











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los bosques estacionalmente secos (BES) en el Neotrópico se distribuyen desde el norte de México hasta el sur de Brasil y conforman 66.7 % de la superficie de este tipo de vegetación del mundo (Miles et al., 2006). En Perú, los BES son ecosistemas amenazados, poco estudiados y comprenden varios subtipos (Linares-Palomino, 2006; Linares-Palomino y Pennington, 2007). Los BES de montaña y de llanura de la costa norte de Perú son muy frágiles; se extienden desde la Península de Santa Elena, en el sur del Ecuador, hasta varias regiones del noroeste del Perú, entre las cuales están Tumbes, Piura, Lambayeque y el norte de La Libertad, así como el piso inferior del valle del Marañón. Las dos últimas áreas se comunican por medio del Paso de Porculla, aunque son unidades fitogeográficas distintas (Brack y Mendiola, 2004). También se le conoce como región de endemismos Tumbesina, la cual es compartida por Ecuador y Perú, y se le considera una de las zonas más importantes del mundo (Aguirre et al., 2006).
Algunos estudios han revelado el gran número de especies endémicas presentes en el BES del norte y noroeste del Perú, con alrededor de 26 % de ellas diseminadas en varias comunidades, tales como Prosopis limensis Benth. (algarrobo) y Colicodendron scabridum Seem. (zapote) (Sagástegui et al., 1999). Pero, no solo los niveles de diversidad y endemismo se refieren a plantas, sino también a vertebrados como Tremarctos ornatus Cuvier, 1825 (oso de anteojos), Lagidium peruanum Meyen, 1833 (vizcacha), Penelope albipennis Taczanowski, 1878 (pava aliblanca) y Bothrops barnetti Parker, 1994 (macanche), entre otros (Aguilar, 1994).
Los trabajos sobre el BES, aunque no son muy numerosos,4 sí han resultado significativos;. son particularmente interesantes los realizados por Weberbauer (1922; 1930; 1936; 1945), quien aportó aspectos florísticos y fitogeográficos de la flora del Perú y elaboró un mapa de vegetación, hasta ahora insuperable, con énfasis en el BES. En la década pasada destacan la lista anotada de especies leñosas y formaciones vegetales en los BES de la región de endemismo Tumbesina de Ecuador y Perú (Aguirre et al., 2006), así como de las plantas leñosas como una nueva herramienta en internet para estudios taxonómicos, ecológicos y de biodiversidad (Linares-Palomino y Pennington, 2007).
Con base en datos de inventarios forestales se analizaron y discutieron los patrones de estructura y diversidad de los BES de dos áreas de conservación de la Reserva de la Biosfera del Noroeste (Perú), el Parque Nacional Cerros de Amotape y la Zona Reservada de Tumbes (Leal-Pinedo y Linares-Palomino, 2005). Se abordaron, los patrones de distribución espacial de las especies arbóreas como Eriotheca ruizzii (K. Schum.) A. Robyns (pasayo), Bursera graveolens Triana & Planch. (palo santo), Caesalpinia glabrata Kunth (charán) y Cochlospermum vitifolium (Willd.) Spreng. (polo polo) en un BES del Parque Nacional Cerros de Amotape, en el noroeste del Perú, (Linares-Palomino, 2005). En los alrededores de Jaén (Cajamarca, Perú), en relictos del bosque tropical estacionalmente seco, se han efectuado ocho inventarios sobre la vegetación leñosa, endemismos y su estado de conservación (Marcelo-Peña, 2008).
Por otro lado, con la finalidad de diseñar mapas y llevar a cabo la clasificación de vegetación en ecosistemas estacionales de los bosques secos de Piura (Perú), con latos de inventarios florísticos, se identificaron seis grupos de vegetación; de ellos, cinco coincidieron con los tipos de vegetación descritos hace 100 años por A. Weberbauer (Weberbauer, 1922; 1930; 1936; 1945) (La Torre-Cuadros y Linares-Palomino, 2008). En la provincia de Ayabaca (Piura), se evaluaron las características edáficas y la composición florística de los BES La Menta y Timbes (Rasal et al., 2011).
En la región Lambayeque (Perú), también se han hecho algunos estudios en el BES. En la Reserva Ecológica Chaparrí, que forma parte del punto candente de biodiversidad Tumbes-Chocó-Magdalena, se elaboró un patrón estructural e inventario florístico, del que resultó evidente un bosque bastante empobrecido, en relación con otros BES de la región (Linares-Palomino y Ponce-Álvarez, 2009). En esta misma Reserva Ecológica Privada Chaparrí se determinó la fenología (desarrollo vegetativo, floración y fructificación) de 17 especies vegetales de las que se alimenta pava aliblanca, especie endémica seriamente amenazada (Martos et al., 2009).
Con base en lo anterior, además de que los bosques naturales del Perú han sido escasamente estudiados, y que cada vez es mayor la presión antrópica que conlleva a su depredación; y que el cambio climático continúa alterando los patrones de distribución vegetal, entre otros factores (Steffen et al., 2011), se llevó acabo el presente trabajo, cuyo objetivo fue determinar la composición florística del BES del Cerro Tres Puntas con el propósito de facilitar la elaboración de planes de reforestación, conservación y utilización, para con ello contribuir al conocimiento de los ecosistemas naturales del Perú.
Materiales y Métodos
Área de estudio
El área de estudio se localiza entre los 6°14'16.23" S - 79°35'34.81" O y los 6°12'46.46" S 79°33'59.58" O; comprende el Cerro Tierra Blanca (1 986 msnm), el Cerro Gato (1 545 msnm), ambos pertenecientes al distrito de Salas, y el Cerro Tres Puntas (1 590 msnm), del distrito de Motupe, en la provincia y región de Lambayeque, Perú (Figuras 1a -b); estos tres cerros abarcan una extensión aproximada de 2 975 ha.
