











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
El bosque mesófilo de montaña (BMM) es uno de los ecosistemas que alberga la mayor diversidad de especies de flora y fauna, aunque cubre una extensión menor a 1 % del territorio nacional (Challenger, 1998). Se calcula que de 2 500 a 3 000 taxa de plantas vasculares habitan exclusiva y preferentemente en este tipo de vegetación (Rzedowski, 1993). Su distribución coincide con un clima templado, de humedad alta, en el que la precipitación y la niebla se presentan de manera continua o con mucha frecuencia, de ahí que también se le conozca como bosque de niebla o bosque húmedo de montaña (Rzedowski, 1978, Villaseñor, 2010).
Un componente importante del BMM son las plantas epífitas, las cuales crecen sobre otros vegetales sin obtener de ellos sus nutrientes, por lo que el hospedante o forofito solo tiene la función de soporte (Benzing, 1990). Las epífitas contribuyen significativamente, a la biodiversidad de los trópicos, ya que constituyen hasta 40 % de la flora de una zona tropical, y 10 % de todas las plantas vasculares del mundo (Gentry y Dodson, 1987). Proveen hábitat y alimento a muchos invertebrados y aves (Barthlott et al., 2001) y participan, en forma activa en la dinámica de los nutrimentos y el agua retenidos en el dosel, los cuales son reincorporados al medio a través del escurrimiento de agua o por la caída y muerte de epifitas (Nadkarni y Matelson, 1991).
Aunque se han reconocido 800 especies de epífitas en los BMM de México, solo en algunos se conoce con detalle la composición florística de estas plantas y su contribución a la flora total (Sánchez-Ramos y Dirzo, 2014). Lo anterior se debe, principalmente, a la dificultad que implica en sitios, por lo regular inaccesibles (Ingram y Lowman, 1995), si bien en los últimos años nuevos métodos de ascenso han mejorado esa situación.
De cualquier manera, el conocimiento de la distribución de las epífitas con base en condiciones bióticas y abióticas es importante para entender los patrones de diversidad en distintos ecosistemas, especialmente si están amenazados, como es el caso de BMM (Gentry y Dodson, 1987). En el presente trabajo se determinó la composición de especies epífitas en uno de los puntos de distribución más boreales del bosque de niebla. Se enfatiza la posible asociación entre epífitas y forofitos, así como la preferencia de las primeras por crecer en secciones del tronco o la copa de su árbol hospedero y la relación entre la distribución de especies y su filogenia.
Materiales y Métodos
Área de estudio
La Reserva de la Biósfera El Cielo se localiza en el suroeste del estado de Tamaulipas, en las estribaciones de la Sierra Madre Oriental. Está limitada por los paralelos 23°12´y 23°03´N, el meridiano 99°18´ O (Puig y Bracho, 1987). De acuerdo con la Conabio (2010), El Cielo incluye los fragmentos de distribución más septentrional de BMM en el continente americano. En la reserva se ubica la porción más extensa de BMM en Tamaulipas (aproximadamente 16 142.86 ha), además presenta un alto grado de conservación (Sánchez-Ramos et al., 2005).
Los límites altitudinales del bosque mesófilo de montaña en El Cielo son de 800 y 1 500 m, que comprenden un área aproximada de 100 km2. Los suelos predominantes son litosoles (negros), rendzinas (negras) y luvisoles crómicos (negros) (Puig y Bracho, 1987). El clima semicálido subhúmedo, corresponde a la mayor parte del bosque mesófilo; las precipitaciones son mayores a 2 000 mm anuales, y se ubican en alturas entre 700 y 1 400 msnm (Casas y Requena, 2005).
