










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.9 no.49 México sep./oct. 2018
https://doi.org/10.29298/rmcf.v9i49.177
Articles
Interception loss in four scrubland species in Northeastern Mexico
1Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León, México.
Rainfall interception from the arboreal mass is the component of the hydrological cycle that is determined by the climatic conditions and the characteristics of vegetation. The aim of this study was to quantify the redistribution of precipitation by means of gross precipitation, throughfall and stemflow, estimating interception losses and canopy storage capacity for four species of thorn scrub in northeastern Mexico. For the measurement of the components of gross precipitation and throughfall, four troughs were used for each species, while for the quantification of stemflow hoses adhered in stem were used. The analysis of 47 rainfall events distributed between September 23th, 2016 to October 1st, 2017, added a total of 488.9 mm of gross precipitation. The results for Acacia farnesiana, Condalia hookeri, Leucaena leucocephala and Casimiroa greggii for throughfall showed values of 77, 76, 86 and 83 %, respectively; while the component of the stemflow oscillated from 1.12 % for C. hookeri to 1.73 % for A. farnesiana. Linear regression analyzes between gross precipitation and interception loss for four species, showed values of 13, 16, 20 and 22 % for L. leucocephala, C. greggii, A. farnesiana and C. hookeri, respectively, key species when taking management decisions on water yield in a basin; thus, the null hypothesis is rejected and it is concluded that interception loss differs among the evaluated species.
Key words: Canopy storage capacity; stemflow; Tamaulipan thorn scrub; throughfall; gross precipitation; interception loss
La intercepción de lluvia por la masa arbórea es un componente del ciclo hidrológico que está determinado por las condiciones climáticas y por las características de la vegetación. El objetivo del presente estudio fue cuantificar la redistribución de las precipitaciones vía: precipitación incidente, precipitación directa y escurrimiento fustal, a fin de estimar las pérdidas por intercepción y la capacidad del almacenamiento del dosel para cuatro especies presentes en el matorral del noreste de México. Para la medición de los componentes de precipitación incidente y directa se utilizaron cuatro canaletas por especie, y para la cuantificación del escurrimiento fustal, mangueras adheridas al fuste. El análisis de 47 eventos de lluvia, distribuidos entre el 23 de septiembre de 2016 al 1 de octubre del 2017, sumaron un total de 488.9 mm de precipitación incidente. Los resultados para Acacia farnesiana, Condalia hookeri, Leucaena leucocephala y Casimiroa greggii para la precipitación directa fueron valores de 77, 76, 86 y 83 %, respectivamente; mientras que el componente del escurrimiento fustal osciló de 1.12 % para C. hookeri a 1.73 % en A. farnesiana. Los análisis de regresión lineal entre la precipitación incidente y las pérdidas de intercepción arrojaron valores de 13, 16, 20 y 22 % para L. leucocephala, C. greggii, A. farnesiana y C. hookeri, respectivamente, especies clave al momento de tomar decisiones de manejo en el rendimiento de agua en una cuenca; con ello se rechaza con la hipótesis nula y se concluye que las pérdidas por intercepción difieren entre las especies evaluadas.
Palabras clave: Capacidad de almacenamiento del dosel; escurrimiento fustal; matorral espinoso tamaulipeco; precipitación directa; precipitación incidente; pérdidas por intercepción
Introduction
In forest ecosystems, hydrology is made up by complex processes in which biotic and abiotic factors interact. Even though the interception of rain is not the main one for determining water estimates (Webb et al., 2005), it is one of the ways that makes it possible to identify how changes in tree cover affect the hydrological balance of a forested area, which alters the content of soil moisture and surface runoff of the land (Díaz, 2007).
In semi-arid regions, the water cycle is important in the functioning of forest ecosystems, due to the prevailing water and limiting deficit; therefore, they are often considered marginal and, in general, are little studied, although they represent 50 % of the national territory (Wattenbach et al., 2005).
In a hydrological system, the amount of water intercepted can be a loss or gain of the resource, depending on the scenario. When the water retained in the vegetation cover evaporates, due to the effect of wind and temperature, it is a loss (Navarro et al., 2009). If the water comes from the humidity of the air or condensation, the interception represents an increase or gain of the resource.
