











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La ecología espacial ofrece diversas herramientas que evalúan el grado de agregación y permiten inferir la escala de variación de los parámetros ecológicos (Ver Hoef et al., 2018). En general, se trata de métodos que descomponen la variabilidad de un parámetro y que definen las escalas de análisis en función de distintos radios de acción en los que se establecen distancias a vecinos más próximos (Dale y Fortin, 2014). Todos los métodos determinan y describen los patrones espaciales que permiten evaluar hipótesis sobre diversos procesos ecológicos asociados con la dinámica forestal de un ecosistema (Rozas y Camarero, 2005; Martensen et al., 2017).
Condit et al. (2002) plantean que el análisis del patrón espacial describe, cuantitativamente, la distribución horizontal de los individuos en una comunidad, y es crucial para entender como un grupo de especies usa los recursos del entorno. De acuerdo con Hubbell (2001), existen tres patrones generales de distribución: i) aleatorio, donde los individuos están distribuidos al azar por la homogeneidad ambiental y siguen un patrón no selectivo; ii) uniforme, que se presenta a ciertas distancias, como resultado de las interacciones negativas entre individuos; y iii) agregado, en el cual los individuos de una especie están aislados de otras, por la limitación en dispersión o especialización de hábitat.
Existen varios métodos para identificar los patrones espaciales en forma de índices (Aguirre-Calderón, 2002; Rozas y Camarero, 2005; Ledo et al., 2012; Dale y Fortin, 2014), entre los más robustos se pueden citar al de Morisita-Horn (Morisita, 1959), que estima la variación del tipo y la intensidad del patrón a lo largo de un gradiente escalar espacial; la función K de Ripley, que analiza la distancia al vecino más próximo e informa sobre el patrón espacial observado como una función de la distancia d (Ripley, 1977; Dixon, 2013).
En selvas tropicales se ha descrito que las especies se distribuyen de acuerdo a jerarquías de abundancia, y lo común es que haya menos taxones raros, pocos abundantes y muchos con una abundancia intermedia (Martínez-Ramos y Álvarez-Buylla, 1995; Franklin y Rey, 2007; Yili et al., 2013). Algunos autores (John et al., 2007; Blach-Overgaard et al., 2010) plantean que las principales limitantes para que una especie exhiba determinado patrón espacial son de tipo abiótico (altitud, latitud, temperatura, luminosidad, precipitación, evapotranspiración, tipo de suelo, topografía y pendiente) y biótico (competencia intra e interespecífica y depredación).
De acuerdo con Fossa (2004) y Hernández et al. (2018), la evaluación de los patrones espaciales a lo largo de gradientes altitudinales es una herramienta fundamental para predecir el comportamiento de las especies y la dinámica de flujo vegetal entre estratos, particularmente, ante escenarios de cambio climático y con propósitos de conservación (Cruz, 2013). Los meta-análisis a nivel global han planteado que las especies arbóreas se están desplazando hacia altitudes mayores, producto del cambio climático, y, precisamente, en las zonas altas de los trópicos se han registrado con mayor constancia estos cambios, debido a la reducción del intervalo de distribución de dichos taxa (Parmesan, 2006; Brown et al., 2015).
Montañez et al. (2010) proponen que a mayor altitud se incrementa la probabilidad de encontrar un patrón agrupado de individuos arbóreos, por lo que se infiere que en las tierras bajas tropicales el patrón de distribución será diferente (Hubbell, 2001). En consecuencia, las especies que no logren adaptarse a estos cambios ambientales desaparecerán por estrés fisiológico (Parmesan, 2006; Brown et al., 2015), y las que sí, serán afectadas sus relaciones interespecíficas actuales (Root et al., 2003).
En México, Martínez-Ramos y Álvarez-Buylla (1995) indican que los árboles en las selvas húmedas se distribuyen, por lo general, de manera agregada, debido a que presentan una baja dispersión y alta mortalidad en las etapas tempranas. En contraste, Zarco et al. (2010) en una selva de Tabasco señalan una distribución, principalmente, aleatoria para las especies arbóreas; aunque en algunos sitios observaron un patrón agregado en el estrato inferior; lo cual es similar a lo citado en los trabajos de Martínez-Ramos (2006), quien registró un patrón agregado en la Reserva de la Biósfera Montes Azules, Chiapas, que relaciona con terrenos de topografía irregular y a la disponibilidad lumínica en el sotobosque. Pese a estas investigaciones, los estudios de distribución espacial a lo largo de gradientes altitudinales son escasos en el país (Hernández et al., 2018), y más aún en selvas tropicales.