En los lugares sin disturbio se encuentra un bosque denso en el estrato arbóreo, el que a su vez se diferencia de acuerdo a la altura en cuatro subestratos. El dosel, tiene una altura entre 18 y 30 m, está compuesto por Liquidambar styraciflua L., Clethra pringlei S. Watson, Quercus germana Schltdl. & Cham. y Quercus sartorii Liebm., además de Fagus mexicana Martínez en algunas áreas. Entre 12 y 18 m se distribuyen otras especies, como Carya ovata (Mill.) K. Koch, Magnolia tamaulipana A. Vázquez, Ostrya virginiana (Mill.) K. Koch, Podocarpus reichei J. Buchholz & N.E. Gray y Tilia houghi Rose. En un tercer subestrato (8 a 12 m), se desarrollan Meliosma oaxacana Standl. y Turpinia occidentalis (Sw.) G. Don; y en el cuarto Eugenia capuli (Schltdl. & Cham.) Hook. & Arn., Rapanea myricoides (Schltdl.) Lundell y Ternstroemia sylvatica Schltdl. & Cham. (Puig et al., 1983).
Selección de sitios y muestreo
El trabajo de campo se inició en abril de 2011, con salidas mensuales de una semana, en un periodo de seis meses. Se seleccionaron sitios ubicados en las cotas altitudinales de 800, 1 000, 1 300 y 1 500 msnm, de acuerdo a la distribución del bosque mesófilo de montaña en la Reserva (Puig y Bracho, 1987).
Con base en la metodología propuesta por Gradstein et al. (2003), en cada cota altitudinal se escogieron cinco árboles maduros, con una distancia mínima de 25 m entre cada individuo. Se eligieron los de mayor tamaño, porque se presume que pueden ser los más viejos, y por tanto, estar sujetos a mayor colonización por epífitas (Gradstein et al., 1996).
Alrededor de cada árbol maduro se delimitó un cuadrante de 20 × 20 m, con el fin de muestrear los ejemplares vecinos con presencia de epífitas. Se registró la ubicación de cada individuo arbóreo muestreado, mediante un GPS Garmin e-trex y se midió, tanto su diámetro a la altura del pecho (DAP), con una cinta diamétrica de 10 m Forestry suppliers; y su altura, con un clinómetro PM5/360PC Suunto.
Para el registro de las especies epífitas, el árbol muestreado (tanto maduros como los de los cuadrantes) fue subdividido en las cinco zonas propuestas por Johansson (1974) (Figura 1). Se consideraron las epífitas por individuo arbóreo, desde la base del tronco hasta las ramas exteriores.
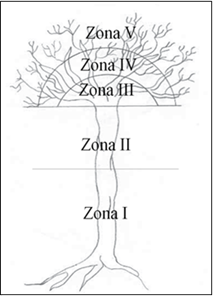
Fuente: Bøgh, 1992.
Zona 1 = 0.22-3 m a partir del suelo; Zona 2 = 3 m-primeras ramas mayores; Zona 3 = Ramas grandes; Zona 4 = Ramas de talla media y delgadas; Zona 5 = Las ramas más delgadas.
Figura 1 Zonificación del árbol hospedero (Johansson, 1974).
Para el acceso al dosel, se usó equipo de escalar, arnés, martillos, ascensores, estribos, mosquetones y dispositivo de descenso (Benzing, 1990). Además, se hicieron observaciones desde el suelo, con ayuda de binoculares (Eagle Optics Ranger ED). Se muestreó un total de 188 árboles de 24 especies, 20 géneros y 17 familias: 40 en la cota de 1 500 msnm, 50 a 1 300 msnm, 46 a 1 000 msnm y 52 a 800 msnm.
Se incluyeron en el muestreo las especies de epífitas vasculares, tanto ejemplares adultos como juveniles. Se tomaron como unidades vegetativas: individuos, rosetas y láminas, (Hietz y Hietz-Seifert, 1995). Para la primera categoría se consideró el género Peperomia; para la segunda, la mayor parte de los taxones de bromeliáceas; y para la tercera, los helechos y la mayor parte de los géneros de orquídeas que forman grandes agregados, en los que no es confiable distinguir un individuo de otro. Se colectaron ejemplares de todos los taxa, los cuales se herborizaron, identificaron y depositaron en el herbario de la Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León.
Análisis estadístico
La diversidad de las epífitas se obtuvo con el índice de Shannon. Se elaboraron fenogramas para conocer el grado de similitud que hay entre las zonas del árbol, de acuerdo a las especies epífitas contenidas. Estos proporcionan además el valor del índice de similitud de Sørensen. Se aplicó una prueba de χ2 para determinar la preferencia de las epífitas hacia los hospederos, así como para conocer la asociación entre las epífitas y cada una de las zonas del árbol propuestas por Johansson (1974). La distancia filogenética entre las especies, medida en millones de años desde un ancestro en común se detectó con Timetree (Hedges et al. 2006), y las diferencias entre distancias filogenéticas fueron analizadas con un ANOVA.