Studies related to this topic have been widely investigated in cold, temperate and tropical climates, but in semi-arid and arid environments they have been scarce due to the difficulty of applying methodologies to quantify this phenomenon in shrub and herbaceous species (Dohnal et al., 2014). The wind, solar radiation, temperature and atmospheric pressure are the climatic factors that affect the hydrological cycle (López, 2007).
The redistribution of rain is the product of interception by the canopy of trees, a process that dramatically reduces the amount of precipitated water that reaches the surface of the soil; its main components are incident precipitation, direct precipitation, fusional runoff and interception losses (Yáñez et al., 2014).
Interception is defined as the amount of total precipitation that is captured during a rain event, so in addition to the vegetation other factors such as wind speed, intensity and duration of the rain, influence during each event (Flores et al., 2013). Interception in the hydrological cycle is one of the most difficult components to quantify, due to long periods of intercepted precipitation that can, in turn, evaporate into the atmosphere (Torres, 2013).
In this phenomenon, water modifies its trail through the canopy of the trees, which exerts a great influence on the rain pattern that reaches the forest floor (Návar et al., 2008). It is retained by leaves and branches, even in the presence of epiphytic plants; from this moment, it continues its passage through one or more of these three routes: evaporation, dripping from the crown, or flow through the branches to the base of the stem (García, 2006).
Incident precipitation (PI) is the total rainfall that falls on the vegetation cover. Direct precipitation (PD) is the fraction of rain that passes directly through the foliage, leaves and branches (García, 2006). Fustal runoff (Ef) consists of the fraction of precipitation that makes contact with the cover and flows down the stem, and reaches the soil surrounding the base (Iroumé and Huber, 2000). The amounts of fustal runoff are directly related to the architecture of the tree (Murakami, 2009).
Intercept losses are governed by the number of wetting and drying cycles in the plants. The amount of water stored in the canopy during a single event is sufficient to exceed the capacity of vegetation and retain water on its surface (Cantú and Okumura, 1996). When modeling in hydrology, interception is taken as a fixed percentage of precipitation; however, it strongly influences soil moisture conditions, which is transcendental in runoff (Savenije, 2004).
Among the elements involved in interception, the duration and intensity of the rain, the speed and direction of the wind, as well as the structure of the vegetation are the most studied factors (Huber, 2003, Ahmadi et al., 2009). The duration and intensity of rain is important in dry weather situations, because it increases or decreases the amount of precipitation intercepted in the foliage canopies (Toba and Ohta, 2005). High speed and wind direction are conditions that can orient the path of the leaves and reduce the chances of interception. Likewise, for tropical areas with abundant vegetation, the possibility of a drop reaching the ground without first touching any surface of vegetation is relatively low (Herwitz and Slye, 1995). The structure of vegetation refers to the morphological variations such as trunks, branches and leaves characteristic of the species, which cause differences in the ability of interception (Mastachi, 2007).
Based on the above considerations, the present study was proposed whose objectives were to determine the redistribution of precipitation, estimate the storage capacity and compare interception losses for the species Acacia farnesiana (L.) Willd., Condalia hookeri M.C.Johnst., Casimiroa greggii (S.Watson) F.Chiang and Leucaena leucocephala (Lam.) De Wit via incident precipitation, direct precipitation, and fusional runoff, according to the hypothesis that under different canopies there are no differences in the losses by interception.
Materials and Methods
Study area
The study area is located in the experimental campus of the Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León (School of Forest Sciences of the Autonomous University of Nuevo Leon) 8 km south of the Linares municipality, in the state of Nuevo León, Mexico (Figure 1), at coordinates 24°47' N; 99°32' W, 350 meters above sea level. It is located in the hydrological region RH25, known as San Fernando-Soto La Marina or Coastal Plain of the North Gulf, in the basins of the Bravo and Pánuco rivers, and the Arroyo-Camacho sub-basin, next to the Sierra Madre Oriental (Uvalle, 2008).

A subtropical and semi-arid climate prevails, with warm summer, rains between April and November, and a period of intra-festival dryness, with temperatures of up to 45 °C during the summer, with monthly averages of 14.7 °C in January to 22.3 °C in August; the annual average precipitation is 805 mm, with bimodal distribution (Saucedo, 2011).