Con base en lo expuesto, destaca la importancia de identificar los patrones espaciales de distribución de los taxa arbóreos en las selvas tropicales, dada la alta complejidad funcional y estructural de esos ambientes (Condit et al., 2002; Lohbeck et al., 2015), así como a las amenazas que enfrentan (Arroyo-Rodríguez et al., 2017).
Particularmente en Chiapas, la Selva Lacandona es uno de los ecosistemas más frágiles, cuya cobertura forestal se ha reducido 70 % en las últimas décadas debido a la expansión agrícola-ganadera y a la extracción ilegal de madera (Martínez-Ramos, 2006); mismos factores que afectan a varias de sus áreas naturales protegidas, como es el caso del Área de Protección de Flora y Fauna Metzabok (Conanp, 2006).
El presente trabajo tiene como objetivo analizar y describir el patrón de distribución y correlación espacial de las especies arbóreas a lo largo de un gradiente altitudinal en el Área Natural Protegida Metzabok, Selva Lacandona, Chiapas. Se espera que el patrón de distribución espacial sea diferente entre etapas de desarrollo (brinzal, latizal y fustal), categorías de altura (inferior, intermedia, superior) y a lo largo del gradiente altitudinal, independientemente del índice utilizado.
Materiales y Métodos
Área de estudio
El presente trabajo se realizó en el Área Natural Protegida (ANP) Metzabok, localizada al noreste de la Selva Lacandona (17°08’36’’ latitud norte y 91°34’42’’ longitud oeste), en la subcomunidad lacandona Puerto Bello Metzabok, Ocosingo, Chiapas. Esta ANP tiene una superficie de 3 368 ha, y se ubica a una altitud de 545-950 m (Conanp, 2006). El clima es Af(m)w”(i)g, cálido húmedo con lluvias todo el año, temperatura media anual de 24 °C y una precipitación media anual de 3 160 mm (García, 2004). Los tipos de suelo corresponden a: redzinas, luvisoles, gleysoles, vertisoles, cambisoles y litosoles (Sánchez-Gutiérrez et al., 2017). De acuerdo con la clasificación de Miranda y Hernández-X (2014), la vegetación predominante es la Selva Alta Perennifolia.
Estratificación altitudinal y muestreo
Se determinaron cinco estratos altitudinales (EA1= < 600 msnm, EA2= 600-700 msnm, EA3= 700-800 msnm, EA4= 800-900 msnm y EA5= > 900 msnm) con base en la generación de curvas de nivel en intervalos de 100 m con el Software Global Mapper v15. En cada estrato se instaló una unidad de muestreo (UM) de 20 × 50 m (1 000 m2), como se muestra en la Figura 1; estas se dividieron en 10 subunidades de muestreo (SUM), de 10 × 10 m (100 m2). En cada SUM todos los individuos de la clase fustal (árboles reproductivos) con diámetro normal (DN) ≥ 2.5 cm fueron medidos con cinta diamétrica (marca Richter Messwerkzeuge modelo 283D/10M) y ubicados mediante sus coordenadas cartesianas (x, y) (Linzaga-Román et al., 2011; Dale y Fortin, 2014), se consideró la esquina inferior izquierda del plano con respecto a la pendiente de cada UM como el origen (0, 0) (Ruiz-Aquino et al., 2015). Dentro de cada SUM se instaló al azar un cuadro de 5 x 5 m para el registro de los individuos de la clase latizal (juveniles con DN < 2.5 cm y altura > 1.5 m), y al interior de estos se establecieron dos cuadros de 2 × 2 m, para el reconocimiento de los individuos categorizados como brinzales (plántulas con una altura < 1.5 m) (Zarco et al., 2010).

5Figura 1 Ubicación de las unidades de muestreo (UM) por estrato altitudinal (EA) en el Área Natural Protegida Metzabok, Selva Lacandona, Chiapas.