Resultados
Abundancia y riqueza de especies
Se identificaron 10 familias, 23 géneros y 33 especies de epífitas (Cuadro 1). El total de superficie muestreada fue 0.8 ha (8 000 m2). La familia Orchidaceae representó 43 % del total de taxones, seguida de Polypodiaceae (15 %) y Bromeliaceae (12 %).
Cuadro 1 Inventario florístico de epífitas identificadas en el bosque mesófilo de montaña de la Reserva de la Biósfera El Cielo.
| Angiospermas | |||
|---|---|---|---|
| Familia | Especie | Altitud (msnm) | Zonas del forofito |
| Agavaceae | Agave celsii Hook. | 800, 1 500 | 1-3 |
| Begoniaceae | Begonia sp. | 1 300 | 1 |
| Bromeliaceae | Tillandsia bartramii Elliott | 800-1 500 | 2-5 |
| Tillandsia deppeana Steud. | 800-1 500 | 1-5 | |
| Tillandsia utriculata L. | 800, 1 000, 1 500 | 2-5 | |
| Tillandsia sp. | 1 300, 1 500 | 1,3,4 | |
| Cactaceae | Rhipsalis baccifera (Sol.) Stearn | 800 | 1-4 |
| Selenicereus sp. | 800 | 1,2 | |
| Crassulaceae | Sedum sp. | 1 300 | 3 |
| Orchidaceae | Arpophyllum laxiflorum Pfitzer | 1 500 | 4 |
| Catasetum integerrimum Hook. | 800, 1 000 | 1-4 | |
| Desconocida 1 | 1 500 | 2,4 | |
| Desconocida 2 | 1 500 | 2 | |
| Encyclia alata (Bateman) Schltr. | 800, 1 000 | 1-3 | |
| Encyclia parviflora (Regel) Withner | 800 | 1-4 | |
| Isochilus unilateralis B.L. Rob. | 800-1 500 | 1-5 | |
| Laelia anceps Lindl. | 1 300 | 3 | |
| Oncidium sphacelatum Lindl. | 800 | 1-4 | |
| Prosthechea cochleata (L.) W.E. Higgins | 800 | 1-4 | |
| Prosthechea lívida (Lindl.) W.E. Higgins | 800 | 2-4 | |
| Stanhopea tigrina Bateman ex Lindl. | 800-1 500 | 1-4 | |
| Trichocentrum ascendens (Lindl.) M.W. Chase & N.H. Williams | 800 | 1-4 | |
| Trichocentrum cosymbephorum (C. Morren) R. Jiménez & Carnevali | 800 | 1-3 | |
| Piperaceae | Peperomia quadrifolia (L.) Kunth | 800-1 500 | 1-4 |
| Peperomia sp. | 800 | 1,2 | |
| Pteridofitas | |||
| Lycopodiaceae | Huperzia dichotoma (Jacq.) Trevis. | 800-1 500 | 1-4 |
| Aspleniaceae | Asplenium cuspidatum Lam. | 800, 1 300-1 500 | 1-5 |
| Asplenium sp. | 1 300 | 2-5 | |
| Polypodiaceae | Campyloneurum angustifolium (Sw.) Fée | 800-1 500 | 1-5 |
| Pecluma alfredii (Rosenst.) M.G. Price | 800, 1 000 | 1-4 | |
| Pleopeltis polylepis (Roem. ex Kunze) T. Moore | 800-1 500 | 1-5 | |
| Polypodium polypodioides (L.) Watt | 800-1 500 | 1-5 | |
| Polypodium sp. | 1 300 | 1-4 | |
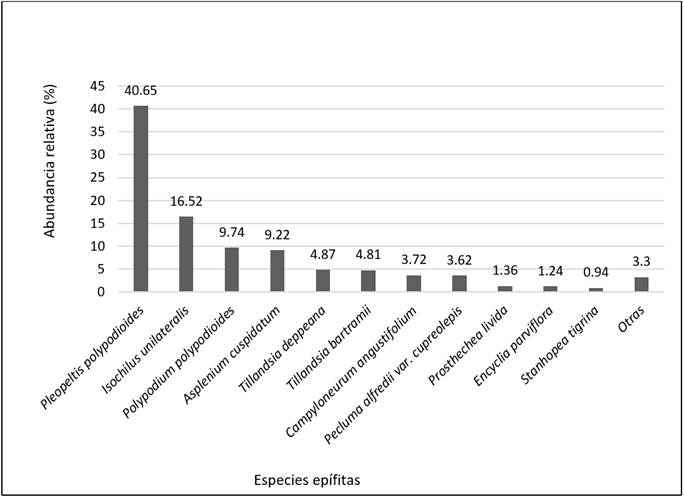