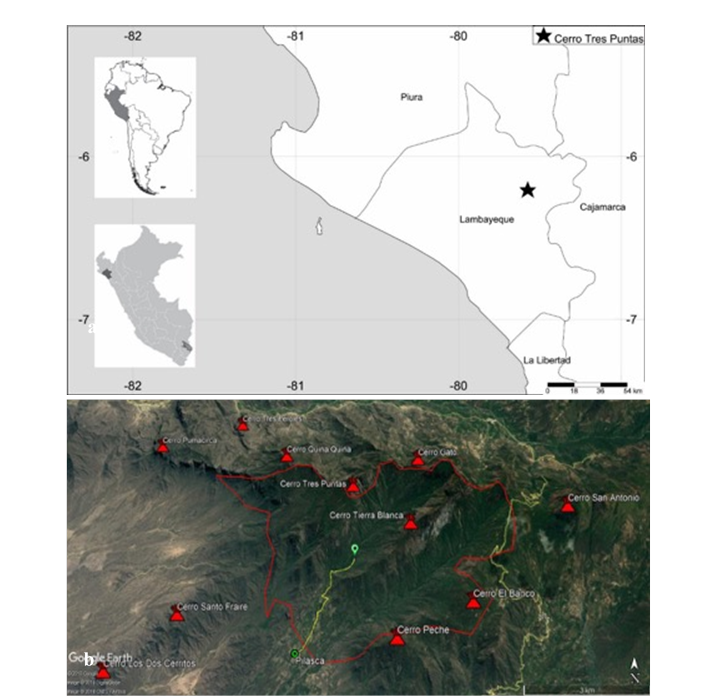
Fuente: Google Earth (2019).
Figura 1 a) Mapa de la región Lambayeque, indicando con la estrella la localidad de Pilasca, donde se ubica el Cerro Tres Puntas; b) Área que comprende los Cerros Tierra Blanca, Gato y Tres Puntas del BES (Salas-Motupe, Lambayeque, Perú).
El caserío Pilasca es uno de los más pobres de Lambayeque (Gobierno Regional de Lambayeque, 2013) en el que, la agricultura de maíz (Zea mays L.), yuca (Manihot esculenta Crantz) y frutales, así como la ganadería de vacunos y caprinos son de subsistencia; ocupa pequeños claros del bosque, donde no hay selección de semillas, fertilización, ni control de plagas y enfermedades. Los comuneros de Pilasca han utilizado las especies del bosque desde tiempos ancestrales para la fabricación de muebles, construcción de viviendas rústicas y corrales, al igual que la leña como combustible; razón por la cual Loxopterygium huasango Spruce ex Engl. (hualtaco), Handroanthus chrysanthus (Jacq.) S. O. Grose (guayacán amarillo), Colicodendron scabridum (Kunth) Seem. (zapote), Prosopis limensis Benth. (algarrobo) y Cedrela sp. (cedro), prácticamente ya no existen.
En el aspecto geológico, la zona corresponde a la unidad litoestratigráfica Grupo Salas (Oi) del Paleozoico o Formación Salas, (Pi-s) Ordovícico inferior (480 a 470 millones de años) (Gobierno Regional de Lambayeque, 2013; López, 2013).
El área en la que se hizo la investigación es un ecosistema de bosque semidenso estacionalmente seco de colina y montaña (Bes-cm) (Minam, 2019). El levantamiento de los datos se realizó en un lugar ubicado entre los 373 y los 720 msnm; el punto extremo A corresponde al ingreso del caserío Pilasca (373 m), donde se inició la evaluación, hasta el punto extremo E (720 msnm), en el que terminó, cuya topografía es agreste con enormes peñascos y quebradas profundas, secas y escasos claros pedregosos. Los datos de temperatura, humedad relativa y precipitación correspondieron a los de la Estación CO-Meteorológica Jayanca (LA Vic) (78 msnm), la más cercana al Cerro Tres Puntas de Pilasca, de la que se registraron los siguientes datos: temperatura máxima y mínima y humedad relativa de marzo 2018, 34.0 y 19.0 oC y 78.4 %, respectivamente; mientras que, en junio 2018, 26.2 y 15.0 oC y 79.1 %, respectivamente. En los cuatro meses de evaluación, la precipitación fue 0.0 mm día-1 (Senamhi, 2018).
Muestreo
El trabajo de campo se llevó a cabo de marzo a junio de 2018 y se utilizó el método propuesto por Gentry (1982; 1995a) para el muestreo de plantas leñosas, con sustanciales modificaciones, debido a la difícil y peligrosa geografía del terreno. Se aplicó un muestreo lineal, cuya línea base fue una trocha (camino estrecho o brecha) relativamente segura que los lugareños utilizan para acceder, ocasionalmente, a la cima del Cerro Tres Puntas (Figura 2).

Se establecieron cuatro unidades de muestreo en una extensión total de 3 801 m, designadas con los puntos A (373 msnm), B (456 msnm), B-C (565 msnm), C-D (677 msnm) y D-E (720 msnm), que abarcaron 958 m, 961 m, 968 m y 914 m, respectivamente. Del centro de la trocha, con un GPS Garmin 60CSX y una cinta métrica, se midieron 15 m a la derecha y 15 m a la izquierda, para un total de 30 m, lo que permitió evaluar alrededor de 114 030 m2. Se registraron todos los individuos leñosos de DAP (1.3 m de la base del tronco) ≥ 5.0 cm.
