











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Por su alta fragilidad, la estructura es uno de los aspectos más relevantes en el ámbito forestal, lo que repercute en una fácil modificación de la misma. Usualmente, está referida a la forma en que los atributos de los árboles se distribuyen en los ecosistemas (Bettinger et al., 2008; Gadow et al., 2011). La importancia de la estructura biológica es su auto organización, la cual involucra a la regeneración, el crecimiento y a la mortalidad de los árboles; así como, a una variedad de interacciones entre individuos que, a su vez, afecta las propiedades del ecosistema (João y Carvalho, 2011; Gadow et al., 2011). Estas incluyen la producción de biomasa, el almacenamiento de carbono, la diversidad vegetal, la captura de agua, entre otros; y la calidad de esos servicios ecosistémicos dependen, en mayor medida, del manejo que se realice para llegar a un fin determinado (Ruiqiang et al., 2014). Por ello, es necesario realizar una caracterización correcta de la estructura de los ecosistemas que fundamente una toma de decisiones adecuada para la gestión de los recursos forestales (Jiménez et al., 2001; Wehenkel et al., 2011; Bannister y Donoso, 2013), con la finalidad de incrementar el contenido de carbono que un bosque puede almacenar, sin deteriorar la diversidad de la vegetación.
El manejo del bosque para la producción maderable no se contrapone con el dirigido para incrementar los almacenes de carbono y mucho menos, si al aumentar el carbono almacenado se incide en la pérdida de la diversidad del estrato arbóreo.
La caracterización de estos ecosistemas se puede realizar mediante el uso de índices que midan elementos de su diversidad, los cuales son indicadores de la calidad y cantidad de los recursos ecosistémicos (Magurran, 1988). Además, permiten comparar comunidades naturales o modificadas por efecto del manejo forestal (Halffter, 1998). Con lo anterior, es posible diseñar estrategias de conservación y de manejo sustentable a escala local (Moreno, 2001).Al respecto, la hipótesis planteada se refiere a que hay una relación entre la diversidad horizontal y vertical de la vegetación con el contenido de carbono, ya que al aumentar estas, se incrementan tanto la superficie, como los estratos existentes en un bosque, lo que hace eficiente el almacén de carbono. Por tal motivo, el objetivo fue evaluar la estructura de un ecosistema forestal y generar un modelo que describa el comportamiento del contenido de carbono con relación a la diversidad de la vegetación, a través de índices de diversidad, en un bosque en el noreste de México; con la finalidad de tener una herramienta nueva para la adecuada toma de decisiones, con respecto al manejo forestal.
Materiales y Métodos
El área de estudio se localiza en el sur del estado de Nuevo León, localizado geográficamente entre los 23°45’-25°32’ de latitud norte y 99°27’-100°25’ de longitud oeste (Figura 1). El clima es un semifrío subhúmedo con lluvias en verano [C(E)(w)], con temperatura media anual de 16 a 18 °C, precipitación media anual de 400 a 1 200 mm (García, 2004). La vegetación de la región corresponde a bosques conformados por asociaciones de pino-encino, encino-pino y pequeñas zonas con presencia de los géneros Abies, Pseudotsuga y Picea (Inegi, 2016).
