











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El clima del mundo está cambiando a un ritmo sin precedentes y continuará en las siguientes décadas más rápido e intenso que lo predicho, y la influencia antrópica en este proceso es clara (IPCC, 2014). Existen numerosas evidencias de que el cambio climático tiene consecuencias ecológicas (Parmesan, 2006; Visser, 2008). La predicción de la respuesta de la diversidad biológica al cambio climático se ha convertido en un campo muy activo de la investigación científica (Beaumont et al., 2011; McMahon et al., 2011); esta respuesta juega un papel importante en alertar a científicos y tomadores de decisiones de los posibles riesgos futuros, proporciona un medio para reforzar la atribución de las modificaciones en el ecosistema y contribuyen al desarrollo de estrategias proactivas para reducir los impactos (Parmesan et al., 2011).
Se prevé que los múltiples componentes del cambio climático afectarán a todos los niveles de la biodiversidad, desde los organismos hasta las áreas bióticas (Parmesan, 2006); en los niveles más básicos, es capaz de disminuir la diversidad genética de las poblaciones debido a la selección direccional y a una rápida migración; lo que podría, a su vez, afectar el funcionamiento y la resiliencia del ecosistema (Lasco et al., 2008; Schneider et al., 2009), que conlleva a transformar la red de interacciones a nivel comunitario (Bellard et al., 2012).
Las modificaciones en el clima afectan la distribución de las especies, quienes pueden mostrar contracciones importantes en su hábitat disponible y alcanzable, e incluso algunas se extinguen (Parmesan, 2006). Ciertos modelos climáticos han demostrado ser particularmente útiles para comprender el alcance de la distribución y el impacto del cambio climático en taxones amenazados. MaxEnt es uno de ellos, tiene el potencial para predecir la pérdida de la biodiversidad en escenarios futuros (Bertrand et al., 2012), también ha mejorado significativamente la predicción de la distribución y hábitat adecuado de las especies afectadas por el cambio climático (Hu et al., 2015; Carvalho et al., 2017).
México es uno de los países con mayor diversidad biológica, por su gran complejidad fisiográfica y por su intrincada historia tanto geológica, como climática. Se estima que 12 % de la biodiversidad mundial se localiza en el territorio nacional, de esta 12 % de los géneros y de 50 - 60 % de las especies de plantas son endémicas (Sarukhán et al., 2009). Esa riqueza comprende taxa con uso potencial o actual para la agricultura y la ganadería, como es el caso de Leucaena leucocephala (Lam.) de Wit, taxon que se distribuye en los trópicos, por todo México y América Central; se produce de forma natural en la Península de Yucatán y en el Istmo de Tehuantepec, en el sur de México (Cook et al., 2005, Loya et al., 2014). Se ha extendido en una amplia gama de sitios, un tanto libres de heladas, también está presente en muchas zonas semiáridas, siempre que exista algo de humedad en el suelo (Heuzé y Tran, 2015). Posee una alta calidad nutricional de alimento para los bovinos, los ovinos y los caprinos, su biomasa aérea es rica en nutrientes, es tolerante a la sequía, además de fijar nitrógeno (N) atmosférico al suelo y controlar la erosión; su sombra atenúa la intensidad lumínica y reduce el efecto de las altas temperaturas tropicales, además favorece la vida silvestre, incrementándose así la biodiversidad (Bacab et al., 2013); se usa como alimento y medicina, su madera sirve para la construcción y para elaborar utensilios; las hojas, flores y semillas se comen frescas, crudas o cocinadas (Zárate, 1999; Heuzé y Tran, 2015).
Con el cambio climático, la diversidad de climas de México se puede alterar (Lasco et al., 2008; Schneider et al., 2009), al igual que la riqueza y distribución de especies, entre ellas la de interés para el presente estudio. Por ello, es necesario evaluar, entre otros aspectos, el efecto que tendrá el cambio climático sobre las áreas con aptitud ambiental para el desarrollo de los taxones, y con dicha información proyectar posibles medidas de adaptación o mitigación encaminadas a minimizar el impacto del cambio climático sobre los recursos naturales. Por tal motivo, los objetivos del presente estudio fueron obtener los descriptores ecológicos de L. leucocephala y estimar las alteraciones potenciales del cambio climático en las áreas con aptitud ambiental para su distribución en los periodos 2041-2060 y 2061-2080 en México.
Materiales y Métodos
Bases de datos biológicos
Se consideraron los datos de 1 138 sitios de muestreo georreferenciados, correspondientes a la base de datos del Inventario Nacional Forestal y Suelos (INFyS) de la Comisión Nacional Forestal (Conafor) (Conafor, 2016).