The soil is of the Vertisol type of deep alluvial-colluvial origin of dark gray color, with a mazic-pelic subunit; fine clay-silt type textural class with high montmorillonite content and low content of organic matter and moderately alkaline pH (Uvalle, 2008).
The predominant vegetation is semiarid-subtropical, consisting of deciduous and evergreen species, with a wide range of growth patterns, diversity in leaf longevity, growth dynamics and contrasting phenological developments (González and Cantú, 2001). The Tamaulipan thorn scrub (MET, for its acronym in Spanish), is the most abundant and historically used ecosystem in the arid and semi-arid zones of northeastern Mexico (García and Jurado, 2008); it gathers a diversity of 15 000 to 21 000 ha-1 individuals of arboreal and shrub species; it includes a third of the diversity of woody flora, which is taken by the rural population for their traditional agroforestry use, as well as food source, fuel, herbalism, construction of rustic furniture and rural houses, as well as handicrafts (Foroughbakhch et al., 2009).
Species of interest
The study species are A. farnesiana, Condalia hookeri, Casimiroa greggii and L. leucocephala (Figure 2), which are part of the MET; Table 1 shows the mensuration characteristics of each species.

Figure 2 Studied species: a) Condalia hookeri M.C.Johnst., b) Acacia farnesiana (L.) Willd., c) Casimiroa greggii (S.Watson) F.Chiang and d) Leucaena leucocephala (Lam.) de Wit.
Table 1 Mensuration characteristics of the studied species (N = 4 trees sp-1.
| Species | DBH (cm) | Height (m) | Crown area (m2) |
|---|---|---|---|
| Acacia farnesiana (L.) Willd. | 28.32 | 5.30 | 42.2 |
| Condalia hookeri M.C.Johnst. | 9.54 | 8.50 | 13.5 |
| Casimiroa greggii (S.Watson) F.Chiang | 17.02 | 7.15 | 22.6 |
| Leucaena leucocephala (Lam.) de Wit. | 7.25 | 9.25 | 4.0 |
Acacia farnesiana or Huizache, belongs to the Fabaceae family. It is a small tree 2 to 4 m high, with straight stem, ascending and, occasionally, horizontal branches; alternate leaves; golden yellow flowers; fruit, resembles a reddish brown to purple or black legume with two seeds per row. In the warm zones of the country, it is part of the secondary vegetation of low deciduous uncultivated or thorny forests and thorny thickets such as the MET (García, 1997).
Condalia hookeri, or Brazil, belongs to the Rhamnaceae family. In general terms, it is a tall tree up to 9 m, with irregular expansion of the crown, alternate or fasciculate leaves, small greenish flowers, its fruit a drupe of black color when ripe. It is distributed mainly in dry soils (García, 1997).
Casimiroa greggii or Yellow Chapote, belongs to the Rutaceae family. It is a tree 4 to 8 m high with rounded crown, alternate leaves, with pointed glands; yellowish-green flower; fruit, drupe, globose, yellow when ripe with a chestnut-brown seed. Its distribution occurs in mixed forests or submontane scrub (Alanís and González, 2003).
Leucaena leucocephala or Guaje, belongs to the Fabaceae family. It is a generally tall tree from 3 to 12 m, with rounded cup slightly open and sparse; alternate leaves, bipinnate; fruit as oblong pods, stipulated, in floral chapters of 30 or more pods, 11 to 25 cm long by 1.2 to 2.3 cm wide, green when tender and brown when ripe, containing 15 to 30 seeds. It is distributed mainly in dry soils (Zárate, 1987); although this species is not native to MET, it was considered within this study due to its great density in the area, utilization and participation in its hydrological balance.
Measurement of the components of precipitation
Interception losses and storage capacity were estimated by measurements of incident precipitation, direct precipitation and stem runoff. A plot of 10 × 10 m was selected for each of the species of interest. The measurement of the components was carried out after each precipitation event, considered as any event of rain, drizzle or squall of indeterminate duration and separated by a dry period with a minimum duration of 8 hours or until the end of the total runoff in the vegetation.