La identidad taxonómica de los individuos se determinó con el apoyo del personal del Herbario “ECOSC-H”, del Colegio de la Frontera Sur (Ecosur) de San Cristóbal de las Casas, Chiapas. La nomenclatura se validó en el sitio web Tropicos.org del Missouri Botanical Garden (2008).
Análisis de distribución espacial
Para calcular la distribución espacial de los fustales se utilizaron tres categorías de alturas (inferior, intermedia, superior) obtenidas en las curvas de inflexión del trabajo de Sánchez-Gutiérrez et al. (2017). La determinación de los patrones de distribución espacial de los estados de desarrollo brinzal y latizal, así como de las categorías de altura de los fustales, se llevó a cabo mediante el índice de Morisita-Horn (Morisita, 1959); cuando su valor es igual a 1, la distribución es aleatoria, mayor que 1, agregada y menor que 1, uniforme (Morisita, 1959; Zarco et al., 2010; Rivera-Fernández et al., 2012). El índice de Morisita-Horn se define en la ecuación (1):
Donde:
q = Número de cuadros
ni = Número de individuos en el i-ésimo cuadro
N = Número total de individuos en todos los q cuadros
Con el propósito de obtener un índice estadísticamente más robusto y gráficamente representativo (Rozas y Camarero, 2005; Ledo et al., 2012; Dale y Fortin, 2014), también se calculó la función K(t) de Ripley (Ripley, 1977) para la distribución espacial de las categorías de altura de los fustales, a partir de la ecuación (2):
Donde:
n = Número de individuos en la unidad de muestreo (UM-A)
(A( = Área de la UM-A (m2)
u ij= Distancia entre i-esimo y j-esimo individuo en la UM-A
t = Distancia entre dos árboles (m)
I t (u) = 1 si u ( t y 0 u > t
W ij = Proporción de la circunferencia de un círculo con centro en UM-A (la sumatoria de todos los pares de individuos no mayor que t)
Para estabilizar la varianza, se utilizó la transformación de la raíz cuadrada de K(t) de Ripley, y para determinar la significancia (p ≤ 0.01) de la función L(t) se realizaron simulaciones Monte Carlo (Besag, 1977). Los límites máximos y mínimos de L(t) se definieron con 99 % de confiabilidad:
Donde:
K(t) = Función univariada de Ripley
π = 3.1416
t = Distancia (m)
Si el valor de L(t) = 0, el patrón espacial con radio t es aleatorio, L(t) > 0 indica agrupación, y L(t) < 0, un patrón uniforme.
Correlación espacial
Se analizó la correlación espacial entre categorías de alturas (inferior-intermedia, inferior-superior e intermedia-superior) de las especies arbóreas de la clase fustal, mediante la función bivariada L 12 (t), derivada de la K 12 (t) (Ripley, 1977):
Donde:
K 12 (t) = Función bivariada de Ripley
π = 3.1416
t = Distancia (m)
Un valor de L 12 (t) = 0 indica que los dos grupos son espacialmente independientes, L 12 (t) > 0 refiere a una asociación positiva (atracción), y L 12 (t) < 0 expresa una asociación negativa (repulsión). La significancia (p ≤ 0.01) de L 12 (t) se determinó por simulaciones de Monte Carlo (Besag, 1977; Dale y Fortin, 2014), y los límites máximos y mínimos se definieron con 99 % de confiabilidad. Los patrones univariados y bivariados de Ripley se generaron con el programa Toolbox (Fisher, 2000).
Resultados y Discusión
Patrones de distribución espacial de especies arbóreas
De acuerdo al índice de Morisita-Horn, el patrón de distribución
espacial de las especies arbóreas para la clase fustal en los cinco EA y para
las tres categorías de altura (inferior, intermedia y superior) tuvo un
comportamiento promedio tipo agregado (
Cuadro 1 Valores del índice de Morisita-Horn para fustales (categorías de altura: inferior, intermedia, superior) latizales y brinzales en cinco estratos altitudinales (EA) del Área Natural Potegida Metzabok, Selva Lacandona, Chiapas.