Bases de datos y sistema de información geográfica
Se utilizaron imágenes ráster de precipitación y temperatura de los periodos 1960-1990 (climatología de referencia), 2041-2060 y 2061-2080, los cuales se obtuvieron del portal de Global Climate Data de WorldClim (2017); y se trabajaron mediante imágenes ráster y ascii, con una resolución de 30 segundos de arco, además se utilizó el ráster de textura de suelos del Instituto Nacional de Estadística y Geografía (Inegi) (Inegi, 2009); para las climatologías 2041-2060 y 2061-2080 se consideró el MCG: GFDL-CM3, bajo los rcp 4.5 y rcp 8.5 que forma parte de la modelación del clima desarrollado por el proyecto de intercomparación de modelos acoplados Fase 5 (CMIP5) (Walton et al., 2013). Los rcp se refieren a la radiación global de energía expresada en W/m2 y asumen que esta varía al incrementarse los gases efecto invernadero (GEI).
A partir de las variables climáticas básicas antes mencionadas se derivó una serie de variables adicionales para realizar el análisis MaxEnt de nichos potenciales de distribución, bajo condiciones de la climatología 1960-1990, como para las climatologías 2041-2060 y 2061-2080. Las variables utilizadas fueron: temperatura máxima media anual, temperatura máxima de mayo a octubre, temperatura máxima de noviembre a abril, temperatura media anual, temperatura media de mayo a octubre, temperatura media de noviembre a abril, temperatura media del mes más cálido, temperatura media del mes más frío, temperatura mínima media anual, temperatura mínima de mayo a octubre, temperatura mínima de noviembre a abril, precipitación acumulada anual, precipitación acumulada de mayo a octubre, precipitación acumulada de noviembre a abril, precipitación del mes más seco, precipitación del mes más húmedo, oscilación térmica anual, fotoperiodo promedio de mayo a octubre y textura del suelo (tex).
La implementación de esos parámetros y la generación de sus respectivas imágenes ráster, se hizo con el sistema Idrisi Selva (Eastman, 2012). Las variables empleadas son las que, normalmente, se han utilizado para la potencialidad territorial de especies vegetales (Costa et al., 2010; Moreno et al., 2011).
Modelo de nicho ecológico
Se aplicó el enfoque de Máxima Entropía (MaxEnt) para modelar el nicho ecológico y predecir la distribución actual y futura del taxon, en su versión de escritorio 3.4.1 (Steven et al., 2017). El modelo MaxEnt establece predicciones de distribución de especies a partir de información de los datos de presencia (Phillips et al., 2006) y con base en las condiciones ambientales más significativas (Moreno et al., 2011). Los píxeles donde se tiene certeza de que se encuentra la especie en estudio constituyen los puntos de muestreo y las características son las variables climáticas, edáficas y otras funciones de las mismas. La finalidad del modelo es predecir la idoneidad del medio ambiente para los taxa en función de las variables ambientales (Phillips et al., 2006).
A partir de las coberturas de los parámetros y los datos de presencia de la especie, se modelaron los nichos de distribución para los períodos 1960-1990, 2041-2060 y 2061-2080; para ello se usaron 75 % de los registros de presencia como puntos de entrenamiento y 25 % como puntos de validación, 50 réplicas y 500 iteraciones (Sánchez et al., 2018). Se utilizó el índice AUC (área bajo la curva) para evaluar el modelo estadístico, ya que este índice es de los más utilizados para medir la calidad de los modelos (Moreno et al., 2011). La complejidad del modelo se controla con la elección de las variables; además, MaxEnt presenta, dentro de su configuración, un operador denominado “regularización de parámetros” que permite evitar la sobreposición de variables, lo que evita su efecto perjudicial sobre la predicción (Phillips y Dudík, 2008).
Los valores de los puntos obtenidos representan valores de idoneidad relativa para la presencia de las especies, y constituyen la base de un modelo de distribución potencial. Las imágenes resultantes de la modelación MaxEnt de nichos de distribución se recuperaron en el sistema ArcGis (ESRI, 2010) y se editaron en el formato de mapas.
Ajuste del modelo
Maxent predice por medio del modelo de nicho ecológico la tasa de ocurrencia relativa (curva ROC) de la especie, como una función de los predictores ambientales en cada localidad (Merow et al., 2013); a su vez, el área bajo la curva (AUC) se interpreta como el ajuste del modelo, en el que un valor de 1.0 sería un clasificador perfecto y uno aleatorio tendría un valor de 0.5 (Phillips et al., 2006), por lo que aquéllos cercanos a 1.0 muestran mayor ajuste del modelo a los datos.