The incident precipitation (PI) was determined by the use of PVC gutters of 0.1 m2 (10 cm wide × 100 cm long) and a resolution of 0.01 mm of measurement; each of them was covered with a mesh with the objective of preventing the passage of insects or any material that could obstruct the passage of water. To collect the rain, containers of 20 L capacity were installed, equipped with hoses to facilitate the collection of water during each event. Four gutters were installed 1 m above the ground in an open area without trees, next to the experimental plots.
Direct precipitation (PD) was evaluated by a similar procedure to measure the incident precipitation with four channels distributed randomly under the canopy for the species of A. farnesiana, Condalia hookeri, Casimiroa greggii and L. leucocephala.
Stem runoff (Ef) was measured in three of the four species under study except L. leucocephala due to its diameter of shafts smaller than 7 cm, preventing the quantification of this component, this being a criterion for the selection of trees. To determine Ef, a plastic hose attached to the spiral shaft was used, using a silicone adhesive sealant to prevent the loss of water.
The sum of the components PD and Ef represents the amount of net precipitation (Pn), which is the total amount of water that reaches the ground. Canopy storage capacity (CAD) is the amount of rainfall that the canopy can hold while saturated, and is determined by Leyton’s et al. method (1967), which relates the incident precipitation and direct precipitation based on rainfall events with minimum values of 2.5 mm.
With the measurements of the components mentioned above and through the application of descriptive statistics and linear regression analysis, interception losses were estimated using the following formula:
Where:
I = Loss from interception (mm)
PI = Incident precipitation (mm)
Pn = Sum of direct precipitation and stem runoff (mm)
Results and Discussion
Incident precipitation
The analysis of 47 rain events, distributed between September 22, 2016 and October 1, 2017, added a total of 488.9 mm of incident precipitation, with events in a range of 0.14 to 56.5 mm. Of the total rainfall events, 45 % were less than 5 mm of precipitation, 19 % between 5 and 10 mm and 30 % between 10 and 25 mm. Finally, events greater than 25 mm were recorded only three times and corresponded to 6 % (Figure 3).

In the monthly distribution of rainfall (Figure 4) it can be observed that the highest rainfall occurred in June 2017, with an accumulated amount of 66.0 mm, while the opposite occurred in February 2016, with only 1.8 mm in two events.

Precipitación = Rainfall; Precipitación incidente = Gross precipitation; Núm. De evento = Event number; septiembre = September; octubre =October; noviembre = November; diciembre = December; enero = January; febrero = February; marzo = March; abril = April; mayo = May; junio = June; julio = July; Agosto = August.
Figure 4 Rainfall events and monthly cumulative rainfall.
Net precipitation
The component of direct precipitation was 77 % for Acacia farnesiana, 76 % for Condalia hookeri, 86 % for Leucaena leucocephala and 83 % Casimiroa greggii, which is equivalent to 80.5 % on average of the incident precipitation. The linear regression analysis for the fusional runoff component yielded values of 1.73 % for A. farnesiana, of 1.12 % for Condalia hookeri, and of 1.29 % for Casimiroa greggii, which are relatively low and difficult to quantify in shrub species in this type of ecosystems. Cantú and Gonzáles (2005) indicate that this component represents a minimum proportion of the incident precipitation for species of the Tamaulipan thorn scrub. The average net precipitation was 81.5 %; when analyzing this variable by species, it was for Acacia farnesiana 79 %; for Condalia hookeri 77 %, for Leucaena leucocephala 86 % and for Casimiroa greggii 84 %.
Intercept losses
The intervals of interception and losses by total interception, during 45 events and for each of the four species of study are listed in Table 2. A. farnesiana recorded from 0.04 to 26.9 mm, Condalia hookeri from 0.5 to 15.0 mm, Casimiroa greggii from 0.09 to 10.6 mm, and L. leucocephala from 0.2 to 7.3 mm. Intercept losses were 20, 22, 15 and 13 % for A. farnesiana, Condalia hookeri, Casimiroa greggii and L. leucocephala, respectively (figures 5, 6, 7 and 8).