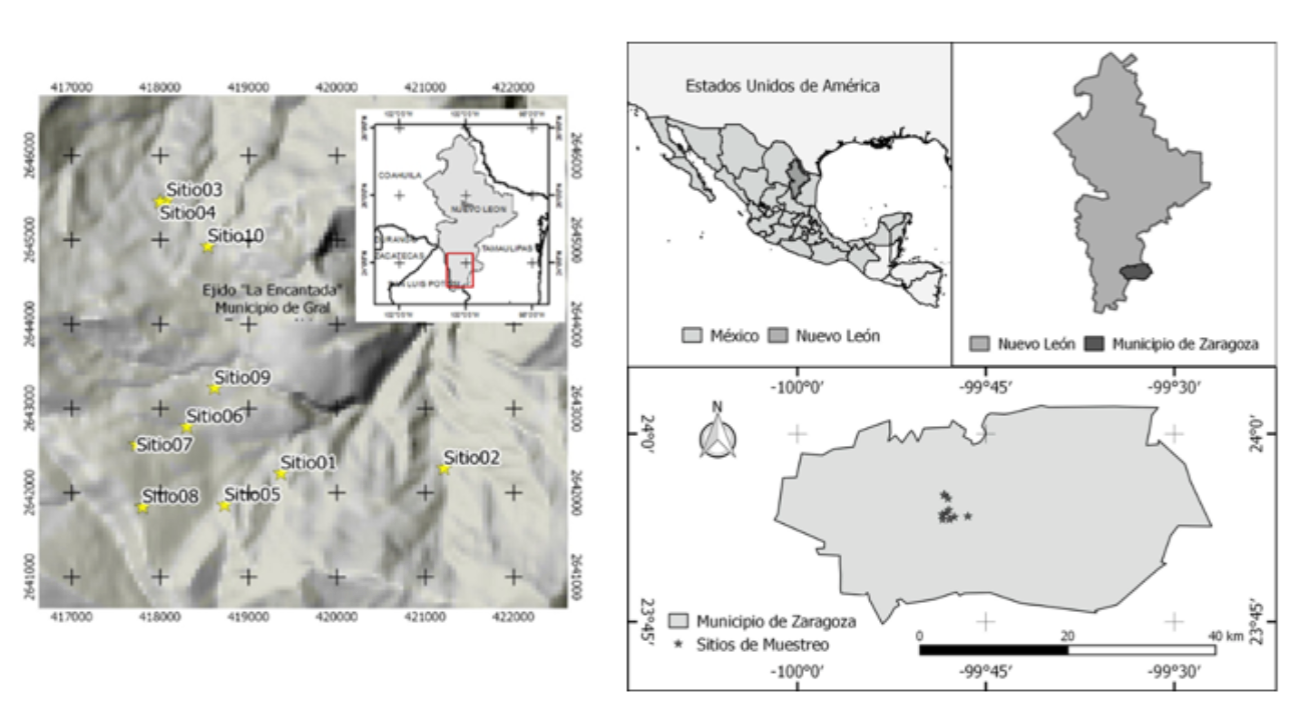
Se establecieron 10 sitios de muestreo de 2 500 m2 cada uno (50 × 50 m), ubicados geográficamente con un GPS Garmin ® 62 en diferentes condiciones fisiográficas (Cuadro 1), conforme a los parámetros utilizados en los sitios de investigación forestal y de suelo (Corral-Rivas et al., 2009). Para medir la pendiente de los sitios se empleó un clinómetro electrónico (Haglöf ®); asimismo, se utilizó una brújula Brunton ® para definir la orientación. En todos los sitios se midieron el diámetro normal (d 1.30 ), con una cinta diamétrica Forestry Suppliers Inc ®; la altura total (h), con un hipsómetro Vertez III ®, para cada uno de los árboles > 7.5 cm de diámetro normal. La evaluación de la similitud entre los sitios de muestreo se hizo en función de las características fisiográficas; para ello, se realizó un análisis de conglomerados (cluster) jerárquico, mediante la medida de distancia Bloque y el método de agrupación fue el de Ward’s con su respectivo dendrograma de similaridad (IBM Corporation, 2013).
Cuadro 1 Características fisiográficas de los sitios de muestreo.
| Sitio | UTM-X | UTM-Y | Altitud (msnm) | Exposición | Pendiente (%) |
|---|---|---|---|---|---|
| 1 | 419 365 | 2 642 230 | 2 600-2 700 | Noroeste | 0-20 |
| 2 | 421 211 | 2 642 230 | < 2 500 | Norte | > 100 |
| 3 | 418 074 | 2 642 783 | 2 500-2 600 | Norte | > 100 |
| 4 | 417 997 | 2 645 453 | 2 500-2 600 | Noreste | 0-20 |
| 5 | 418 729 | 2 641 853 | 2 700-2 800 | Noroeste | 0-20 |
| 6 | 418 301 | 2 642 783 | 2 700-2 800 | Noreste | 20-40 |
| 7 | 417 732 | 2 642 572 | > 2 900 | Este | 20-40 |
| 8 | 417 801 | 2 641 831 | > 2 900 | Este | 0-20 |
| 9 | 418 615 | 2 643 242 | 2 700-2 800 | Este | 20-40 |
| 10 | 418 540 | 2 644 924 | 2 600-2 700 | Noreste | 40-60 |
Para cada sitio se calculó la altura y el diámetro promedio, el área basal (G), el contenido de carbono (CC), y para cada una obtuvo la dispersión de datos por medio de la desviación estándar a nivel de sitio de muestreo. También se determinaron los índices de diversidad de especies horizontal (índice de Shannon-Wiener), el de Pretzsch para la diversidad vertical y el de valor de importancia (IVI).