Los resultados de Maxent incluyen imágenes con áreas de distribución potencial de la especie, donde cada celda tiene un valor de predicción; por otra parte, en un gran número de investigaciones se requiere fijar un valor umbral para estimar las áreas con aptitud ambiental para cada taxón (Liu et al., 2016), para lo cual el modelo Maxent integra un archivo con los diferentes valores umbrales usados con mayor frecuencia. En el presente estudio se utilizó como umbral logístico el valor 10 acumulativo fijo (Fixed cumulative value 10).
Resultados y Discusión
Modelo del nicho de distribución potencial
Los resultados de la aplicación de la técnica Receiver Operating Characteristic (ROC) indicaron que el modelo es adecuado, ya que el valor del área bajo la curva del escenario actual fue de 0.948 para los datos de entrenamiento (línea roja), y para los escenarios futuros fue superior a 0.95 (Figura 1). Lo que indica que el modelo para clasificar la presencia de L. leucocephala fue satisfactorio (Parolo et al., 2008).

Por lo general, se considera que un valor de AUC mayor a 0.7 tiene un buen ajuste (Hanley y McNeil, 1982; Elith, 2006). Las variables ambientales que contribuyeron en 74 % a explicar la distribución de la especie fueron: el fotoperiodo, la precipitación del mes más húmedo, temperatura media del mes más frío, precipitación acumulada de mayo a octubre y la temperatura media de noviembre a abril.
Descriptores ecológicos
En el Cuadro 1, se resumen los parámetros de los descriptores ecológicos de las variables ambientales que, en mayor medida, contribuyeron a explicar la distribución de L. leucocephala. Se observa que los intervalos ambientales de distribución resultaron amplios, tanto para las variables relacionadas con la temperatura como para las de precipitación. Este hecho evidencia una amplia rusticidad de la especie, característica que podría ser favorable en ambientes donde el cambio climático se manifieste de manera más agresiva (Heuzé y Tran, 2015).
Cuadro 1 Descriptores ecológicos de las variables ambientales que contribuyen en 74 % a la distribución geográfica de Leucaena leucocephala (Lam.) de Wit en México.
| Variables | Leucaena leucocephala | ||
|---|---|---|---|
| Mín | Máx. | % C | |
| Fotoperiodo noviembre-abril (h) | 11.2 | 11.6 | 28.8 |
| Precipitación del mes más húmedo (mm) | 60 | 640 | 24.8 |
| Temperatura media del mes más frío (°C) | 11 | 28 | 11.1 |
| Precipitación acumulada de mayo a octubre (mm) | 297 | 3 157 | 4.7 |
| Temperatura media de noviembre a abril (°C) | 14.2 | 28.5 | 4.4 |
Mín = Valor mínimo; Máx = Valor máximo; % C = Porcentaje de contribución.
Con base en la información del Cuadro 1, L. leucocephala se distribuye en regiones donde el fotoperiodo de la temporada noviembre-abril se ubica entre 11.2 y 11.6 horas; condición que coincide con lo consignado por Heuzé y Tran (2015), quien señala que es una especie de día corto, cuya floración y reproducción se estimula con un fotoperiodo inferior a 12 horas. Sin embargo, Whitesell y Parrotta (2008) indica al respecto que las variedades comunes de L. leucocephala florecen todo el año. Otra variable de importancia es la temperatura media del mes más frío, misma que se distribuye en sitios con un valor de 11 a 28 °C, condición que corresponde a zonas desde templadas a muy cálidas (Medina et al., 1998); por lo que, la temperatura parece no ser un factor tan restrictivo para su distribución.
Otra variable importante fue la precipitación del mes más húmedo; en el Cuadro 1 se muestra que L. leucocephala se desarrolla en lugares con valores entre 60 y 640 mm, los cuales existen en zonas de semiáridas a húmedas y que corresponden a valores de humedad relativa de bajos a altos en el mes más húmedo del año. Lo anterior coincide con lo señalado por Petit et al. (2010), quienes refieren que L. leucocephala se distribuye en forma natural en áreas donde la humedad relativa en el mes de septiembre (por lo general, el mes más húmedo) se registra hasta con 85 %.
Áreas con aptitud ambiental de Leucaena leucocephala
La aptitud ambiental durante el periodo 1960-1990 (Figura 2), se apreció favorable para la especie bajo estudio como resultado de sus amplios intervalos ambientales (Heuzé y Tran, 2015). La superficie con aptitud ambiental fue notoria en gran parte del centro y sur de México, lo que demuestra que las condiciones tropicales y subtropicales corresponden mejor a las necesidades climáticas de la especie (Zárate, 1999; Heuzé y Tran, 2015).