Table 2 Ranges of interception and total interception loss during the study period for the four-studied species.
| Type of dosel | n | Interception range | Total interception | |||
|---|---|---|---|---|---|---|
| (mm) | % | (mm) | % | |||
| Acacia farnesiana (L.) Willd. | 45 | 0.04-26.9 | 2.88-82.6 | 112.6 | 20 | |
| Condalia hookeri M.C.Johnst. | 45 | 0.50-14.7 | 5.88-91.4 | 145.5 | 22 | |
| Casimiroa greggii (S.Watson) F.Chiang | 40 | 0.09-10.6 | 3.36-92.2 | 111.4 | 16 | |
| Leucaena leucocephala (Lam.) de Wit. | 43 | 0.02-7.3 | 6.42-74.2 | 98.7 | 13 | |

Precipitación incidente = Gross rainfall; Intercepción = Interception.
Figure 5 Relationship between gross rainfall and interception loss for Acacia farnesiana (L.) Willd.
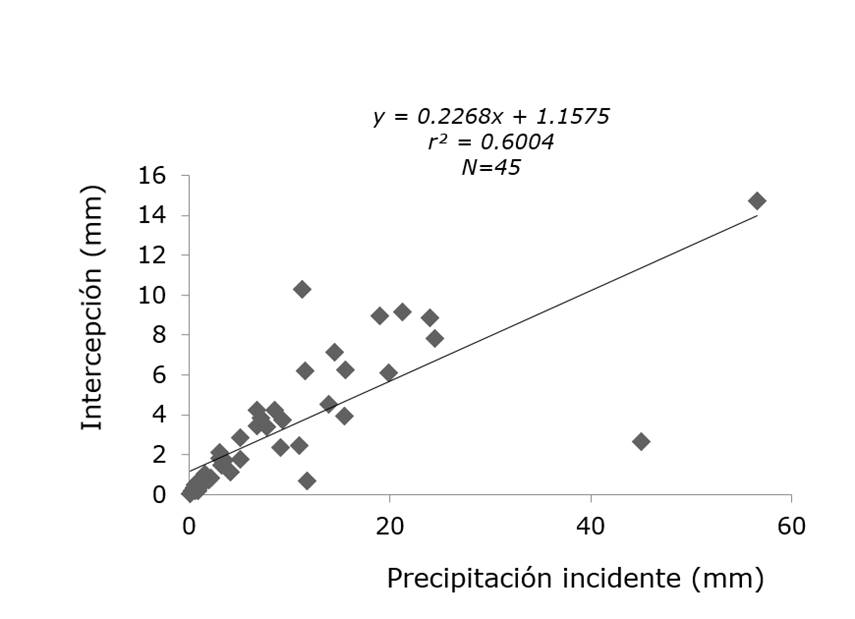
Precipitación incidente = Gross rainfall; Intercepción = Interception.
Figure 6 Relationship between gross rainfall and interception loss for Condalia hookeri M.C.Johnst.

Precipitación incidente = Gross rainfall; Intercepción = Interception.
Figure 7 Relationship between gross rainfall and interception loss for Leucaena leucocephala (Lam.) de Wit.

Precipitación incidente = Gross rainfall; Intercepción = Interception.
Figure 8 Relationship between gross rainfall and interception loss for Casimiroa greggii (S.Watson) F.Chiang.
Likewise, Cantú and González (2005) calculated interception losses of 18, 15 and 22 % for Acacia berlandieri Benth., Acacia rigidula Benth. and Diospyros texana Scheele, respectively. Yáñez et al. (2014) reported intercept values of 33, 34, 15 and 25 %, respectively for Helietta parvifolia (Gray ex Hemsl.) Benth., Ebenopsis ebano (Berland.) Barneby & J. W. Grimes, Eucalyptus camaldulensis Dehnh. and Prosopis laevigata (Humb. & Bonpl.) ex Willd. in the same region.
Belmonte and Romero (1999) mention that in arid and semi-arid environments, interception losses can be in some cases greater than 35 %. Gerrits and Savenije (2011) report that in temperate forests it varies between 9 and 48 % of gross precipitation.