Se registraron 115 069 unidades vegetativas. Seis especies acaparan el mayor porcentaje de abundancia de individuos: Pleopeltis polylepis (Roem. ex Kunze) T. Moore (Polypodiaceae, 40.65 %), Isochilus unilateralis B.L. Rob. (Orchidaceae, 16.52 %); Polypodium polypodioides (L.) Watt (Polypodiaceae, 9.74 %); Asplenium cuspidatum Lam. (Aspleniaceae, 9.22 %), Tillandsia deppeana Steud. (Bromeliaceae, 4.87 %); y Tillandsia bartramii Elliott (Bromeliaceae, 4.81 %). El resto presentó una baja abundancia, menor a 1 % (Figura 2).
Quercus germana fue el forofito con el porcentaje de epífitas más alto (38.08 %), seguido de Q. sartorii (21.61 %) y Clethra pringlei (13.31 %).
Asociación epífita-forofito
En el Cuadro 2 se muestran los resultados de la prueba χ2. Las epífitas asociadas con forofitos no fueron filogenéticamente más distantes de los forofitos que las epífitas no asociadas (F=0.51, P=0.43).
Cuadro2 Valores de χ2 que muestran asociación entre especies epífitas más frecuentes y forofitos.
| Epífita/Forofito | Pleopeltis polylepis | Isochilus unilateralis | Polypodium polypodioides | Asplenium cuspidatum | Tillandsia bartramii | Campyloneurum angustifolium | Pecluma alfredii var. cupreolepis | Sedum sp. | Arpophyllum laxiflorum |
|---|---|---|---|---|---|---|---|---|---|
| Quercus germana | N.S. | 20.1* | 6.82* | N.S. | 5.36* | 25* | N.S. | N.S. | N.S. |
| Quercus sartorii | N.S. | N.S. | N.S. | N.S. | 6.99* | N.S. | N.S. | N.S. | N.S. |
| Clethra pringlei | N.S. | N.S. | N.S. | N.S. | N.S. | 4.95* | N.S. | N.S. | N.S. |
| Quercus canbyi | N.S. | N.S. | N.S. | 5.72* | N.S. | N.S. | N.S. | N.S. | N.S. |
| Sapindus saponaria | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 6.78* | N.S. | N.S. |
| Liquidambar styraciflua | 7.9* | N.S. | 5.21* | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. |
| Decatropis bicolor | N.S. | N.S. | N.S. | N.S. | N.S. | 0.36 | 4.8* | N.S. | N.S. |
| Turpinia occidentalis | 2.86 | N.S. | N.S. | 2.94 | N.S. | 6.35* | N.S. | N.S. | N.S. |
| Rapanea myricoides | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 14.3 | 18.8 |
| Quercus affinis | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | N.S. | 6.2 |
χ2 tabulada = 3.84, p< 0.05); N.S. = No significativas; * = Diferencias significativas (p<0.05).
Similitud entre forofitos por carga de epífitas
El fenograma para analizar la similitud entre especies arbóreas, respecto a las epífitas que albergan (Figura 3) evidencia que las especies con mayor similitud fueron: 1) Nectandra sanguinea Rol. ex Rottb., Meliosma oaxacana y Abies vejarii Martínez (Índice de Similitud= 1.0); 2) Quercus germana y Quercus sartorii (Índice de Similitud= 0.945); 3) Podocarpus reichei y Magnolia tamaulipana (Índice de Similitud= 0.880); y 4) Turpinia occidentalis y Rapanea myricoides (Índice de Similitud= 0.875).