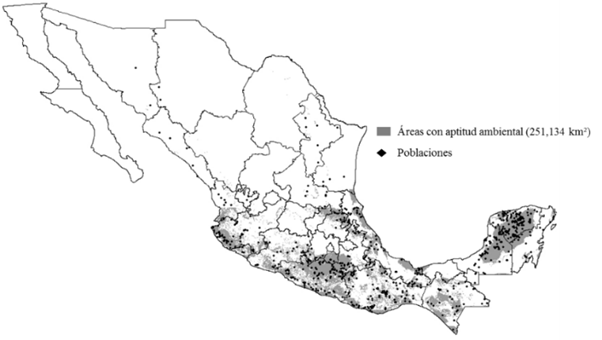
Figura 2 Áreas con aptitud ambiental para Leucaena leucocephala (Lam.) de Wit en el periodo 1960-1990.
La mayor presencia de las poblaciones se concentró en la Sierra Madre del Sur, en las llanuras costeras del Golfo de México y, en gran proporción, en la Península de Yucatán; esa presencia está en correspondencia con lo documentado, en el sentido de que L. leucocephala crece en gran variedad de ambientes, ya que posee una amplia gama de adaptación climática (Loya et al., 2014).
En la Figura 2, se observa que la superficie con aptitud ambiental abarca una extensión de 251 134 km², ubicada principalmente en las zonas costeras del Golfo de México, Sierra Madre Oriental, Sierra Madre Occidental, Sierra Madre del Sur, Sierra Norte de Puebla, Istmo de Tehuantepec y gran parte de la Península de Yucatán. Información que coincide con lo registrado por Loya et al. (2014) y Heuzé y Tran (2015), quienes señalan que esta leguminosa tiene una amplia distribución en México y se propaga en climas tropicales y subtropicales con altitudes que varían desde el nivel del mar hasta los 1 500 m. Este intervalo altitudinal de la especie podría ampliarse debido a la presión del incremento en la temperatura de los periodos estudiados, por lo que L. leucocephala migrará hacia altitudes mayores.
Aptitud ambiental para Leucaena Leucocephala en los escenarios de cambio climático
La delimitación de áreas para la especie y su dinámica territorial, en función de los cambios climáticos, determinan en qué sentido impactarán sobre el incremento, disminución o mantenimiento de las áreas de distribución.
En los mapas de la Figura 3, se muestra la dinámica territorial que tendrán las áreas con aptitud ambiental para L. leucocephala; al respecto, la predicción del MCG augura una dispersión favorable de la superficie en los dos periodos climáticos y en las dos rutas representativas de concentración de GEI (Cuadro 2); aunque se acentúa con el rcp 4.5 para ambos periodos. Referente al periodo 2041-2060, el área con aptitud ambiental se incrementará 72 %, y para el escenario 2061-2080 de 61 % (Figura 4); lo cual concuerda con lo señalado por Guitérrez y Trejo (2014) quienes registran que el efecto del cambio climático sobre los ecosistemas se reflejará en la alteración de la distribución y abundancia de las especies, o en la desaparición directa de algunos taxa y de poblaciones.
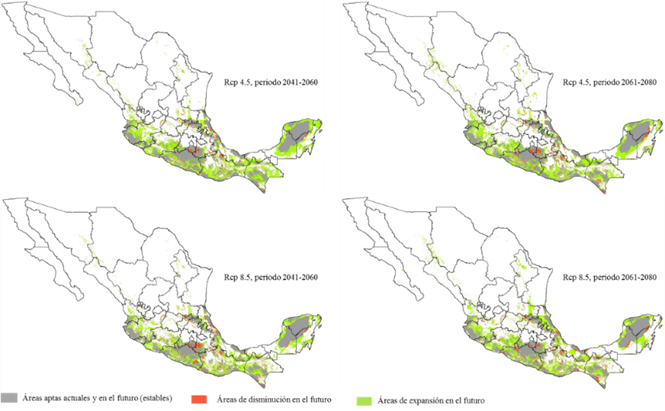
Figura 3 Áreas con aptitud ambiental para dos periodos de tiempo y dos rcp con el MCG GFDL-CM3 para Leucaena leucocephala (Lam.) de Wit.
