 10.00 | 1.41 | |||||
| Q. rugosa | 13 | 41.73 | 7.67 | 19.41 | 10.43 | 15.00 | 5.00 | 8.73 | 3.30 | |||||
| Total | 71 | 84.60 | 7.67 | 45.50 | 18.49 | 30 | 5 | 20.95 | 7.36 | |||||
| 2 | I | A. vejarii | 3 | 41.13 | 37.20 | 38.57 | 2.22 | 23.90 | 22.69 | 23.11 | 0.68 | 1.05 | 2.48 | 42.07 |
| P. teocote | 14 | 56.93 | 34.23 | 44.01 | 5.97 | 26.92 | 21.67 | 23.83 | 1.57 | |||||
| Q. mexicana | 11 | 44.57 | 33.93 | 38.07 | 3.42 | 25.74 | 21.59 | 22.78 | 1.27 | |||||
| II | A. vejarii | 20 | 31.07 | 14.00 | 20.96 | 4.08 | 19.91 | 15.10 | 17.04 | 1.37 | ||||
| P. teocote | 13 | 35.07 | 17.57 | 25.29 | 5.31 | 21.19 | 16.10 | 18.53 | 1.58 | |||||
| Q. mexicana | 58 | 42.33 | 14.00 | 24.94 | 5.78 | 21.14 | 13.60 | 16.74 | 2.21 | |||||
| Q. polymorpha | 2 | 32.77 | 17.80 | 25.28 | 10.58 | 17.54 | 13.92 | 15.73 | 2.56 | |||||
| III | A. vejarii | 5 | 14.70 | 7.67 | 9.83 | 2.79 | 12.00 | 6.59 | 8.84 | 2.00 | ||||
| Q. mexicana | 20 | 20.07 | 11.83 | 15.81 | 2.56 | 13.24 | 7.16 | 10.84 | 1.92 | |||||
| Q. polymorpha | 3 | 17.60 | 12.10 | 14.54 | 2.80 | 12.10 | 10.39 | 10.99 | 0.96 | |||||
| Total | 149 | 56.93 | 7.67 | 25.53 | 9.85 | 26.92 | 6.59 | 17 | 4.38 | |||||
| 3 | I | P. pseudostrobus | 5 | 61.27 | 45.67 | 53.85 | 7.43 | 22.93 | 21.50 | 21.99 | 0.57 | 1.32 | 2.89 | 45.52 |
| Q. mexicana | 2 | 94.67 | 76.83 | 85.75 | 12.61 | 21.56 | 18.64 | 20.10 | 2.06 | |||||
| Q. rugosa | 1 | 43.33 | 43.33 | 43.33 | - | 18.80 | 18.80 | 18.80 | - | |||||
| II | P. pseudostrobus | 7 | 33.87 | 15.70 | 20.69 | 6.57 | 14.60 | 11.72 | 13.38 | 0.95 | ||||
| P. teocote | 5 | 42.43 | 17.20 | 32.71 | 13.19 | 17.90 | 12.60 | 15.56 | 2.44 | |||||
| Q. mexicana | 30 | 66.00 | 18.73 | 38.49 | 13.74 | 17.82 | 11.62 | 14.97 | 2.05 | |||||
| Q. polymorpha | 2 | 38.80 | 24.50 | 31.65 | 10.11 | 15.30 | 14.43 | 14.87 | 0.62 | |||||
| Q. rugosa | 4 | 39.47 | 17.83 | 29.25 | 11.13 | 15.80 | 11.50 | 13.68 | 1.76 | |||||
| III | A. xalapensis | 2 | 8.73 | 7.93 | 8.33 | 0.57 | 6.69 | 4.00 | 5.35 | 1.90 | ||||
| P. pseudostrobus | 25 | 17.87 | 9.07 | 11.75 | 2.20 | 10.75 | 5.63 | 8.58 | 1.68 | |||||
| P. teocote | 11 | 17.97 | 7.97 | 12.66 | 2.72 | 10.60 | 5.00 | 9.06 | 1.73 | |||||
| Q. mexicana | 11 | 40.17 | 7.77 | 16.50 | 8.78 | 10.53 | 4.47 | 7.07 | 2.06 | |||||
| Total | 105 | 94.67 | 7.77 | 26.27 | 17.92 | 22.93 | 4 | 12.16 | 4.97 | |||||
| 4 | I | P. teocote | 7 | 56.10 | 27.67 | 38.90 | 9.22 | 26.60 | 21.50 | 23.57 | 1.90 | 1.69 | 3.04 | 55.39 |
| II | P. pseudostrobus | 2 | 44.40 | 33.07 | 38.73 | 8.01 | 18.73 | 15.89 | 17.31 | 2.01 | ||||
| P. teocote | 22 | 42.33 | 9.83 | 24.06 | 7.59 | 20.80 | 14.60 | 18.04 | 1.98 | |||||
| Q. cambyi | 6 | 31.30 | 15.33 | 22.96 | 5.59 | 14.80 | 13.60 | 14.08 | 0.40 | |||||