López (2007) calculated that in herbaceous vegetation or shrubland, 3 to 10.9 % of total rainfall is lost. Sadeghi et al. (2015) managed a wide range of interception in semi-arid environments, comprising values from 2.5 to 56 % of the total recorded rainfall. In another study conducted in a community of thorny, unarmed scrubland where Acacia farnesiana (huizache) predominates in Atotonilco El Grande, Hidalgo, Flores et al. (2013) obtained an average interception of 18.6% and an r2 = 0.4076. This result is very similar to the present study for the same species with an intercept of 20 % and r2 = 0.4122.
The results of the regression analysis between the incident precipitation and the intercept for A. farnesiana, Condalia hookeri, Casimiroa greggii and L. leucocephala showed that the values of the coefficient of determination (r2) were 0.41, 0.60, 0.51 and 0.49, respectively (Table 3); this indicates that other factors influence the interception losses such as the duration and intensity of rain, wind speed and direction and the structure of vegetation. The values of Cantú and González (2005) for A. berlandieri were of 0.735, for A. rigidula of 0.614 and for D. texana of 0.718; Yáñez (2014) calculated 0.723 for H. parvifolia; 0.821 for Ebenopsis ebano; 0.395 for Eucalyptus camaldulensis and 0.571for P. laevigata.
Table 3 Results of the regression analyzes for interception losses for the canopies of the studied species.
| Type of dosel | n | Y- value of the intercept (β ̥) | Slope (β 1) | r2 Ajusted |
||||
|---|---|---|---|---|---|---|---|---|
| (β ̥) | (EEE) | (P-value) | (β 1) | (EEE) | (P-value) | |||
| Acacia farnesiana (L.) Willd. | 45 | 0.3174 | 1.791 | 0.003 | 0.208 | 0.360 | < 0.001 | 0.41 |
| Condalia hookeri M.C.Johnst. | 45 | 1.1575 | 1.527 | 0.699 | 0.226 | 0.329 | < 0.001 | 0.60 |
| Casimiroa greggii (S.Watson) F.Chiang | 40 | 1.0821 | 2.076 | 0.411 | 0.161 | 0.502 | < 0.001 | 0.51 |
| Leucaena leucocephala (Lam.) de Wit. | 43 | 1.0701 | 1.784 | 0.748 | 0.137 | 0.566 | < 0.001 | 0.49 |
Storage capacity of the canopy
The storage capacity of the canopy for Condalia hookeri registered the highest saturation value with 0.09 mm and with a coefficient of determination of r2 = 0.83 and the opposite occurred with L. leucocephala with 0.0006 mm and a r2 = 0.60 (Table 4). In this context, Cantú and González (2005) calculated 0.24 mm for A. berlandieri, 0.14 mm for A. rigidula and 0.14 mm for D. texana. Yáñez et al. (2014) did it for H. parvifolia with 0.0303 mm, for Ebenopsis ebano with 0.1201, for Eucalyptus camaldulensis with 0.2782 mm and for P. laevigata 0.1558 mm. In several species of scrub, Návar and Bryan (1994) obtained a greater storage capacity with 0.87 mm in a range of 0.39 to 1.59 mm, which they attributed to storms with contributions less than 1 mm of rain.
Conclusions
From the recorded rainfall events, 45 % came from events less than 5 mm. Leucaena leucocephala and Casimiroa greggii presented the highest amount of direct precipitation, while Acacia farnesiana and Condalia hookeri intercepted a greater amount of rainfall with respect to the incident precipitation L. leucocephala recorded the lowest loss by interception
The component of the stem runoff has a minimum contribution on the net precipitation and a difference between the storage capacity of the canopy. Based on the results obtained, it is concluded that interception losses differ among the taxa evaluated, which makes this information an important tool for decision making regarding the selection of species and their impact on water yield in a basin of the characteristics of the one studied.
Acknowledgements
The authors wish to thank the Facultad de Ciencias Forestales de la Universidad Autónoma de Nuevo León and CONACyT Fondo Ciencia Básica SEP-CONACYT, Project No. 250732 for their support in the achievement of the actual study. Also, to the two anonymous reviewers for improving this paper with their critical observations.