El área basal se estimó como la suma por unidad de superficie de todos los fustes, a nivel del diámetro normal (Ecuación 1).
Donde:
G = Área basal
d = Diámetro normal
π = Constante (3.1416)
n = Número de árboles presentes dentro del sitio de muestreo
La cuantificación de carbono almacenado se realizó con las ecuaciones alométricas de las especies más importantes, generadas previamente por diferentes autore/s; para aquéllas que no se tuvieron expresiones específicas en la región de estudio, se usaron las desarrolladas en otras zonas de clima templado-frío del país para el taxón de interés (Cuadro 2).
Cuadro 2 Ecuaciones alométricas por especie.
| Especie | Ecuación | Autor |
|---|---|---|
| Picea martinezii T. F. Patterson |
|
Zianis et al. (2005) |
| Abies vejarii Martínez |
|
Avendaño et al. (2009) |
| Pseudotsuga menziesii (Mirb.) Franco |
|
Návar (2009) |
| Cupressus arizonica Greene |
|
Vigil (2010) |
| Pinus ayacahuite Ehrenb. ex Schltdl. |
|
Návar (2009) |
| Pinus teocote Schiede ex Schltdl et Cham. |
|
Aguirre-Calderón y Jiménez-Pérez (2011) |
| Pinus pseudostrobus Lindl. |
|
Aguirre-Calderón y Jiménez-Pérez (2011) |
| Taxus globosa Schltdl |
|
Volumen de un cono |
| Quercus spp |
|
Tomás (2013) |
| Otras hojosas |
|
Probosque (1990) |
V = Volumen (m3 ha-1); BA = Biomasa aérea (Mg ha-1); CC = Carbono capturado (Mg ha-1); D = Diámetro Altura de Pecho (m); H = Altura total (m); R = Radio (D/2).
La mitad de las especies arbóreas tuvieron ecuaciones alométricas para obtener el CC; para las restantes se aplicaron las usadas para estimar biomasa multiplicada por un factor (0.50) (IPCC, 2016). En el caso de Picea martinezii T. F. Patterson, Taxus globosa Schltdl. y las hojosas solo se tienen ecuaciones de volumen; por ello, se multiplicó por el factor 0.50 para obtener la biomasa aproximada (Silva-Arredondo y Návar-Cháidez, 2009). Los encinos y otras latifoliadas se agruparon para emplear una sola ecuación. Tanto para el área basal, como para el contenido de carbono se realizaron los cálculos por sitio de muestreo y después se efectuó la extrapolación a una hectárea, esto para hacer inferencias y comparaciones con otros estudios.
El índice de valor de importancia (IVI) es un índice sintético estructural, desarrollado principalmente para jerarquizar la dominancia de cada especie en rodales mezclados; se estimó sumando la abundancia relativa, la densidad relativa y la frecuencia relativa (Curtis y McIntosh, 1951) (Ecuación 2):
Donde:
A rel = Abundancia relativa
D rel = Densidad relativa
F rel = Frecuencia relativa.
Para la estimación de la abundancia relativa se utilizaron las ecuaciones 3 y 4:
Donde:
A i = Abundancia absoluta
N i = Número de individuos de la especie i
S = Superficie de muestreo (ha)
A rel = Abundancia relativa de la especie i respecto a la abundancia total
La dominancia se evaluó con las ecuaciones 5 y 6:
Donde:
D i = Dominancia absoluta
Ab i = Área basal de la especie i
S = Superficie muestreada (ha).