| Estrato | Fustales | Latizales | Brinzales | ||
|---|---|---|---|---|---|
| Altitudinal (EA) | Inferior | Intermedia | Superior | ||
| EA1 (< 600) | 1 | 1 | 1.1 | 1.3 | 1.1 |
| EA2 (600 - 700) | 1 | 1.3 | 1.3 | 1.1 | 1.1 |
| EA3 (700 - 800) | 1 | 1 | 0.8 | 1.1 | 1.2 |
| EA 4 (800 - 900) | 1.1 | 1 | 1 | 1.1 | 1.6 |
| EA5 (> 900) | 1.3 | 1 | 0.8 | 1.0 | 1.0 |
Cabe precisar que el patrón general registrado en el presente estudio muestra diferencias al analizarse por EA. La categoría de altura inferior presentó un patrón aleatorio y agregado a menor y mayor altitud, respectivamente (EA4 y 5) (Cuadro 1). Se considera que el patrón agregado puede deberse a los claros formados por los disturbios naturales: caída o muerte de los árboles, plagas y enfermedades (Brown et al., 2015); mismos que son colonizados por especies pioneras, cuya regeneración depende de luz solar y semillas con menor alcance de dispersión (Hubbell, 2001).
Sin embargo, la presencia de ambos patrones espaciales (aleatorio y agregado) registrados en el presente estudio pueden ser, también, el resultado de las actividades humanas que han modificado el hábitat y afectado, de manera directa, la sobrevivencia, reproducción y establecimiento de las especies, como lo demostraron Pavón-Hernández y Rico-Gray (2004) para un paisaje transformado en el estado de Veracruz, México.
La categoría de altura intermedia mostró un comportamiento aleatorio
Algunas otras investigaciones (Blach‐Overgaard et al., 2010) han demostrado que el patrón agregado en las selvas tropicales se explica porque la temporada de lluvias está asociada a vientos fuertes, que promueven la caída de los árboles, lo que forma microclimas que propician la repoblación en grupos. Martínez-Ramos (2006) documentan que el patrón agregado está relacionado con las características topográficas y edáficas; pero, además, se sugiere que puede deberse a deficientes mecanismos de dispersión de semillas y baja depredación de las mismas (Franklin y Rey, 2007; Hosaka et al., 2017).
Para la categoría de altura superior, la distribución de las especies (Sebastiana longicuspis Standl, Pseudolmedia spuria (Sw.) Griseb, Manilkara zapota (L.) Royen y Pouteria reticulata (Engl.) Eyma) fue uniforme a mayor altitud y agregada, cuando disminuyó (Cuadro 1). Se considera que el patrón agregado en esa categoría se atribuye a la cercanía con la comunidad, ya que son áreas donde los comuneros extraen maderas para el uso doméstico, situación que genera la formación de claros que son aprovechados por un grupo de especies (Dialium guianense (Aubl.) Sandwith, Heliocarpus appendiculatus Turcz. y Pseudolmedia oxiphyllaria Donn. Sm.) que necesitan la luz directa para su desarrollo y crecimiento (Conanp, 2006; Rivera-Fernández et al., 2012).
Los estados de desarrollo latizal y brinzal presentaron un patrón agregado, con excepción del último EA5 (> 900 msnm), en el que se observó un comportamiento aleatorio (Cuadro 1). Los claros permiten la entrada de luz, lo cual afecta la incorporación y crecimiento de las plantas del sotobosque (Martínez-Ramos, 2006), como es el caso de los brinzales y latizales. Por lo tanto, cualquier alteración en las características del dosel influye en la germinación, tasa de crecimiento, sobrevivencia y distribución de las plantas (Brienen y Zuidema 2006; Yili et al., 2013), dadas las condiciones específicas dominantes al interior del claro que promueven la aglomeración de las especies.
Martínez-Ramos (2006) cita que el patrón agregado es mayor en individuos pequeños (1 a 5 cm de DN), en comparación con aquellos de mayores dimensiones (≥ 10 cm de DN), que se distribuyen de manera aleatoria. De hecho en el patrón aleatorio, los individuos son completamente independientes de la posición de cualquier otro en la población, comportamiento que se observa en ciertas especies de bosques templados (Hernández et al., 2018), pero es muy común en individuos arbóreos de las selvas tropicales (Franklin y Rey, 2007; Yili et al., 2013).