REFERENCES
Ahmadi, M. T., P. Attarod., M. R. Marvi M., R. Rahmani and J. Fathi. 2009. Partitioning rainfall into throughfall, stemflow, and interception loss in an oriental beech (Fagus orientalis Lipsky) forest during the growing season. Turkish Journal of Agriculture and Forestry 33(6): 557-568. [ Links ]
Alanís, G. J. y D. González. 2003. Flora nativa ornamental para el área metropolitana de Monterrey, Nuevo León, México. Descripción y requerimiento de las especies para el paisaje urbano. Monterrey, México: Universidad Autónoma de Nuevo León, R. Ayuntamiento de Monterrey. Monterrey, NL., México. 128 p. [ Links ]
Belmonte S., F. y A. Romero D. 1999. Interceptación en algunas especies del matorral mediterráneo. Primera edición. Universidad de Murcia. Murcia, España. 202 p. [ Links ]
Cantú S., I. and T. Okumura. 1996. Throughfall, stemflow and interception loss in a mixed white Oak forest (Quercus serrata Thunb.) Journal of Forest Research 1(3): 123-129. [ Links ]
Cantú S., I. y H. González R. 2005. Pérdidas por intercepción de la lluvia en tres especies de matorral submontano. Revista Ciencia UANL. 3(1):80-84. [ Links ]
Díaz, A. 2007. Series temporales de vegetación para un modelo hidrológico distribuido. Grupo de hidrología e hidráulica agrícola. Maestría en Hidráulica Ambiental. Departamento de Agronomía, Universidad de Córdoba. Córdoba, España. 121 p. [ Links ]
Dohnal, M., T. Černý, J. Votrubová and M. Tesař. 2014. Rainfall interception and spatial variability of throughfall in spruce stand. Journal of Hydrology and Hydromechanics 62(4):277-284. [ Links ]
Foroughbakhch, R., J. L. Hernández-Piñero, M. A. Alvarado-Vázquez, E. Céspedes-Cabriales, A. Rocha-Estrada and M. L. Cárdenas-Ávila. 2009. Leaf biomass determination on woody shrub species in semiarid zones. Agroforestry Systems 77(3): 181-192. [ Links ]
Flores A., E., F. Becerra L., E. Buendía R., F. Carrillo A., G. Terrazas H., T. Pineda y M. Acosta M. 2013. Intercepción de lluvia por matorral inerme espinoso en Atotonilco el Grande, Hidalgo. Revista Mexicana de Ciencias Agrícolas 4(2): 285-298. [ Links ]
García A., L. C. 1997. Estudio fenológico y de crecimiento de once especies leñosas del matorral espinoso tamaulipeco en Linares, Nuevo León, México. Tesis Maestría. Facultad de Ciencias Forestales. Linares, NL., México. 92 p. [ Links ]
García E., M. 2006. Efecto de la estructura de la copa en la partición de lluvia de tres especies arbustivas en clima semiárido. Tesis Doctoral. Universidad de Almería. Almería, España. 438 p. [ Links ]
García, J. y E. Jurado. 2008. Caracterización del matorral con condiciones prístinas en Linares, NL. México. Revista Ra Ximhai 4(1): 1-21. [ Links ]
Gerrits A., M. J. and G. H.Savenije. 2011. Interception. In: Wilderer, P. (ed.). Treatise on Water Science. Elsevier Science. Oxford, England. pp. 89-101. [ Links ]
González R., H. e I. Cantú S. 2001. Adaptación a la sequía de plantas arbustivas de matorral espinoso tamaulipeco. Ciencia UANL 4(4): 454-459. [ Links ]
Herwitz, S. R. and R. E. Slye 1995. Three-dimensional modeling of canopy tree interception of wind-driven rainfall. Journal of Hydrology 168(1-4): 205-226. [ Links ]
Huber, A. 2003. Cambios en el balance hídrico provocado por la forestación con Pinus radiata D. Don en el secado interior del centro de Chile. Gestión Ambiental 9: 57-66. [ Links ]
Iroumé, A. y A. Huber. 2000. Intercepción de las lluvias por la cubierta de bosques y efecto en los caudales de crecida en una cuenca experimental en Malalcahuello, IX Región, Chile. Bosque (Valdivia) 21(1):45-56. [ Links ]
Leyton, L. and R. Reynolds E., B. Thompson F. 1967. Rainfall interception in a forest and moorland. In: Sopper, W. E. and H. W. Lull (eds.). International Symposium on Forest Hydrology. Pergamon Press. Toronto, Canada. pp. 163-178. [ Links ]
