











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
La capacidad productiva de una masa forestal está definida por la especie o grupo de especies y por los factores bióticos y abióticos que interactúan con el ambiente; por tanto, el crecimiento y la producción de madera en rodales forestales están determinados por la distribución de edades, la capacidad de producción innata que soporta un área determinada, y su grado de utilización, así como por los tratamientos silviculturales aplicados (Clutter et al., 1983; Torres y Magaña, 2001; Diéguez-Aranda et al., 2009).
Las herramientas cuantitativas modernas pueden usarse para optimizar la planeación, seguimiento y control del manejo forestal sustentable, ya que contribuyen a determinar el turno, intervenciones silvícolas y la cosecha de madera, de forma más precisa. Por lo tanto, los modelos matemáticos permiten inferir el comportamiento biológico de árboles individuales o rodales para la planeación de escenarios de manejo forestal, basados en modelos de predicción y proyección (Quiñonez-Barraza et al., 2015).
Con la relación altura dominante-edad, es factible obtener, de manera implícita, el índice de sitio (IS), el cual se define como el valor de la altura dominante de una especie determinada a una edad base de referencia (Clutter et al., 1983); esta última, por lo general, se relaciona con el turno biológico de rodales con manejo forestal. Una familia de curvas de índice de sitio (denominadas también curvas de calidad de estación o de productividad) es un conjunto de curvas que muestran el patrón de crecimiento en altura dominante que los rodales de una especie en particular y zona geográfica seguirán a lo largo del tiempo (Rennolls, 1978; Bengoa, 1999), cada una de las cuales lleva asociado un número arábigo que las identifica (valor del índice de sitio) o simplemente el valor del IS. A su vez, las líneas intermedias entre cada curva de calidad limitan franjas que se denominan clases de calidad.
Las curvas son anamórficas o polimórficas, cuyas ecuaciones se generan con el método de diferencias algebraicas (Algebraic Difference Approach o ADA por las siglas en inglés) (Bailey y Clutter, 1974), o bien las familias de curvas con polimorfismo asintótico que combina las tasas de crecimiento con la potencialidad del sitio, y que se obtienen con el método de diferencia algebraica generalizada (Generalized Algebraic Difference Approach o GADA por las siglas en inglés) (Cieszewski y Bailey, 2000; Cieszewski, 2002; Cieszewski, 2003). Este método de ajuste de ecuaciones dinámicas permite que en una ecuación base se considere la variación en las tasas instantáneas de crecimiento en cada curva y la potencialidad del sitio, lo que resulta ideal para prescribir tratamientos silvícolas en rodales, de acuerdo a su potencial productivo (Santiago-García et al., 2013).
En los bosques de la Unidad de Manejo Forestal Regional 2001 (Umafor), en Oaxaca, México son escasas las ecuaciones de índice de sitio para las especies del género Pinus que puedan utilizarse como herramientas biométricas para la planeación y ejecución del manejo forestal sustentable. El objetivo de este trabajo fue construir modelos dinámicos de índice de sitio para Pinus oaxacana Mirov, P. douglasiana Martínez, P. patula Schiede ex Schltdl. & Cham. y P. pseudostrobus Lindl. en la Sierra Norte de Oaxaca, mediante el uso de la metodología GADA.
Materiales y Métodos
Área de estudio
El estudio se llevó a cabo en los rodales bajo aprovechamiento forestal localizados en la Umafor 2001, ubicada al noreste del estado de Oaxaca, México, y que abarca una superficie de 195 397 ha. Se localiza a los 17°22’58” N y los 96°22’29” O (Figura 1). La altitud de las áreas de muestreo varía de 1 800 a 3 361 m. La temperatura anual oscila entre -3 °C y 28 °C, con precipitación promedio anual de 3 797 mm (Granados-Sánchez, 2009). El tipo de vegetación predominante corresponde a bosques de pino-encino, con dominancia de Pinus oaxacana, P. douglasiana, P. patula, P. pseudostrobus, P. leiophylla Schiede ex Schltdl. & Cham., P. pringlei Shaw., P. rudis Endl., P. teocote Schiede ex Schltdl. & Cham., Quercus crassifolia Bonpl., Q. laurina Bonpl. y Q. rugosa Née, principalmente.