| Q. laeta | 9 | 55.80 | 19.28 | 33.26 | 12.15 | 17.62 | 14.50 | 16.45 | 1.13 | |||||
| III | A. xalapensis | 30 | 23.33 | 7.67 | 13.20 | 4.28 | 9.60 | 3.10 | 6.59 | 1.89 | ||||
| J. flaccida | 4 | 16.00 | 10.33 | 13.43 | 2.82 | 11.30 | 7.60 | 9.77 | 1.58 | |||||
| P. pseudostrobus | 7 | 23.33 | 11.57 | 14.80 | 4.21 | 12.60 | 10.40 | 11.61 | 0.70 | |||||
| P. teocote | 6 | 14.83 | 10.97 | 12.66 | 1.69 | 13.20 | 11.10 | 12.08 | 0.81 | |||||
| Q. cambyi | 29 | 23.37 | 7.73 | 12.23 | 3.47 | 12.80 | 7.00 | 10.06 | 1.40 | |||||
| Q. laeta | 19 | 23.43 | 9.03 | 14.49 | 4.48 | 12.00 | 4.12 | 8.39 | 2.25 | |||||
| Q. rugosa | 3 | 30.00 | 12.53 | 21.08 | 8.74 | 11.60 | 7.20 | 9.63 | 2.24 | |||||
| Total | 144 | 56.10 | 7.67 | 18.32 | 9.84 | 26.60 | 3.10 | 11.80 | 5.04 | |||||
No. Ind = Número de individuos; Max= Valor máximo; Min= Valor
mínimo; D
130
= Diámetro normal; H = Altura;
El índice vertical estimado fue de A=1.02 a 1.69 y un A max =2.48 a 3.04 en los sitios de muestreo, el más alto se ubicó en S4 con un A rel =55.39 %; lo que indica que el sitio evaluado tiene uniformidad media en la diversidad de alturas (Cuadro 4). Estos resultados son comparables con los documentados por Buendía-Rodríguez et al. (2019) de A=1.5 a 2.53, con A max =2.48 a 3.5 en 10 sitios de clima templado del noreste de México.
Conclusiones
Los resultados revelan que los sitios muestreados en este estudio son heterogéneos y de baja similitud. Las especies con mayor valor de importancia ecológica son P. teocote, P. pseudostrobus y Q. mexicana, de las cuales la primera ocupa un área basal superior en S1; Q. mexicana registra el número más alto de árboles en S2 y S3, y la mayor diversidad de especies en S4. No obstante, los valores de los índices de diversidad (Shannon, Simpson) y riqueza (Margalef, Menhinick) calculados son relativamente bajos en comparación con los determinados en bosques de clima templado en México. En función de la distribución diamétrica, se identifican algunos de los árboles de mayor dimensión dentro de las áreas de estudio, lo que es de interés para los manejadores del bosque al momento de hacer remociones del arbolado que ha alcanzado su madurez, sin perjudicar la estabilidad del ecosistema. Mediante la estratificación de las alturas, a través del índice de Pretzsch, se proporciona un panorama detallado de la estructura vertical de las especies que conforman los sitios de muestreo. Los resultados obtenidos, así como los datos descriptivos de este trabajo son útiles como referencia de los cambios que experimentan los bosques en estas áreas y a través del tiempo aplicar el manejo forestal correspondiente.