Proporción de especies por familias en cotas altitudinales
A 1 500 msnm se identificaron 15 especies y seis familias, Orchidaceae presentó mayor proporción de especies (33 %,), seguida de Bromeliaceae (26 %) y Polypodiaceae (20 %). A 1 300 msnm ocurrieron 16 especies y siete familias, sobresalieron Polypodiaceae (29 %), Orchidaceae y Bromeliaceae (18 % y 17 %, respectivamente). Dos familias se restringen a este piso altitudinal: Crassulaceae y Begoniaceae, aunque su representación dentro de los cuadrantes es de pocos individuos. En la cota de 1 000 msnm se registraron 12 especies y cinco familias, Orchidaceae fue la mejor representada (34 %), seguida de Bromeliaceae y Polypodiaceae, ambas con 25 %. A 800 msnm se presentaron 24 especies y ocho familias, de las cuales Orchidaceae tuvo la mayor proporción de especies (41.67 % del total), Polypodiaceae (16.67 %) y Bromeliaceae (12.50 %). El número más alto de epífitas se obtuvo a 800 msnm, lo cual coincide con el Índice de Shannon por cota altitudinal (Cuadro 3). La distancia filogenética entre especies epífitas fue similar dentro y entre altitudes (F= 1.41, P=0.18).
Distribución vertical de epífitas
La distribución vertical de las epífitas en cada una de las secciones del forofito, muestra su marcada preferencia hacia las zonas III y IV del hospedante, de acuerdo a la clasificación de Johansson (Figura 4; χ2= 31 700.00, p<0.001), sitios que corresponden a la parte basal y media de las ramas; hay más epífitas de las que se esperaría en una distribución aleatoria.
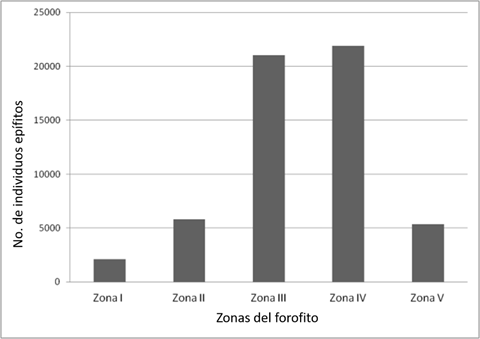
Hubo diferencias en la distribución por zonas de todas las especies, cuyos valores de χ2 calculada fueron superiores a los de χ2 tabulada (χ2= 9.48, p= 0.05). Esto significa, que las epífitas, aun las que ocupan las cinco zonas del forofito, son más comunes en una o dos de ellas y es ahí donde se registró la mayor abundancia.
Discusión
Abundancia y riqueza de especies
La predominancia de las familias Orchidaceae, Polypodiaceae y Bromeliaceae coincide con la clasificación hecha por Gentry y Dodson (1987) de estos grupos taxonómicos, con respecto al número total de especies epífitas. Cabe mencionar que Orchidaceae, aunque tiene el número más alto de taxones, su abundancia es baja. Isochilus unilateralis (Orchidaceae) le correspondió el valor más grande en el fragmento de bosque de niebla estudiado, quizás por el pequeño espacio que ocupa. El patrón de distribución de esta familia concuerda con lo citado en la literatura, respecto a que las orquídeas registran una alta diversidad de especies, aunque la característica principal de muchas de ellas es su escasez de individuos (Nieder et al., 1999; Valverde, 2006).
Destaca pos su abundancia Pleopeltis polylepis, lo cual se explica por su tendencia a formar grandes conglomerados, que en ocasiones cubren casi por completo las ramas de los forofitos, sin permitir el crecimiento de otras especies en la vecindad.