Datos
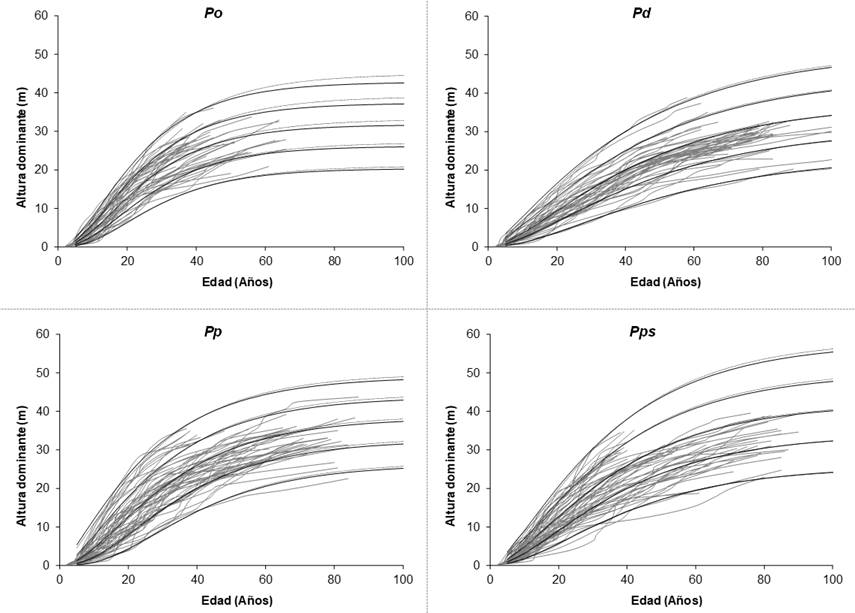
Para ajustar las ecuaciones dinámicas de altura dominante e IS se usó una base de datos procedente de 206 análisis troncales, integrada por 44 árboles de Pinus oaxacana, 45 de P. douglasiana, 58 de P. patula, y 59 de P. pseudostrobus. Los individuos fueron derribados a una altura del tocón que varió de 0.1 m a 0.25 m, de tal manera que la primera sección correspondió a la altura del tocón, la segunda a 1.3 m y posteriormente, cada 2.54 m hasta llegar a la punta del árbol. Los ejemplares se seleccionaron en rodales mezclados y se intentó cubrir las diferentes calidades de estación, así como las categorías de diámetro presentes en las áreas con manejo forestal. Si bien, el concepto de índice de sitio está diseñado para masas coetáneas y puras (Daniel et al., 1979), el carácter de dominante para un árbol implica que estos individuos son poco sensibles al efecto de la densidad, la competencia y la mezcla de especies, ya que su atura se mantiene constante en un intervalo amplio de densidades y grados de mezcla, lo cual refleja a lo largo de su vida el potencial productivo de un sitio en particular, y por ello el concepto se ha hecho extensivo a las masas de pino mezcladas (Corral et al., 2004; Vargas-Larreta, 2010; Castillo et al., 2013).
Los estadísticos descriptivos de la base de datos se presentan en el Cuadro 1, se observa que la muestra cubre un intervalo amplio de combinaciones edad-altura dominante, por lo que se considera adecuada para el ajuste de las ecuaciones dinámicas de crecimiento.
Cuadro 1 Estadísticos descriptivos de los datos de edad y altura dominante por especie, de la muestra utilizada para ajustar las ecuaciones dinámicas de índice de sitio.
| Clave | Especie | Árboles | n | Edad (años) | Altura dominante (m) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mín. | Máx. | Media | SD. | Mín. | Máx. | Media | SD | ||||
| Po | Pinus oaxacana Mirov | 44 | 563 | 2 | 66 | 21.8 | 13.7 | 0.1 | 36.1 | 13.5 | 9.2 |
| Pd | Pinus douglasiana Martínez | 45 | 638 | 2 | 126 | 36.2 | 24.1 | 0.1 | 38.8 | 15.3 | 9.9 |
| Pp | Pinus patula Schiede ex Schltdl. & Cham. | 58 | 804 | 2 | 87 | 27.3 | 19.1 | 0.1 | 43.7 | 15.4 | 10.6 |
| Pps | Pinus pseudostrobus Lindl. | 59 | 811 | 2 | 90 | 29.0 | 20.5 | 0.1 | 39.5 | 15.2 | 10.3 |
| Total | 206 | 2816 | 2 | 126 | 28.7 | 20.4 | 0.1 | 43.7 | 15.0 | 10.1 | |
n = Número de pares de altura dominante - edad; Min. = Valor mínimo; Máx. = Valor Máximo; SD = Desviación estándar.