López L., A. A. 2007. Intercepción de la vegetación herbácea de ambiente semiárido y su efecto en la relación intensidad-duración-frecuencia de la precipitación. Tesis de Maestría. Universidad Autónoma de Querétaro. Querétaro, Qro., México. 173 p. [ Links ]
Mastachi L., C. 2007. Intercepción de precipitación en dos especies de ambiente semidesértico: Acacia farnesiana y Prosopis laevigata. Tesis de Maestría. Universidad Autónoma de Querétaro. Querétaro, Qro., México.150 p. [ Links ]
Murakami, S. 2009. Abrupt changes in anual stemflow with growth in a young stand of Japanese cypress. Hydrological Research Letters 3:32-35. [ Links ]
Návar, J. and R. B. Bryan 1994. Fitting the analytical model of rainfall interception of Gash to individual shrubs of semi-arid vegetation in northeastern México. Agricultural and Forest Meteorology 68:133-143. [ Links ]
Návar C., J. J., J. Méndez G. y H. González R. 2008. Intercepción de la lluvia en especies de leguminosas del nordeste de México. Terra Latinoamericana 26(1):61- 68. [ Links ]
Navarro, J., A. Martínez. A. y J. Mongil. 2009. Hidrología de conservación de aguas: captación de precipitaciones horizontales y escorrentías en zonas secas. Universidad de Valladolid- Secretariado de Publicaciones e Intercambio Editorial. Valladolid, España. 446 p. [ Links ]
Sadeghi, S. M. M., P. Attarod and T. G. Pypker. 2015. Differences in rainfall interception during the growing and non-growing seasons in a Fraxinus rotundifolia Mill. plantation located in a semiarid climate. Journal of Agricultural Science and Technology 17(1):145-156. [ Links ]
Saucedo, Q. E. 2011. Estudio etnobotánico de especies arbustivas y arbóreas en los municipios de Linares y Hualahuises, Nuevo León, México. Tesis de Maestría. Facultad de Ciencias Forestales, Universidad Autónoma de Nuevo León. Linares, N.L., México. León. 83p. [ Links ]
Savenije, H. H. G. 2004. The importance of interception and why we should delete the term evapotranspiration from our vocabulary. Hydrological Processes 8(8): 1507-1511. [ Links ]
Toba, T. and T. Ohta. 2005. An observational study of the factors that influence interception loss in boreal and temperate forests. Journal of Hydrology 313(3-4): 208-220. [ Links ]
Torres, B. 2013. Características y funciones hidrológicas de los bosques nublados en la provincia de Zamora Chinchipe. Tesis de Maestría. Universidad de Zamora. Zamora- Chinchipe, Ecuador. 91 p. [ Links ]
Uvalle J., I. 2008. Características fisiológicas y nutrimentales en especies arbustivas forrajeras nativas de la flora del noreste de México. Tesis de Doctorado. Facultad de Ciencias Biológicas. Universidad Autónoma de Nuevo León. San Nicolás de los Garza, NL., México. 171 p. [ Links ]
Wattenbach, D., M. Hattermann., K. Weng, F. Wechsung, V. Krysanova and F. Badeck. 2005. A simplified approach to implement forest ecohydrological properties in regional hydrological modeling. Ecological Modeling 187(1):40-59. [ Links ]
Webb, T. J., F. I. Woodward, L. Hannah and K. J. Gaston. 2005. Forest cover rainfall relationship in a biodiversity hotspot: The Atlantic Forest of Brasil. Ecological Applications 15(6):1968-1983. [ Links ]
Yáñez D., M. I., I. Cantú S., H. González R. y J. I. Uvalle S. 2014. Redistribución de la precipitación en tres especies arbustivas nativas y una plantación de eucalipto del noreste de México. Tecnología y Ciencias del Agua 2: 71-84. [ Links ]
Zárate P., S. 1987. Taxonomic identity of Leucaena leucocephala (Lam.) de Wit, with a new combination. Phytologia 63(4):304-306. [ Links ]
Received: December 14, 2017; Accepted: July 27, 2018
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