Para individuos con troncos ramificados, se midió el diámetro de cada ramificación a una altura de 1.3 m de la base del tronco, con una cinta diamétrica Forestry Suppliers, Inc. (Modelo 283D/5M). Como las especies evaluadas eran conocidas, solamente, se colectaron muestras botánicas para su registro y depósito en el Herbario de la Universidad Nacional Pedro Ruiz Gallo de Lambayeque (Herbario HPR). Asimismo, se prensaron ejemplares herbáceos, presentes en el área de colecta, para su posterior identificación taxonómica. Para las familias, se siguieron los postulados filogenéticos del Grupo de Filogenia de las Angiospermas (APG, 2009; 2016).
Procesamiento y análisis de la información
Para determinar la abundancia o densidad de los taxones muestreados se utilizó la clasificación de Maleheiros y Rotta (1982), la cual los cataloga como raros o escasos (1 - 6 individuos); poco abundantes (7 - 25); y abundantes (más de 25 individuos). La estructura horizontal del bosque se describió a partir de la distribución del número de árboles por clase diamétrica y de altura (Hawley y Smith, 1972). Los análisis fitosociológicos se hicieron por medio del freeware FITOPAC 2.1 (Shepherd, 2010). Con dicho soporte, se calcularon los siguientes parámetros: Densidad Relativa (DER), Frecuencia Relativa (FER), Área Basal Relativa (AB) o Cobertura Relativa (COB), Índice de Valor de Importancia (IVI) y Valor de Importancia para la Familia (VIF); fórmulas que fueron descritas por Rangel y Velásquez (1997).
Resultados
Composición florística
En los 114 030 m2 estudiados se inventariaron 410 individuos de árboles y arbustos con DAP≥5.0 cm que correspondieron a 17 especies, 17 géneros y 10 familias, registrados en 47 unidades de muestreo (Cuadro 1). A los taxa arbóreos y arbustivos (Cuadro 1) se sumaron numerosas herbáceas con diferentes hábitos de crecimiento como bejuco, enredadera, epífita, subarbusto, sufrútice, entre otros; colectadas con flores y frutos en mayo de 2018, entre las que pueden citarse: Salvia occidentalis Sw. (Lamiaceae), Psittacaenthus chanduyensis Eichler (Loranthaceae), Croton pavonis Müll. Arg. (Euphorbiaceae), Scutia spicata (Wild.) Weberb. (Rhamnaceae), Alternanthera pubiflora Kuntze (Amaranthaceae), Dalechampia scandens L. (Euphorbiaceae), Cordia macrocephala (Desv.) Kunth (Boraginaceae), Plumbago scandens L. (Plumbaginaceae), Heliotropium angiospermum Murray (Boraginaceae), Waltheria ovata Cav. (Malvaceae), Wedelia latifolia DC. (Asteraceae), Cardiospermum corindum L. (Sapindaceae), Tessaria integrifolia Ruiz & Pav. (Asteraceae), Pseudogynoxys cordifolia (Cass.) Cabrera (Asteraceae), Tetramerium nervosum Nees (Malvaceae), Scoparia dulcis L. (Scrophulariaceae) y Cestrum auriculatum L’Hér. (Solanaceae).
Cuadro 1 Principales características dasométricas de las especies presentes en el área evaluada del BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Familia/Especie | Nombre común | Háb. crec. | Núm. indiv./% | Unidad muestr. | DAP (cm) | Altura total (m) | Diám. copa (m) |
|---|---|---|---|---|---|---|---|
| Anacardiaceae | |||||||
| Loxopterygium huasango Spruce ex Engl. | Hualtaco | Árbol | 20/ 4.9 | 4 | 27.79 | 12.83 | 8.84 |
| Boraginaceae | |||||||
| Cordia lutea Lam. | Overo | Árbol/⁓ Arbust. | 29/ 7.1 | 3 | 1.83 | 5.97 | 6.52 |
| Burseraceae | |||||||
| Bursera graveolens (Kunth) Triana & Planch. | Palo santo | Árbol | 4/ 1.0 | 2 | 22.45 | 11.00 | 6.88 |
| Fabaceae | |||||||
| Albizia multiflora (Kunth) Barneby & J.W. Grimes | Angolo | Árbol | 8/ 2.0 | 4 | 33.39 | 14.75 | 11.43 |
| Caesalpinia glabrata Kunth | Charán | Árbol | 5/ 1.2 | 1 | 11.92 | 7.00 | 6.92 |
| Erythrina smithiana Krukoff | Frejolillo | Árbol | 32/ 7,8 | 3 | 36.03 | 11.69 | 9.14 |
| Pithecellobium excelsum (Kunth) Mart. | Chaquiro | Árbol/ ⁓ Arbust. | 21/ 5.1 | 4 | 11.04 | 7.33 | 9.64 |
| Prosopis limensis Benth. | Algarrobo | Árbol | 9/ 2.2 | 1 | 37.31 | 16.56 | 12.89 |
| Senna atomaria (L.) Irwing & Barneby | Chapa | Árbol | 15/ 3.7 | 2 | 9.83 | 5.47 | 4.23 |
| Vachellia macracantha (Willd.) Seigler & Ebinger | Faique | Árbol/⁓ Arbust. | 154/ 37.6 | 4 | 16.34 | 8.54 | 8.99 |
| Lauraceae | |||||||
| Beilschmiedia sulcata (Ruiz & Pav.) Kosterm | Palta de zorro | Árbol | 1/ 0.2 | 1 | 124.10 | 12.00 | 22.10 |
| Malvaceae | |||||||
| Eriotheca ruizii (K. Schum.) A. Robyns | Pasayo | Árbol | 6/ 1.5 | 1 | 31.3 | 10.58 | 7.37 |
| Guazuma ulmifolia Lam. | Guazumo | Árbol | 11/ 2.7 | 3 | 27.42 | 12.83 | 10.52 |
| Moraceae | |||||||
| Ficus obtusifolia Kunth | Higuerón | Árbol | 17/ 4.2 | 4 | 120.06 | 19.35 | 24.00 |
| Olacaceae | |||||||
| Ximenia americana L. | Ciruelillo | Árbol | 17/ 4.2 | 3 | 11.40 | 10.06 | 5.9 |
| Sapindaceae | |||||||
| Sapindus saponaria L. | Choloque | Árbol | 6/ 1.5 | 3 | 17.6 | 10.00 | 6.45 |
| Cannabaceae | |||||||
| Celtis iguanaea (Jacq.) Sarg. | Palo blanco | Árbol | 55/ 13.4 | 4 | 38.48 | 14.62 | 12.99 |
| Total | 410 | 47 | 26.20 | 10.45 | 9.81 | ||
Háb. crec.= Hábito de crecimiento; Núm. indiv. = Número de individuos; Unidad muestr. = Unidad de muestreo; DAP = Diámetro a la Altura del Pecho; Diám. copa = Diámetro de copa.