D rel = Dominancia relativa de la especie i respecto a la dominancia total
La frecuencia relativa se estimó con las ecuaciones 7 y 8:
Donde
F i = Frecuencia absoluta (porcentaje de presencia en los sitios de muestreo)
f i = Número de sitios en los que está presente la especie i
S = Superficie muestreada (ha)
N = Número de sitios de muestreo
F rel = Frecuencia relativa de la especie i respecto a la frecuencia total
El índice de diversidad horizontal de especies se define como el número de taxones en una unidad de área, tiene dos componentes principales: la riqueza (número de especies) y la equitatividad (número de individuos de una sola especie). Uno de los más usados es el Índice de Shannon-Wiener (H’) (Shannon y Weaver, 1949) (Ecuación 9):
Donde:
S = Número de especies presentes
P i = Proporción de las especies
p i = n i /N
n i = Número de individuos de la especie i
N = número total de individuos
La distribución vertical de las especies se calculó mediante el Índice de Pretzsch (A) (Pretzsch, 2009), el cual usa diferentes intervalos de altura para la detección de cambios en la diversidad en los distintos estratos del bosque (Ecuación 10).
Donde:
S = Número de especies presentes
Z = Número de estratos de altura
p ij = Porcentaje de especies en cada zona
p ij = n ij /N
n ij = Número de individuos de la especie i en el estrato j
N = Número total de individuos
Posteriormente, se generó un modelo estadístico que describe la relación entre ellos. Los análisis se hicieron con el paquete estadístico SPSS® ver. 22.0 (IBM Corporation, 2013). Se aplicó un análisis de varianza de un factor y se realizó una comparación de medias mediante la prueba de Tukey para determinar la existencia de diferencias significativas (p<0.05) (Zar, 2010).
Resultados y Discusión
Con base en el dendograma de similaridad, se observaron cinco grupos (Figura 2); los más diferentes (superior a 80 %) fueron los sitios 2 y 3, 1 y 5. El primer grupo se caracterizó por presentar una exposición norte, con pendientes por arriba de 100 %, un intervalo altitudinal de 2 500 a 2 600 m; al segundo grupo le correspondió una exposición noreste y pendientes de 0 a 20 %, con una altitud de 2 600 a 2 800 m.

Un tercer grupo se integró con los sitios con mayor similitud (menores a 30 %), los cuales tuvieron exposición hacia el este, con una altitud superior a 2 900 m y una pendiente de 0 a 40 %; aunque fueron discriminados los sitios 7 y 8.
Los sitios 6 y 9 conformaron el cuarto grupo, que se ubicó en un gradiente altitudinal de 2 700 a 2 900 m, con una pendiente de 20 a 40 %. Por último, un quinto agrupamiento, se conformó con los sitios 4 y 10, cuya exposición fue hacia el noreste y su altura varío de 2 500 a 2 700 msnm.
Respecto al Índice de Valor de Importancia (IVI), el mayor peso específico fue para los géneros Pinus (sitios 5, 6, 7, 8, 9), Quercus (sitios 4, 10), Picea (sitio 1), Abies (sitio 2) y Cupressus (sitio 3). Los taxones con más importancia en la mayoría de los sitios muestreados fueron: Pinus teocote Schiede ex Schldl. et Cham. (sitios del 6 al 10) y Quercus cordifolia Trel. (8, 9 y 10) (Cuadro 3). Este comportamiento de la vegetación se debe a que son bosques mezclados de pino-encino.
Cuadro 3 Relación de especies por sitio de muestreo y su valor de IVI.