Los resultados de este estudio sugieren cierta tendencia con los gradientes altitudinales, e indican que en el ANP Metzabok existe una variación climática por gradiente que modifica la estructura y la composición de la selva, lo que concuerda con otras investigaciones sobre la estructura, composición y diversidad arbórea realizados en la misma área de estudio (Martínez-Ramos, 2006; Sánchez-Gutiérrez et al., 2017). En cambio, Cruz (2013) en Costa Rica registró que en altitudes de 400 a 900 m no existe influencia del gradiente altitudinal en el patrón de distribución espacial, y explica que puede deberse a que el intervalo de la altitud es, relativamente, pequeño; y por lo tanto, las condiciones climáticas no varían entre estratos. En consecuencia, se ha sugerido que los trabajos de vegetación en gradientes altitudinales deberían realizarse en extensiones geográficas amplias (Fossa, 2004).
Distribución espacial de especies arbóreas con la función K(t) de Ripley
De acuerdo con la función univariada de K(t) de Ripley, la distribución espacial de las tres categorías de altura de la clase fustal evidenció un patrón significativo (p ≤ 0.01) entre agregado, aleatorio y uniforme a diferentes distancias, por lo que se acepta la hipótesis planteada (Figura 2). Este resultado fue similar al obtenido con el índice de Morisita-Horn para los fustales (Cuadro 1).

Figura 2 Valores de distribución univariada L(t) para las tres categorías de altura de fustales en especies arbóreas en cinco estratos altitudinales (EA) en el Área Natural Protegida Metzabok, Selva Lacandona, Chiapas.
La categoría de altura inferior tuvo, en general, un patrón agregado, con variación a distintas distancias y estratos altitudinales (Figura 2). Los EA1 y 4 se distribuyen de forma agregada en distancias de 1 a 6 m, y uniforme a más de 7 m; en cambio, el EA2 tuvo un comportamiento aleatorio, y el EA3 a distancias de 1 a 2 m fue uniforme, mientras que de 3 a 10 m cambió a una distribución agregada (Figura 2). El EA5 mostró una distribución agregada en todas las distancias.
La variación en los patrones de distribución en los diferentes EA se relaciona con los claros generados por las categorías intermedias y superiores, ya que al ser afectados por algún disturbio (caída de ramas o el árbol completo, tala, muerte por plagas y enfermedades), dichas superficies son repobladas por especies demandantes de luz, que mediante procesos de mortalidad transforman su patrón inicial (agregado) a uno aleatorio (Franklin y Rey, 2007; Barreto-Silva et al., 2014). En el área de estudio, los claros son colonizados por Rinorea guatemalensis (S. Watson) Bartlett, lo cual concuerda con los resultados de Zarco et al. (2010) en selvas de Tabasco, quienes indican que esa especie se desarrolla y domina en el estrato inferior de las selvas donde hay disturbio.
La categoría de altura intermedia presentó una distribución, principalmente, aleatoria; aunque en distancias menores los árboles se ubican de manera uniforme, con excepción del EA2 que tiene una distribución agregada a distancias de 2 a 10 m (Figura 2). La distribución aleatoria en esta categoría se debe, sobre todo, a la extracción de madera que ocurrió durante la década de los 70 en el área de estudio (Conanp, 2006), que promovió la mortalidad de un conjunto heterogéneo de especies en el espacio. Los patrones agregados, en cambio, podrían ser una respuesta a la variabilidad ambiental entre estratos, y son comunes, ecológicamente, en individuos con dimensiones grandes (Dixon, 2013).
En la categoría de altura superior, los EA1 y 2 exhibieron un patrón agregado a distancias menores de 8 m (Figura 2), relacionado tanto con la muerte natural de algunos árboles viejos, como con la extracción selectiva de maderas preciosas como la caoba (Swietenia macrophylla King) y el cedro (Cedrela odorata L.). De hecho, en los años 1975 y 1984 se autorizó el aprovechamiento de 55 000 m3 y 20 000 m3 de corrientes tropicales en la Selva Lacandona (Vásquez-Sánchez et al., 1992), y continúa la extracción maderable para construcción de viviendas, por parte de los pobladores en ciertas zonas del ANP (Conanp, 2006), los cuales dejan en pie agrupaciones de M. zapota, P. reticulata, Manilkara chicle (Pittier) Gilly y Brosimun alicastrum Sw; especies de menor valor por la complejidad del manejo de su madera (duras y densas).