Estructura
De las 17 especies inventariadas, pocos ejemplares de Cordia lutea Lam., Vachellia macracantha (Willd.) Seigler & Ebinger y Pitecellobium excelsum (Kunth) Benth. mostraron hábito de crecimiento arbustivo con más de 10 fustes por individuo, en algunos casos. Del total de especies registradas, solo seis [L. huasango, V. macracantha, P. excelsum, Albizia multiflora (Kunth) Barneby & J.W. Grimes, Ficus obtusifolia Kunth y Celtis iguanaea (Jacq.) Sarg.] se registraron en las cuatro unidades de muestreo; en tanto que, Caesalpinia glabrata Kunth, P. limensis, E. ruizii y Beilschmiedia sulcata (Ruiz & Pav.) Kosterm. solamente se identificaron en una de ellas.
El mayor número de los 410 individuos inventariados correspondió a la tercera unidad de muestreo (565 - 677 msnm), con 194 (47.32 %); lo que se atribuye a lo agreste y pedregoso del territorio, donde no es posible desarrollar actividades agropecuarias; en contraste, en la segunda unidad de muestreo (456 - 565 msnm) el número de individuos fue menor con 42 (10.24 %), esto porque es el área con mayor vocación agrícola, por ello el bosque ha sido fuertemente depredado (Cuadro 2).
Cuadro 2 Distribución de individuos por unidad de muestreo de las especies presentes en el área evaluada del BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Especie | Unidades de muestreo | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |||||
| No | % | No | % | No | % | No | % | |
| Beilschmiedia sulcata (Ruiz & Pav.) Kosterm | 0 | 0.0 | 0 | 0.0 | 0 | 0.0 | 1 | 1.56 |
| Bursera graveolens Triana & Planch. | 2 | 1.82 | 0 | 0.0 | 2 | 1.03 | 0 | 0.0 |
| Caesalpinia glabrata Kunth | 0 | 0.0 | 0 | 0.0 | 5 | 2.58 | 0 | 0.0 |
| Celtis iguanaea (Jacq.) Sarg. | 9 | 8.18 | 7 | 16.67 | 23 | 11.86 | 16 | 25.00 |
| Cordia lutea Lam. | 10 | 9.10 | 2 | 4.76 | 17 | 8.76 | 0 | 0.0 |
| Erythrina smithiana Krukoff | 0 | 0.0 | 1 | 2.38 | 26 | 13.40 | 5 | 7.81 |
| Eriotheca ruizii (K. Schum.) A. Robyns | 0 | 0.0 | 0 | 0.0 | 6 | 3.09 | 0 | 0.0 |
| Ficus obtusifolia Kunth | 1 | 0.91 | 1 | 2.38 | 3 | 1.55 | 12 | 18.75 |
| Guazuma ulmifolia Lam. | 4 | 3.64 | 5 | 11.90 | 2 | 1.03 | 0 | 0.0 |
| Loxopterygium huasango Spruce ex Engl. | 2 | 1.82 | 4 | 9.52 | 12 | 6.19 | 2 | 3.13 |
| Pithecellobium excelsum (Kunth) Mart. | 11 | 10.0 | 4 | 9.52 | 5 | 2.58 | 1 | 1.56 |
| Albizia multiflora (Kunth) Barneby & J.W. Grimes | 2 | 1.82 | 2 | 4.76 | 2 | 1.03 | 2 | 3.13 |
| Prosopis limensis Benth. | 9 | 8.18 | 0 | 0,0 | 0 | 0.0 | 0 | 0.0 |
| Sapindus saponaria L. | 1 | 0.91 | 3 | 7.14 | 2 | 1.03 | 0 | 0.0 |
| Senna atomaria (L.) Irwing & Barneby | 9 | 8.18 | 6 | 14.29 | 0 | 0.0 | 0 | 0.0 |
| Vachellia macracantha (Willd.) Seigler & Ebinger | 47 | 42.73 | 7 | 16.67 | 81 | 41.75 | 19 | 29.69 |
| Ximenia americana L. | 3 | 2.73 | 0 | 0.0 | 8 | 4.12 | 6 | 9.38 |
| Total = 410 individuos | 110 | 26.83 | 42 | 10.24 | 194 | 47.32 | 64 | 15.61 |
El dosel superior se conformó por árboles dominantes de F. obtusifolia y P. limensis, aunque con individuos muy dispersos. Los ejemplares de mayor tamaño fueron F. obtusifolia con 19.35 m de altura, seguidos por P. limensis (16.56 m), P. excelsum (14.75 m) y C. iguanaea, (14.62 m) en una distribución de las alturas que no es del todo una “J” invertida, como se espera en bosques naturales y en bosques regulares.