Asociación epífita-forofito
La principal función del forofito es la de proveer soporte a las epífitas. Si su función se restringiera a ello, la distribución de estas plantas sería generalista, independientemente de la especie arbórea que la sustenta. Algunas, se correlacionan con los taxa arbóreos, mediante la prueba de χ2. Es decir, existen características de los forofitos que inciden en la distribución y abundancia de las epífitas (Callawell et al., 2002; Krömer Gradstein, 2003). Dicha correlación se determinó tanto en las especies más abundantes, como en las más escasas, ejemplos de las primeras son: Isochilus unilateralis, Campyloneurum angustifolium (Sw.) Fée, Polypodium polypodioides y Tillandsia bartramii; y de las segundas: Sedum sp., Laelia anceps Lindl., Begonia sp., Arpophyllum laxiflorum Pfitzer y Piper sp. Sin embargo, la rareza de esas especies epífitas, también puede ser el factor que incide en los valores de χ2, para que sugieran una correlación con el forofito. Incrementar el número de colectas ayudaría a ampliar el último punto planteado.
Elementos como el tamaño y la forma del árbol determinan la abundancia y riqueza de especies epífitas. La predominancia de Quercus germana como forofito preferido se debe a su abundancia y fisonomía, pues tienen un porte muy ramificado y una mayor área basal, como lo registraron de la Rosa-Manzano et al. (2017) en un bosque de pino-ecino en la RBC. Además, la orientación casi horizontal de sus ramas propicia la acumulación de materia orgánica y la conservación prolongada de la humedad ambiental, lo que es idóneo para el establecimiento de muchas epífitas (Puig, 1993).
Otras especies arbóreas muy comunes en el bosque mesófilo de montaña, como Liquidambar styraciflua, contienen pocas plantas de ese tipo, a pesar de que presentan corteza rugosa. Sin embargo, su fuste es casi recto, sus ramificaciones son delgadas y su cobertura de copa es pequeña, por lo que ofrece poca superficie de fijación para las epífitas (Puig, 1993).
Distribución vertical
En varios trabajos se menciona que las epífitas tienden a ser más abundantes en las zonas III y IV de los forofitos (Nieder et al., 1999; Krömer et al., 2007), como se observó en el presente estudio. De acuerdo con Nieder et al. (1999), las preferencias en altura corresponden a adaptaciones de tipo fisiológico y mecánico. Así, por ejemplo, el género Campyloneurum es más abundante en las primeras dos zonas del forofito, lo que refleja su adaptación a un medio sombrío. En tanto que, Pleopeltis se distribuye en las cinco zonas del forofito, aunque es más abundante en las zonas III y IV. También en esos sitios se concentra gran parte de las orquídeas, con excepción de Isochilus unilateralis, la cual puede encontrarse en la zona V, pero en menor número, debido principalmente a la alta insolación y la rápida desecación imperante en esa zona.
Comparación con estudios similares en bosques de montaña neotropicales
El total de taxa identificados es menor a lo documentado en estudios realizados en otro bosque mesófilo de montaña. Martínez et al. (2006) registran 44 especies en un área de 0.04 ha en Chiapas; Hietz y Hietz-Seifert (1995) citan 53 a una altitud de 1 439 msnm, dentro de un área de 0.06 ha, con un total de 134 taxa para todos sus sitios de muestreo. En el presente se consignan muestra 33 especies en 0.6 ha.
En latitudes más al sur, como Ecuador y Bolivia, el número de especies aumenta, notablemente (Krömer y Gradstein, 2003). El alto número y diversidad en las zonas ecuatoriales se explica a partir de los múltiples nichos y microclimas derivados de la orografía (Küper et al., 2004). El menor número de taxones puede explicarse con el hecho de que conforme aumenta el gradiente latitudinal, la diversidad de epífitas decrece (Gentry y Dodson, 1987).
Dado que la Reserva de la Biósfera El Cielo es uno de los sitios que marcan el límite de distribución boreal del bosque mesófilo en México (Conabio, 2010), se entiende la baja cantidad de especies en relación con bosques similares, pero ubicados en latitudes más al sur.
Conclusiones
Se registraron 33 especies epífitas, pertenecientes a 23 géneros y 10 familias de plantas vasculares, con un total de 115 069 individuos. Se detectó asociación entre algunos taxa de epífitas y sus forofitos; asimismo son más abundantes en la parte basal e intermedia de ellas. La filogenia no explica la distribución de las epífitas.
El bosque mesófilo de montaña de la Reserva de la Biósfera El Cielo evidencia un patrón de distribución de epífitas similar al de ambientes afines en otras localidades.