Lo anterior coincide con lo señalado por Franklin y Rey (2007) y Dale y Fortin (2014), quienes indican que la evolución lógica del patrón espacial al aumentar la edad de los individuos responde al efecto acumulado de los disturbios, la competencia y el ataque de patógenos. Por lo tanto, un patrón aleatorio puede cambiar a uno agregado o uniforme, en función de las características ambientales y antrópicas del medio.
Si bien, el comportamiento uniforme es poco común en los ensambles naturales, es posible encontrarlo en ambientes estables en donde existen disturbios mínimos (Antos y Parish, 2002). De hecho, el modelo propuesto por Janzen (1970) afirma que la mortalidad de los árboles tiene como consecuencia una distribución uniforme en los individuos adultos sobrevivientes; como lo corroboraron Lan et al. (2009) en las selvas tropicales de China donde observaron que, en ambientes con múltiples especies, estas tienden a distribuirse de forma uniforme.
El patrón espacial tan variable registrado en los EA3, 4 y 5 (Figura 2), se debe, posiblemente, al difícil acceso a dichos estratos, ya que no hay registro de aprovechamiento de madera, y es posible observar individuos de Calophyllum brasiliense Cambess., Terminalia amazonia (J.F. Gmel.) Exell, Guatteria anomala R.E. Fr., M. chicle y M. zapota hasta con 190 cm de diámetro y alturas superiores a 50 m. Algunos autores (Rozas y Camarero, 2005; Linzaga-Román et al., 2011) proponen que un comportamiento tan variable en el patrón de distribución se explica por los factores topográficos del sitio. De tal manera que es factible plantear que la distribución de las especies en el ANP de Metzabok podría asociarse tanto con la heterogeneidad ambiental inducida por el relieve, como con los disturbios ocasionados en cada sitio, como lo sugieren Montañez et al. (2010) y Barreto-Silva et al. (2014) para algunas selvas tropicales de Colombia.
Patrones de correlación espacial
De acuerdo con la correlación espacial de K 12 (t) de Ripley, los patrones bivariados de los EA2, 3 y 5 de la categoría de altura inferior-intermedia indicaron repulsión espacial en distancias menores a 2 m y atracción entre los 3 a 10 m (Figura 3). La repulsión a pequeña escala entre ambas categorías sería una consecuencia de la mortandad densodependiente de las plántulas situadas bajo las copas de las categorías intermedias y superiores, por lo que especies de las categorías inferiores (Myriocarpa longipes Liebm. y Piper psilorhachis C. DC.) colonizan los espacios disponibles, excepto bajo la copa de taxones de categorías superiores, como C. brasiliense, T. amazonia y G. anomala (Lan et al., 2009).
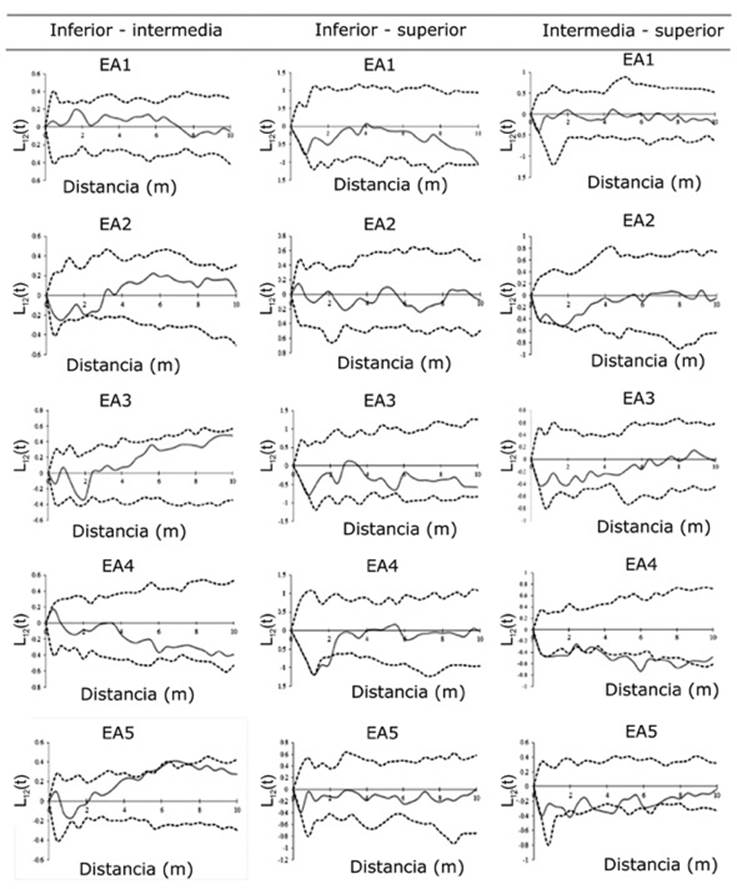
Figura 3 Matriz de relación de los valores de la distribución bivariada L 12 (t) para las tres categorías de altura de fustales de especies arbóreas en cinco estratos altitudinales (EA) en el Área Natural Protegida Metzabok, Selva Lacandona, Chiapas.