El diámetro de copa fue mayor en F. obtusifolia (24.00 m) y B. sulcata (22.10 m) (Cuadro 3). De igual manera, en la estructura horizontal del bosque se determinó que en el transecto 4 se presentaron los individuos con más altura (13.59 m) y diámetro, (48.99 cm), lo que es muy superior al diámetro registrado en el transecto 1, con 20.11 cm, que corresponde a la mitad de su tamaño (Cuadro 4).
Cuadro 3 Distribución de individuos por clase diamétrica y altimétrica, densidad de las especies presentes en el área evaluada del BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Clase diamétrica (cm) | Individuos (Número) | Clase altimétrica (m) | Individuos (Número) | Densidad | Especies (Número) |
|---|---|---|---|---|---|
| 5 - 9.9 | 86 | 0 - 4.9 | 9 | Abundante | 4 |
| 10 - 19.9 | 128 | 5 - 9.9 | 205 | Poco abundante | 8 |
| 20 - 29.9 | 77 | 10 - 14.9 | 129 | Rara o escasa | 5 |
| 30 - 39.9 | 53 | 15 - 19.9 | 44 | ||
| 40 - 49.9 | 29 | 20 - 24.9 | 15 | ||
| 50 - 59.9 | 17 | 25 - 29.9 | 5 | ||
| 60 - 69.9 | 4 | 30 - 34.9 | 2 | ||
| 70 - 79.9 | 2 | 35 - 39.9 | 1 | ||
| 80 - 89.9 | 2 | ||||
| 90 - 99.9 | 0 | ||||
| 100 - 109.9 | 1 | ||||
| 110 - 119.9 | 2 | ||||
| 120 - 129.9 | 3 | ||||
| 130 - 139.9 | 1 | ||||
| 140 - 149.9 | 0 | ||||
| 150 - 159.9 | 0 | ||||
| 160 - 169.9 | 1 | ||||
| 170 - 179.9 | 1 | ||||
| 180 - 189.9 | 0 | ||||
| 190 - 199.9 | 1 | ||||
| 200 - 209.9 | 0 | ||||
| 210 - 219.9 | 2 | ||||
| Total | 410 | 410 | 17 |
Cuadro 4 Estructura horizontal del bosque, a partir de la distribución de árboles por clase diamétrica y de altura, de las especies presentes en los transectos del área evaluada del BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Transecto | Núm. individuos | Altura (m) | Diámetro(cm) |
|---|---|---|---|
| Media ± ds | Media ± ds | ||
| 1 | 110 | 10.21 ± 4.35 | 20.11 ± 12.28 |
| 2 | 42 | 10.33 ± 4.99 | 21.65 ± 12.52 |
| 3 | 194 | 9.58 ± 3.74 | 23.11 ± 24.25 |
| 4 | 64 | 13.59 ± 6.71 | 48.99 ± 44.92 |
La especie con el mayor número de individuos y, por lo tanto, con la más densidad relativa (DER) fue V. macracantha, con 154 (37.6 %), seguido de C. iguanaea con 55 (13.4 %) y Erythrina smithiana Krukoff. con 32 (7.8 %); por consiguiente, a Fabaceae (59.4 %) y Cannabaceae (13.1 %) les correspondieron los valores más altos; en tanto que, el taxon con el menor número de individuos fue B. sulcata (0.2 %) (Cuadros 1 y 4). La frecuencia relativa (FER) alcanzó el máximo valor (8.51) en varios taxones: V. macracantha, L. huasango, P. excelsum, A. multiflora, F. obtusifolia y C. iguanaea. V. macracantha, C. lutea, E. smithiana y C. iguanaea se consideraron abundantes; ocho poco abundantes y cinco raras o escasas: B. graveolens, C. glabrata, E. ruizii, C. iguanaea y B. sulcata, esta última, con apenas un individuo (Cuadro 3).
Área basal
El máximo valor de DAP correspondió a B. sulcata con 124.10 cm, seguido por F. obtusifolia, (120.06 cm), C. iguanaea (38.48 cm) y P. limensis (37.31 cm); aunque, solamente se identificó un ejemplar de B. sulcata y ocho de P. limensis (Cuadro 1). Con excepción de la clase 5-9.9 cm DAP con 86 individuos, el análisis de la distribución diamétrica indicó que la mayoría de los individuos en los transectos se reunieron en las clases 10 - 19.9 cm DAP (205) y 20 - 29.9 cm DAP (129), cuya proporción disminuyó a medida que aumentaban los diámetros (Cuadro 3). El valor acumulado de área basal fue de 343.86 m2 ha-1, conformada por B. sulcata 139.28 m2 ha-1 y F. obtusifolia (120.90 m2 ha-1). La mayor cobertura relativa (COB) o área basal relativa más amplia coincidió con las mismas especies (B. sulcata, con 39.19 y F. obtusifolia, con 36.68), seguidas de manera muy distante por C. iguanaea (3.77), P. limensis (3.54) y E. smithiana (3.30); y a nivel de familia destacaron Lauraceae (39.19), Moraceae (36.68) y Fabaceae (11.28) (Cuadro 5).