| Especie | Sitio | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | |
| Abies vejarii Martínez | 16.81 | 33.04 | 32.31 | 6.24 | 1.80 | |||||
| Arbutus xalapensis HBK. | 1.06 | 7.60 | 4.50 | 11.14 | 12.22 | 6.24 | 7.53 | 4.73 | ||
| Carpinus caroliana Walter | 1.08 | |||||||||
| Cupressus arizonica Greene | 34.17 | 0.71 | ||||||||
| Picea martinezii T. F. Patterson | 29.06 | 8.53 | ||||||||
| Pinus pseudostrobus Lindl. | 1.52 | 14.13 | 4.12 | 11.46 | ||||||
| Pinus ayacahuite Ehrenb. ex Schltdl. | 12.67 | 2.15 | 12.07 | 1.07 | 37.60 | 4.21 | 12.94 | |||
| Pinus teocote Schiede ex Schltdl et Cham. | 5.05 | 2.77 | 54.01 | 44.23 | 50.35 | 42.30 | 16.36 | |||
| Populus tremuloides Michx. | 3.45 | |||||||||
| Pseudotsuga menziesii (Mirb.) Franco | 9.83 | 1.61 | 9.10 | 1.74 | 4.14 | |||||
| Quercus affinis Scheidw. | 18.63 | 19.24 | 9.32 | 62.81 | 10.03 | |||||
| Quercus cordifolia Trel. | 43.41 | 41.85 | 50.36 | |||||||
| Quercus germana Schltdl. & Cham. | 8.92 | |||||||||
| Quercus greggii Trel. | 7.08 | |||||||||
| Quercus laeta Liebm. | 2.42 | 20.99 | ||||||||
| Quercus mexicana Humb. & Bonpl. | 15.68 | |||||||||
| Quercus polymorpha Cham. & Schltdl. | 6.96 | |||||||||
| Quercus sideroxyla Humb. & Bonpl. | 23.90 | 33.06 | 22.55 | |||||||
| Taxus globosa Schltdl. | 13.01 | 9.58 | 1.50 | |||||||
| Ulmus crassifolia Nutt. | 1.07 | |||||||||
En el sitio 2 se registró la cantidad más alta de especies, con 11, seguido del 4, con 10; ambos presentan exposiciones norte-noreste y un intervalo altitudinal de 2 500 a 2 700 m. El sitio con menos taxones fue el 8, en el que solo se identificaron tres y que se caracteriza por tener una exposición este y altitud superior a 2 900 m; los sitios 6 y 7 tuvieron cuatro especies; en los sitios restantes el número de taxa varió entre cinco y siete por sitio (Cuadro 3). Dicho comportamiento evidencia que a mayor altitud disminuye el número de especies por sitio (menor diversidad). En todos los sitios, al menos, hubo un taxon del género Pinus y uno de Quercus. Arbutus xalapensis se observó en ocho sitios; Pinus teocote (cuatro de ellos con el mayor valor de IVI) y P. ayacahuite en siete sitios. Las especies con menos presencia (una por sitio) fueron: Quercus germana, Q. greggii, Q. mexicana, Q. polymorpha, Carpinus caroliana y Ulmus sp.
La mayoría de las alturas promedio fluctuaron entre 11.2 y 14.2 m, excepto en los sitios 1 y 3, cuyos valores fueron de 16.9 a 20.5 m; estos últimos conformaron rodales frondosos, dominados por Cupresus arizonica y Picea martinezii en ubicaciones con exposición norte-noreste. Los diámetros promedio más altos (35.9 y 32.4 cm) se obtuvieron en los sitios 3 y 9, en los cuales dominan Cupressus arizonica y Pinus teocote, respectivamente; mientras que, al sitio 4 le correspondió el menor diámetro promedio, con 18.1 cm, la especie dominante fue Quercus affinis; su exposición es noreste, con una altitud menor a 2 700 m.
El área basal presentó diferencias significativas entre los 10 sitios (p=0.109); el 3 tuvo el valor más alto, con 49.01±7.74 m2 ha-1 y mayor presencia de Cupressus arizonica; y el menor fue para el sitio 8, con 22.67±2.5 m2 ha-1, integrado en su mayoría por Pinus teocote, con exposición este y una altitud >2 900 m. Estos cifras son similares a las citadas por Encinas-Domínguez et al. (2008) para un bosque de oyamel en Coahuila (29.69 m2 ha-1), pero menores a los documentados por Aguirre et al. (2003) para bosques mixtos en el noreste del país (35.9 a 48.2 m2 ha-1), aunque estos coinciden con los del sitio 3 (Cuadro 4).