Las alturas verdaderas de las secciones se estimaron con el método de Carmean (1972) modificado por Newberry (1991), ya que con este se han tenido buenos resultados en otras investigaciones (Dyer y Bailey, 1987; Fabbio et al., 1994; Castillo et al., 2013). El método se basa en dos supuestos: i) entre dos secciones, el árbol crece a un ritmo constante, y ii) el corte se realiza, como promedio, en el centro del crecimiento en altura de un año (Dyer y Bailey, 1987; Amaral et al., 2010). Las ecuaciones empleadas para calcular la altura verdadera varían según la sección del árbol (ecuaciones 1-3).
Donde:
H 1 y H 2 = Alturas de las secciones inferior y superior de la troza
N 1 y N 2 = Número de anillos de las secciones inferior y superior de la troza
N 0 = Edad del árbol; es decir, el número de anillos del tocón (en el tocón N 0 = N 1 )
T 0 = Edad del árbol cuando alcanzó la altura H1, es decir N 0 - N 1
T = Número entero de 1 a N 1 - N 2
Ecuaciones dinámicas de índice de sitio
Se ajustaron tres ecuaciones con el método GADA, las cuales se han documentado en diversos estudios para describir el crecimiento en altura dominante e índice de sitio, esta son: Korf (4), Hossfeld (5) y Bertalanffy-Richards (6) (Cieszewski, 2002; Castillo et al., 2013; Quiñonez-Barraza et al., 2015; González et al., 2016). La expresión de los modelos base, así como su formulación GADA se presentan en el Cuadro 2.
Cuadro 2 Ecuaciones dinámicas de índice de sitio evaluadas en este estudio.
| Ecuación base |
Parámetros del sitio |
Solución para X0 | Expresión GADA | Ecuación |
|---|---|---|---|---|
|
|
|
|
|
(4) |
|
|
|
|
|
(5) |
|
|
|
Donde: |
|
(6) |
H = Altura dominante; t = Edad en años
Descripción del método GADA
El desarrollo de cualquier ecuación dinámica con la formulación GADA considera: i) seleccionar una ecuación base e identificar el número de parámetros que serán dependientes de la productividad del sitio; ii) los parámetros seleccionados se expresan como funciones de la calidad de estación definida por la variable X 0 (variable no observable e independiente que describe la productividad del sitio, como resultado de la suma de factores ecológicos, climáticos, regímenes de manejo, condiciones del suelo y los parámetros nuevos; iii) la ecuación base bidimensional (H=f(t)) seleccionada es expandida a una ecuación tridimensional (H=f(t, X 0 )) de índice de sitio; y iv) se despeja el valor de X 0 , a partir de condiciones iniciales del sitio; es decir, de valores de partida de altura dominante - edad (t 0 , H 0 ), de forma que el modelo pueda definirse implícitamente y aplicarse en la práctica (H=f(t, t 0 , H 0 )) (Cieszewski y Bailey, 2000; Cieszewski, 2001; Cieszewski, 2002; Cieszewski, 2003). Durante el proceso, a menudo se eliminan parámetros redundantes y se obtiene un modelo con un número de parámetros menor o igual que la ecuación base original.
Método y estadísticos de ajuste para la comparación de los modelos
Los parámetros globales y específicos del sitio se estimaron con el método iterativo anidado (nested iterative procedure) (Tait et al., 1988; Cieszewski, 2003; Krumland y Eng, 2005), el cual se emplea cuando las bases de datos proceden de análisis troncales o parcelas permanentes. Los modelos se ajustaron con estructura de error autorregresiva de tiempo continuo (CARi), para corregir problemas potenciales de autocorrelación del término del error (Zimmerman et al., 2001; Nord-Larsen, 2006; Crecente-Campo et al., 2009) y obtener estimadores de los parámetros insesgados y eficientes (Parresol y Vissage, 1998).