Cuadro 5 Variables ecológicas de las especies presentes en el área evaluada del BES Cerro Tres Puntas (Salas-Motupe, Lambayeque, Perú).
| Familia/Especie | Núm. esp. | Núm. ind. | DER | FER | Área basal (m2) | Área Basal relativa (COB) | IVI | VIF |
|---|---|---|---|---|---|---|---|---|
| Anacardiaceae | ||||||||
| Loxopterygium huasango Spruce ex Engl. | 20 | 4.88 | 8.51 | 6.06 | 1.97 | 15.35 | ||
| Total Familia | 1 | 20 | 4.88 | 8.51 | 6.06 | 1.97 | 15.35 | 12.73 |
| Boraginaceae | ||||||||
| Cordia lutea Lam. | 29 | 7.07 | 6.38 | 0.92 | 0.30 | 13.74 | ||
| Total Familia | 1 | 29 | 7.07 | 6.38 | 0.92 | 0.30 | 13.74 | 13.27 |
| Burseraceae | ||||||||
| Bursera graveolens (Kunth) Triana & Planch. | 4 | 0.98 | 4.26 | 3.95 | 1.28 | 6.53 | ||
| Total Familia | 1 | 4 | 0.98 | 4.26 | 3.95 | 1.28 | 6.53 | 8.16 |
| Fabaceae | ||||||||
| Albizia multiflora (Kunth) Barneby & J.W. Grimes | 8 | 1.95 | 8.51 | 8.75 | 2.84 | 13.30 | ||
| Caesalpinia glabrata Kunth | 5 | 1.22 | 2.13 | 1.11 | 0.36 | 3.71 | ||
| Erythrina smithiana Krukoff | 32 | 7.80 | 6.38 | 10.19 | 3.30 | 17.49 | ||
| Pithecellobium excelsum (Kunth) Mart. | 21 | 5.12 | 8.51 | 0.96 | 0.31 | 13.94 | ||
| Prosopis limensis Benth. | 9 | 1.95 | 2.13 | 10.93 | 3.54 | 7.62 | ||
| Senna atomaria (L.) Irwing & Barneby | 15 | 3.66 | 4.26 | 0.76 | 0.25 | 8.16 | ||
| Vachellia macracantha (Willd.) Seigler & Ebinger | 154 | 37.56 | 8.51 | 2.09 | 0.68 | 46.75 | ||
| Total Familia | 7 | 244 | 59.40 | 40.50 | 34.79 | 11.28 | 110.97 | 111.86 |
| Lauraceae | ||||||||
| Beilschmiedia sulcata (Ruiz & Pav.) Kosterm | 1 | 0.24 | 2.13 | 120.90 | 39.19 | 41.57 | ||
| Total Familia | 1 | 1 | 0.24 | 2.13 | 120.90 | 39.19 | 41.57 | 45.33 |
| Malvaceae | ||||||||
| Eriotheca ruizii (K. Schum.) A. Robyns | 6 | 1.46 | 2.13 | 9.74 | 2.49 | 6.80 | ||
| Guazuma ulmifolia Lam. | 11 | 2.68 | 6.38 | 6.53 | 1.91 | 10.98 | ||
| Total Familia | 2 | 17 | 4.16 | 8.52 | 16.27 | 4.40 | 17.78 | 20.32 |
| Moraceae | ||||||||
| Ficus obtusifolia Kunth | 17 | 4.16 | 8.51 | 139.28 | 36.68 | 49.34 | ||
| Total Familia | 1 | 17 | 4.16 | 8.53 | 139.28 | 36.68 | 49.34 | 46.74 |
| Olacaceae | ||||||||
| Ximenia americana L. | 17 | 4.15 | 6.38 | 0.92 | 0.33 | 10.86 | ||
| Total Familia | 1 | 17 | 4.15 | 6.38 | 0.92 | 0.33 | 10.86 | 10.38 |
| Sapindaceae | ||||||||
| Sapindus saponaria L. | 6 | 1.46 | 6.38 | 2.69 | 0.79 | 8.63 | ||
| Total Familia | 1 | 6 | 1.46 | 6.38 | 2.69 | 0.79 | 8.63 | 8.15 |
| Cannabaceae | ||||||||
| Celtis iguanaea (Jacq.) Sarg. | 55 | 13.41 | 8.51 | 13.38 | 3.77 | 25.70 | ||
| Total Familia | 1 | 55 | 13.41 | 8.51 | 13.38 | 3.77 | 25.70 | 23.08 |
| Total | 17 | 410 | 100 | 100 | 343.86 | 100 | 300 | 300 |
Índice de Valor de Importancia por Especie (IVI) e Índice de Valor de Importancia por Familia (VIF)
El análisis del IVI de las 17 especies evaluadas destacó a F. obtusifolia (49.34) sobre V. macracantha (46.75) y C. ruiziana (41.57); a su vez, a Fabaceae (110.97), Moraceae (49.34) y Lauraceae (41.57) (Cuadro 5). Estas familias registraron el VIF más alto (Fabaceae, 111.86; Moraceae, 46.74 y Lauraceae, 45.33); mientras que, las de valor más bajo fueron Burseraceae (8.16) y Sapindaceae (8.15) (Cuadro 5).
Discusión
El área de muestreo corresponde a la clasificación de bosques estacionalmente secos de montaña en la costa norte que cubre los BES sobre los 500 msnm, y abarca los flancos occidentales de los Andes en las regiones de Tumbes, Piura y Lambayeque, hasta La Libertad, incluida parte de Cajamarca (Linares-Palomino y Pennington, 2007). Sin embargo, como lo indica La Torre-Cuadros y Linares-Palomino (2008) este límite altitudinal es bastante variable, discutible y depende de varios factores.
Dado que el muestreo inició en los 373 msnm, incorporó una pequeña área de llanura en la costa norte que cubre los BES debajo de los 500 msnm a lo largo de los flancos occidentales de los Andes, desde Tumbes hasta La Libertad (Linares-Palomino y Pennington, 2007). Esta sería la razón por la que en las condiciones de Pilasca se observó escasa presencia de P. limensis (algarrobo), solamente un espécimen de C. scabridum (zapote) y otros de Beautempsia avicennifolia (Kunth) Gaudich. (vichayo); estos últimos fuera del área de muestreo.