Cuadro 2 Áreas (km²) con aptitud ambiental para Leucaena leucocephala (Lam.) de Wit con el MCG GFDL-CM3, y dos trayectorias representativas de concentración de gases efecto invernadero (rcp 4.5, rcp 8.5) en México.
| Periodos | 2041-2060 | 2061-2080 | ||
|---|---|---|---|---|
| Rcp | 4.5 | 8.5 | 4.5 | 8.5 |
| Áreas con AA | 434 173 | 380 927 | 406 115 | 383 965 |
AA = Aptitud ambiental
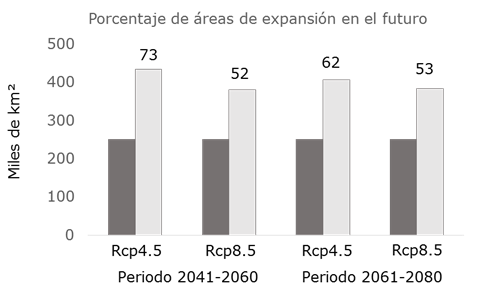
Figura 4 Porcentaje de expansión de áreas con aptitud ambiental (color gris oscuro), respecto al clima de referencia, en los periodos 2041-2060 y 2061-2080 para dos rcp (4.5 y 8.5).
En la Figura 3 se observan pequeñas nuevas áreas con aptitud ambiental para L. leucocephala; ubicadas en la Sierra Madre Occidental, específicamente en los estados de Sonora, Sinaloa, Chihuahua, Durango y Nayarit; en la Sierra Madre Oriental, en Nuevo León y Tamaulipas. También, el modelo pronostica mayores incrementos en la Sierra Madre del Sur, Istmo de Tehuantepec y en la Península de Yucatán; esto coincide con lo citado por Hu et al. (2015), quienes refieren que las tasas del cambio climático afectarán, dramáticamente, los grupos de comunidades y la biodiversidad en las zonas tropicales de la Tierra.
Aunque los rcp coinciden con la tendencia de variación de la superficie de aptitud ambiental, sí se diferencian en cuanto a la magnitud (Cuadro 2). Esto se debe, principalmente, a las variaciones de la precipitación, ya que el rcp 8.5 estima volúmenes anuales de lluvia más bajos en los dos periodos climáticos futuros.
Lo antes expuesto evidencia que las áreas con aptitud ambiental son sensibles a las variaciones de simulación climática de los rcp, incluso hasta en los modelos que se consideran similares en la predicción del cambio climático para México (Conde et al., 2006).
Asimismo, se presentan algunas posibles áreas dinámicas con aptitud ambiental que responden al cambio climático (Figura 3); mismas que aparecen en las regiones centro y norte de México, y que ya han sido registradas con cambios en los patrones de cultivo, debido al cambio climático (Ramírez et al., 2011; Santillán-Espinoza et al., 2011).
Las diferencias en las proyecciones de la superficie con aptitud ambiental para L. leucocephala, en las trayectorias de concentración de GEI del modelo de circulación general utilizado, coinciden con lo referido por Beaumont et al. (2008), quienes mencionan que, independientemente del modelo que se utilice para calcular la pérdida de la biodiversidad al cambiar las trayectorias de emisiones de gases efecto invernadero, los resultados obtenidos pueden ser contrastantes.
En la Figura 3, también se perciben pequeñas áreas con aptitud ambiental que L. leucocephala pierde en los escenarios futuros, las cuales se localizan principalmente para los cuatro rcp, en los estados de Jalisco, Michoacán, Hidalgo, Querétaro, San Luis Potosí, Guerrero, Oaxaca, Chiapas, Puebla, Morelos, Veracruz, Tabasco y Península de Yucatán. Información similar a la señalada por Uribe (2015), en el sentido de que el cambio climático pronosticado para el 2050, tendrá graves impactos sobre la biodiversidad en el territorio mexicano.
Conclusiones
El cambio climático previsto para los periodos 2041-2060 y 2061-2080, con las trayectorias representativas de concentración de GEI (rcp 4.5 y rcp 8.5), incrementará la superficie con aptitud ambiental para L. leucocephala, principalmente, en regiones de la Sierra Madre Occidental, Sierra Madre Oriental y en gran parte de la Península de Yucatán. La ganancia en superficie con AA será mayor con el rcp 4.5 para los dos periodos climáticos, con un promedio de 67 %; para el rcpr 8.5 el modelo pronostica aumentos en la superficie con AA hasta de 53 %, en promedio para ambos periodos climáticos.
Leucaena leucocephala se adapta a un amplio intervalo de condiciones de precipitación y temperatura, lo que le provee ventajas adaptativas a los ambientes con variación climática.