Ese comportamiento también es factible que responda a procesos de depredación, dado que es más probable que los depredadores eliminen a los individuos que se localizan más próximo al árbol semillero (Hosaka et al., 2017); lo cual explicaría la existencia de repulsión entre la categoría inferior-superior a distancias cercanas, y atracción en la intermedia-superior, a medida que incrementa la distancia respecto al árbol parental (Rozas y Camarero, 2005; Barreto-Silva et al., 2014).
Lo anterior contrasta con los EA1 al 4, en los cuales el patrón de correlación fue independiente, que coincide con el observado para todos los estratos de la categoría inferior-superior. En la intermedia-superior (Haematoxylum campechianum L. y Hauya elegans DC. vs. D. guianense y Pouteria durlandii (Standl.) Baehni, respectivamente) a diferentes distancias se presentan asociaciones negativas (repulsión), lo cual fue más notable en el EA4 (Figura 3). Este tipo de comportamiento quizás es una respuesta a la alta mortalidad de semillas e individuos inferiores cerca de los árboles superiores. Las enfermedades y la depredación se concentran en las inmediaciones de los individuos progenitores, lo que causa mortalidad vía denso-dependencia, ya que existe mayor probabilidad de supervivencia a medida que aumenta la distancia entre la descendencia y los adultos productores de semillas (Janzen, 1970; Franklin y Rey. 2007; Hosaka et al., 2017).
Conclusiones
El patrón de distribución espacial de las especies arbóreas está relacionado con la altitud, pero también tiene influencia de los disturbios naturales y antrópicos. Las especies de las etapas de desarrollo latizal (Chamaedorea oblongata Mart., Chamaedorea pinnatifrons (Jacq.) Oerst., P. oxiphyllaria y Chionanthus oblanceolatus (B.L. Rob.) P.S. Green) y brinzal (R. guatemalensis, Eugenia acapulcensis Steud., Aphananthe monoica (Hemsl.) J.-F. Leroy y P. reticulata) se presentan en forma agregada, ya que la mayoría se desarrolla en áreas perturbabas con existencia de claros, pero bajo el dosel de fustales residuales, dado el requerimiento de mayor intensidad de luz solar para su desarrollo y crecimiento.
El patrón de correlación espacial entre categorías de altura es variable. En la categoría inferior-intermedia (con especies dominantes como R. guatemalensis y P. psilorhachis vs. Garcinia intermedia (Pittier) Hammel y Trophis mexicana (Liebm.) Bureau) e intermedia-superior (con dominancia de H. campechianum L. y H. elegans vs. D. guianense y P. durlandii) hay repulsión a menor distancia, debido a la sombra que proyectan los individuos de mayor tamaño, lo cual impide la entrada de luz; y atracción a mayor distancia, principalmente promovida por una mayor probabilidad de supervivencia del ataque de depredadores.
Con base en los resultados es posible proponer estrategias de reforestación y restauración forestal que consideren las interacciones positivas de los ensambles ecológicos formados por latizales, brinzales y fustales de las distintas categorías de altura y gradientes altitudinales, a fin de conservar la estructura y composición espacial de la Selva Lacandona.