Los mismos autores indicaron, además, que los BES secos de montaña en la costa norte, al igual que los del sistema interandino del río Marañón son las más ricos en especies leñosas con 193 y 184, respectivamente. Esa información es corroborada con el estudio del bosque tropical estacionalmente seco (BTES) del Valle del Marañón, en el norte del Perú, en el que se identifican 440 taxa leñosos y 143 (33 %) endémicas, así como dos centros de endemismo dentro del Valle, con 56 % de especies y 78 % de los géneros identificados en los BTES peruanos (Marcelo-Peña et al., 2016).
En el análisis de los patrones de diversidad de las especies (4 660 leñosas) de los BTES de América Latina y el Caribe, se determinó que la notable rotación florística observada implica la necesidad de establecer múltiples áreas de conservación para proteger esa importante diversidad florística (DRYFLOR et al., 2016).
A pesar de que una lista anotada de los BES del Ecuador y otros estudios relacionados (Svenson, 1946; Gentry, 1995b; Aguirre et al., 2006) señalaron que los BES de ambos países, en especial los de la costa, formaban una sola unidad geográfica, con alto número de taxones endémicos (Centro de Endemismo Tumbesino). Las pocas especies (17), géneros (17) y familias (10) inventariadas en el Cerro Tres Puntas de Pilasca demostrarían un alto grado de depredación de la flora original; ello corroboraría lo observado en otros BES, como Chaparrí en Lambayeque (Linares-Palomino y Ponce-Álvarez, 2009; Martos et al., 2009) y La Menta y Timbes en Piura (Rasal et al., 2011).
Por otro lado, habría que considerar otros factores, como los gradientes de diversidad y densidad de especies, así como de precipitación que disminuyen conforme los BES se extienden hacia el sur, y que incluyen a las regiones de Lambayeque y La Libertad.
Por la agreste y peligrosa topografía del terreno, el muestreo lineal que se utilizó en la presente investigación hizo posible evaluar un área extensa que abarcó 11.4 ha, muestreo que se ha usado en los inventarios botánicos rápidos (IBR) en los que se aborda la vegetación leñosa, endemismos y el estado de conservación de los BES de Jaén (Perú) (Marcelo-Peña, 2008), con base en la metodología de Hawthorne y Abu-Juam (1995) y Gordon et al. (2004).
A pesar de que en los últimos años el interés por los BES se ha incrementado (Espinosa et al., 2012; Aguirre y Geada-López, 2017), en los escasos trabajos efectuados en Ecuador y Perú, se observó que los métodos de muestreo fueron muy variados entre el uso de los transectos rectangulares y el muestreo lineal, así como en el tamaño de las unidades muestrales, desde 0.25 hasta 6.5 hectáreas; y en el DAP de las especies muestreadas, de ≥ 1.0 a 10 cm. A esto habría de agregarse la diferencia entre el número de individuos, especies, géneros y familias que suele ser más alta en áreas naturales protegidas como el Parque Nacional Cerros de Amotape (Linares-Palomino, 2005) y muy deprimida en otras áreas históricamente depredadas, como la Reserva Ecológica Chaparrí (Linares-Palomino y Ponce-Álvarez, 2009).
De todos ellos, el que más se aproximó al aquí descrito es el de La Menta y Timbes en Piura (Rasal et al., 2011), donde no obstante que se registraron especies leñosas con DAP ≥ 1.0 cm, el número de individuos inventariados fue significativamente menor, aunque hubo bastante similitud en el número de especies, géneros y familias.
En su estudio de la Reserva Ecológica Chaparrí, Linares-Palomino y Ponce-Álvarez (2009) puntualizaron la dificultad de establecer comparaciones directas de inventarios en los bosques estacionalmente secos de la región, debido a la gran variación entre las metodologías de muestreo utilizadas, pisos altitudinales, área total de muestreo, DAP de los especímenes muestreados, estado de conservación y de depredación del área en el momento de su realización, e incluso en la priorización de la información recolectada, entre otros factores.
Lo anterior se confirma con los resultados de varios estudios en Perú; por ejemplo, en los bosques de Jaén, Cajamarca (Marcelo-Peña, 2008); La Menta y Timbes en Ayabaca, Piura (Rasal et al., 2011); Reserva de la Biosfera del Noroeste del Perú (Parque Nacional Cerros de Amotape y Zona Reservada de Tumbes) (Leal-Pinedo y Linares-Palomino, 2005); Llanuras Costeras de Piura (La Torre-Cuadros y Linares-Palomino, 2008); Parque Nacional Cerros de Amotape (Linares-Palomino, 2005); y Reserva Ecológica Chaparrí, Lambayeque (Linares-Palomino y Ponce-Álvarez, 2009). Sin embargo, no ha sido posible establecer patrones rigurosos de comparación entre toda esta valiosa información y el estudio desarrollado en el Cerro Tres Puntas de Pilasca.
En las formaciones vegetales de los BES de Ecuador y Perú se elaboraron listas de numerosas especies leñosas para el bosque deciduo, bosque seco semideciduo mezclado con especies siempre verdes, bosque caducifolio y el matorral espinoso seco (Aguirre et al., 2006; Aguirre y Kvist, 2009); No obstante, en el trabajo aquí descrito, solamente, se registraron ocho especies: P. limensis, C. lutea, P. excelsum, C. glabrata, V. macracantha, E. smithiana, B. graveolens y L. huasango, lo que, de alguna manera, indicaría la fuerte presión antrópica a la que está sujeta la vegetación del Cerro Tres Puntas de Pilasca en los últimos años, aun cuando fue posible encontrar individuos esporádicos de Bougainvillea peruviana Nees & Mart., Bauhinia aculeata L., C. vitifolium y C. scabridum; aunque estaban fuera del transecto lineal.