Cuadro 4 Variables estructurales por sitio.
| Sitio | Altura promedio (m) | Diámetro promedio (cm) | G (m2 ha-1) | CC (Mg ha-1) | Índice de Pretzsch | Índice de Shannon |
|---|---|---|---|---|---|---|
| 1 | 16.96 ± 0.82 a | 23.94 ± 0.87 a | 23.20 ± 7.28 a | 83.90 ± 24.00a | 2.40 | 1.37 |
| 2 | 14.21 ± 0.68 a,b | 21.29 ± 1.70 a,b | 29.94 ± 9.29 a | 111.23 ± 37.25a | 2.53 | 1.78 |
| 3 | 20.57 ± 0.81 c | 35.96 ± 2.00 c | 49.01 ± 7.74 a | 123.49 ± 17.19a | 2.32 | 1.48 |
| 4 | 12.43 ± 0.44 b | 18.10 ± 0.47 b | 31.89 ± 2.03 a | 71.31 ± 5.10a | 2.01 | 1.23 |
| 5 | 13.02 ± 0.33 b | 24.27 ± 1.22 a,d | 33.44 ± 4.49 a | 94.70 ± 14.06a | 2.27 | 1.57 |
| 6 | 13.92 ± 0.59 a,b | 29.53 ± 1.04 d,e | 27.01 ± 6.57 a | 71.73 ± 18.83a | 1.50 | 0.95 |
| 7 | 12.10 ± 0.75 b | 24.61 ± 0.70 a,d | 33.22 ± 1.55 a | 91.81 ± 3.09a | 2.21 | 1.26 |
| 8 | 13.00 ± 0.96 b | 25.86 ± 1.17 a,d | 22.67 ± 2.50 a | 58.36 ± 5.75a | 1.61 | 0.83 |
| 9 | 13.42 ± 0.57 b | 32.45 ± 0.42 c,e | 28.68 ± 2.23 a | 82.95 ± 10.39a | 1.85 | 0.91 |
| 10 | 11.28 ± 0.42 b | 21.50 ± 0.89 a,b | 28.70 ± 4.85 a | 81.46 ± 14.81a | 2.27 | 1.36 |
G = Área basal; CC = Contenido de carbono. Medias seguidas de diferentes letras (a, b, c, d, e) indican niveles diferentes de significancia para p<0.05.
El carbono promedio en los 10 sitios presentó diferencias (p=0.354). En el sitio 3 se estimó la mayor cantidad de carbono capturado, con 123.48±17.19 Mg ha-1; seguido del 2, con 111.22±37.26 Mg ha-1; las especies representativas fueron Cupressus arizonica y Abies vejarii, respectivamente, además registraron el IVI más alto por sitio; se ubicaron en exposiciones norte, con pendientes superiores a 100 %. Autores como, Razo-Zárate et al. (2013) consignan valores de 62.6 Mg ha-1 y 166.6 Mg ha-1 para bosques similares, tanto conservados como perturbados, respectivamente. El sitio 8 tuvo el CC más bajo con 58.35±5.75 Mg ha-1, en el que dominó Pinus teocote, además registró el menor valor de área basal, se desarrollan en altitudes que superan los 2 900 msnm, con exposición este, y constituyen rodales con arbolado de dimensiones medianas, en comparación con los sitios 3 y 2 (Cuadro 4).
El índice de Shannon-Wiener en el sitio 2 fue de H’=1.78 y H’ max =2.4, que se consideran altos, con 11 especies, lo cual evidencia una gran diversidad; asimismo, resultó con el segundo mayor valor de contenido de carbono almacenado. En tanto que, los valores más bajos fueron para el sitio 8, con H’=0.83 y H’ max =1.1 (Figura 3a), con solo tres especies, pero los valores entre H’ y H’ max son similares, lo que corresponde a una distribución homogénea. Los resultados anteriores son superiores a los señalados por Santibañez-Andrade et al. (2015) en un bosque de pino del Valle de México (H’=1.02-1.28); mientras que, López-Hernández et al. (2017) indican valores de H’=1.37, en bosques templados bajo manejo forestal, cifras similares a los valores de los sitios 1, 3, 4, 7 y 10.
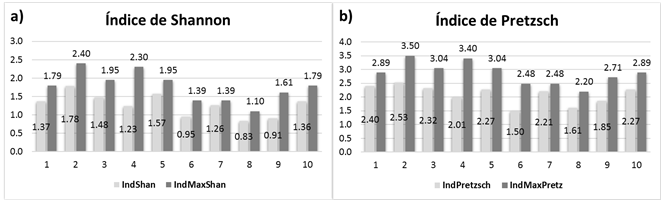
Figura 3 Gráficas de índice de diversidad y su máximo por sitio; a) índice horizontal (Shannon-Wiener), y b) índice vertical (Pretzsch).