La tendencia de distribución de las alturas, que no se ajusta exactamente con la “J” invertida, descrita por Lamprecht (1990) y Günter et al. (2011) reflejaría la característica sucesional del bosque debido a los disturbios constantes y selectivos que ejercen los habitantes en el lugar, manteniéndolo en una sucesión secundaria (Rasal et al., 2012). Aunque en la experiencia referida no se incluyeron individuos con DAP < 5.0 cm, pero sí con altura de planta < 4.9 m, los resultados permiten alcanzar un cabal conocimiento sobre el proceso de regeneración de las especies presentes en las clases diamétricas y de altura mayores, como fue recomendado por García et al. (2010) al estudiar la estructura y diversidad florística de dos bosques naturales en el Cauca, Colombia.
En los BES La Menta y Timbes de Piura (Perú), alrededor de 55.0 % de los individuos muestreados tuvieron un DAP entre 0-10.0 cm con algunos individuos de L. huasango y B. graveolens con DAP entre 50-60 cm, para La Menta; y Ceiba trischistandra Bakh. y E. ruizii con DAP entre 80-90 cm, para Timbes (Rasal et al., 2011). Los resultados que se presentan para el BES Cerro Tres Puntas de las cinco especies indicadas solo se registró E. ruizii. Asimismo, de 410 individuos inventariados, 196 tuvieron DAP ≥ 20.0 cm, y tres especies: V. macracantha, C. iguanaea y E. smithiana contribuyeron con 58.8 % del total cuantificado.
En muy pocos estudios de los bosques secos del Perú se ha recabado información o se han efectuado análisis relacionados con AB (Área Basal), IVI (Índice de Valor de Importancia) y VIF (Valor de Importancia para la Familia); sin embargo, Linares-Palomino y Ponce-Álvarez (2009) calcularon un AB de 11.74 m2, en el que sobresalieron E. ruizii y L. huasango con 4.40 y 4.12 m2, respectivamente en la Reserva Ecológica de Chaparrí (Lambayeque). En los bosques secos de La Menta y Timbes (Piura), Rasal et al. (2011) obtuvieron, para La Menta, un AB de 128.86 m2, en donde destacaron L. huasango (45.97 m2) y B. graveolens (34.69 m2); en Timbes se obtuvieron un AB de 196.63 m2, con especial aportación de E. smithiana (47.16 m2) y C. trischistandra (38.88 m2). En ambos casos, el AB fue muy inferior al estimado en el presente estudio, cuya AB fue de 343.86 m2 ha-1, lo que indicó que a pesar de que en el bosque Cerro Tres Puntas existe una fuerte presión antrópica, esta no ha resultado tan dramática como ocurre en Chaparrí, La Menta y Timbes, donde incluso al bosque de Chaparrí se le definió como una versión empobrecida de los bosques estacionalmente secos de la región (Linares-Palomino y Ponce-Álvarez, 2009).
En esta misma comunidad, cuatro especies alcanzaron valores de IVI superiores a 30.0: E. ruizii (58.22), L. huasango (44.93), C. lutea (33.76) y B. graveolens (30.08), que correspondieron a las familias Malvaceae, Anacardiaceae, Boraginaceae y Burseraceae, respectivamente (Linares-Palomino y Ponce-Álvarez, 2009); en tanto que, en La Menta, los valores más altos fueron para B. graveolens (50.0), L. huasango (43.0) y C. lutea (36.0); y en Timbes, a C. lutea (62.0), E. smithiana (40.0) y C. trischistandra (25.0) (Rasal et al., 2011). Asimismo, como se ha referido anteriormente, en el trabajo de los bosques secos en la provincia de Loja, Aguirre et al. (2013) determinaron las cinco especies con el IVI más alto: C. trischistandra, Simira ecuadorensis (Standl.) Steyerm., Tabebuia chrysantha G. Nicholson, E. ruizii y Terminalia valverdeae A. H. Gentry; aunque ninguna información cuantitativa fue proporcionada. Al comparar esos resultados con los que aquí se documentan, se observan diferencias notorias; puesto que las especies que destacaron ampliamente fueron F. obtusifolia (49.34), V. macracantha (46.75) y B. sulcata (41.57), las cuales fueron relevantes en otros bosques estacionales. Es posible enunciar la hipótesis de que la depredación de las especies en estos bosques no tiene un patrón definido, y ello habría estado supeditado a la preferencia por las especies maderables más valiosas, así como a la misma protección que les habrían dado los pobladores del lugar, por lo que, ciertamente, en el Cerro Tres Puntas la depredación ha sido más selectiva.
Los resultados del VIF de la investigación descrita para el Cerro Tres Puntas de Pilasca en la que sobresalieron las familias Fabaceae (111.86), Moraceae (46.74) y Lauraceae (45.33), discrepan ampliamente de las referidas para Chaparrí (Linares-Palomino y Ponce-Álvarez, 2009), La Menta y Timbes (Rasal et al., 2011); ya que ninguna de las familias consignadas en esos trabajos fue identificada en el estudio realizado en el área de interés.
Conclusiones
El estudio realizado en el Cerro Tres Puntas de Pilasca (Salas-Motupe, Lambayeque) contribuye al conocimiento de los ecosistemas naturales del Perú, específicamente, el de los bosques estacionalmente secos, ya que determina su estructura y composición florística. En el contexto de la biodiversidad, los resultados describen su riqueza vegetal, lo que es una aportación valiosa para efectos de conservación y manejo de dichos recursos, y los datos ecológicos constituyen una aproximación preliminar a la dinámica de los BES de la región referida.
Se sugiere ampliar los estudios a otras áreas en el mismo Cerro Tres Puntas, en áreas geográficas adyacentes, en otros BES de Lambayeque como los de Tongorrape, Ñaupe y Tocmoche e incluso incrementar el intervalo altitudinal del muestreo. Asimismo, orientar las evaluaciones sobre captura de carbono e identificar las especies que más contribuirían a mitigar el cambio climático para propender su multiplicación a gran escala con fines de reforestación.