Para la estructura vertical se calcularon los valores del índice A (Pretzsch), que representa la diversidad de especies por estrato vertical. El sitio 2 presentó el mayor valor de A=2.53 y A max =3.5; por lo que se considera un rodal no uniforme, respecto a las alturas. También tuvo los registros más altos en diversidad horizontal (índice H); por lo tanto, es un rodal con amplia diversidad tanto vertical, como horizontal con valores dasométricos altos (altura promedio, diámetro promedio, área basal y contenido de carbono), debido, principalmente, a su condición fisiográfica que corresponde a una altitud inferior a 2 500 m, la cual es menor a la del resto de los sitios estudiados. Para el sitio 6 se estimó el valor más bajo (A=1.5 y A max =2.48) (Figura 3b), aunque muy similar al sitio 8 (A=1.61 y A max =2.20), esto representó una uniformidad media que corresponde a las condiciones más pobres del bosque, en cuanto a diversidad arbórea, debido, sobre todo, a sus condiciones fisiográficas (pendiente de 20 a 40 % y altitud de 2 700 a 2 900 m).
Los valores altos en el índice de Pretzsch sugieren una gran diversidad de especies en la estructura vertical, como ocurrió en el sitio 2 con 10 especies, pero sin distribución uniforme en todos los estratos. Los valores del sitio 7, cuyos A y A max son muy similares, reflejan una distribución homogénea de los taxoness en los tres estratos. Estos resultados coinciden con los consignados por Rubio et al. (2014), con A max =3.74 y A=2.01 en bosques mixtos; pero son altos en comparación con los de Villavicencio et al. (2006), quienes obtuvieron: A=2 y A max =2.7 para un bosque de pino-encino, donde el encino dominó en los tres estratos.
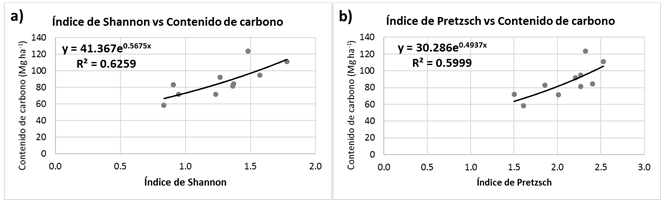
Para describir la relación del contenido de carbono con los índices de diversidad horizontal y vertical, se generaron varios modelos de regresión. En el caso del índice de Shannon-Wiener, el mejor modelo fue uno exponencial (y=41.367e0.5675x), con un R2=0.62 (Figura 4a); para el índice de Pretzsch, también fue un modelo exponencial (y=30.286e0.4937x), con un R2=0.59 (Figura 4b). Ambos presentaron R2 bajos, pero se observó una tendencia entre las variables estudiadas (Figura 4) en las que, a mayor valor en los índices de diversidad, el contenido de carbono aumenta. Estas relaciones positivas se deben a que existe una distribución homogénea de las especies arbóreas dentro de los sitios de estudio; además, se distribuyen en los tres estratos verticales evaluados cubriendo mayor cantidad de superficie tanto vertical, como horizontal.
Conclusiones
Se concluye que sí existen variaciones de composición y estructura dentro del ecosistema estudiado, el cual está compuesto, principalmente, por asociaciones de pino-encino. En general, está constituido por individuos de porte mediano (11 a 14 m de altura) y diámetros de 20-30 cm.
Se demostró que existe una relación entre carbono capturado y los índices de diversidad horizontal de Shannon-Wiener (H’) y el índice de diversidad vertical de Pretzsch (A). Los modelos generados, aunque tienen una R2 baja (0.59 para H’ y 0.62 para A), sí reflejan la relación entre el contenido de carbono y la diversidad de especies arbóreas, tanto horizontal como vertical y evidencian que a medida que el valor de los índices aumenta, el CC se incrementa; por lo tanto, una las estrategias a seguir en los planes de manejo sería tener especies distribuidas uniformemente en toda el área y en los diferentes estratos verticales.