Donde:
La determinación de la bondad de ajuste de los modelos se basó en resultados de análisis cuantitativos, como la raíz del error medio cuadrático (REMC), el coeficiente de determinación ajustado (R 2 adj ), el sesgo promedio (e) y gráficos de residuales para comparar los aspectos siguientes: i) comportamiento biológico de las predicciones de los diferentes modelos ajustados; ii) representación de los residuales frente a los valores predichos por el modelo, y iii) representación de los residuales frente a residuales con diferentes retardos para comprobar la corrección de la autocorrelación de los errores, mediante la modelación de la estructura del error (Goelz y Burk, 1992; Sharma et al., 2011). La presencia de la autocorrelación en los datos utilizados se calculó con el estadístico de Durbin-Watson (Durbin y Watson, 1951). Las expresiones de dichos estadísticos son:
Donde:
n = Número de observaciones
p = Número de parámetros del modelo
e = Valor residual del modelo ajustado
El ajuste simultáneo de las ecuaciones de crecimiento y de la estructura del error (CAR2) se realizó con el procedimiento MODEL del paquete estadístico SAS/ETSTM (SAS, 2004), que permite una actualización dinámica de los residuos en el procedimiento de ajuste.
Resultados y Discusión
Ecuaciones GADA generadas, sin considerar la estructura del error
Los parámetros estimados, los errores estándar y los estadísticos de ajuste de las ecuaciones dinámicas analizadas para Pinus oaxacana, P. douglasiana, P. patula y P. pseudostrobus, se obtuvieron para todos los pares de altura dominante - edad, con el ajuste de las ecuaciones 4, 5 y 6, sin considerar la modelación de la estructura del error (Cuadro 3). En los resultados se observan problemas de autocorrelación de los residuos, debido a las mediciones de altura en un mismo árbol (Figura 4).
Cuadro 3 Parámetros estimados y estadísticos de ajuste de las ecuaciones ajustadas sin considerar la modelación de la autocorrelación.
| Especie | Ecuación | Parámetro | Valor estimado | EE | Valor-t | Valor-p |
REMC (m) |
R 2adj | DW |
|---|---|---|---|---|---|---|---|---|---|
| Po | 4 | b 1 | -9.9723 | 6.2560 | -1.59 | 0.1115 | 1.23 | 0.98 | 0.73 |
| b 2 | 107.1878 | 27.0767 | 3.96 | <0.0001 | |||||
| b 3 | 0.7720 | 0.0268 | 28.76 | <0.0001 | |||||
| 5 | b 1 | 10.3192 | 4.3182 | 2.39 | 0.0172 | 1.22 | 0.98 | 0.70 | |
| b 2 | 19 712.5600 | 3 295.9000 | 5.98 | <0.0001 | |||||
| b 3 | 1.9817 | 0.0357 | 55.56 | <0.0001 | |||||
| 6 | b 1 | 0.0546 | 0.0016 | 33.54 | <0.0001 | 1.24 | 0.98 | 0.71 | |
| b 2 | -1.0411 | 0.5854 | -1.78 | 0.0759 | |||||
| b 3 | 11.9310 | 2.1632 | 5.52 | <0.0001 | |||||
| Pd | 4 | b 1 | -24.6624 | 4.8423 | -5.09 | <0.0001 | 1.24 | 0.98 | 0.62 |
| b 2 | 165.1131 | 21.2032 | 7.79 | <0.0001 | |||||
| b 3 | 0.6067 | 0.0246 | 24.64 | <0.0001 | |||||
| 5 | b 1 | 20.4518 | 2.3387 | 8.74 | <0.0001 | 1.19 | 0.98 | 0.64 | |
| b 2 | 11 664.3400 | 1 459.4000 | 7.99 | <0.0001 | |||||
| b 3 | 1.6595 | 0.0292 | 56.74 | <0.0001 | |||||
| 6 | b 1 | 0.0300 | 0.0010 | 29.98 | <0.0001 | 1.18 | 0.99 | 0.65 | |
| b 2 | -1.8106 | 0.2727 | -6.64 | <0.0001 | |||||
| b 3 | 12.8909 | 1.0198 | 12.64 | <0.0001 | |||||
| Pp | 4 | b 1 | -762.5240 | 289.6000 | -2.63 | 0.0086 | 1.77 | 0.97 | 0.61 |
| b 2 | 3 227.2260 | 1 191.2000 | 2.71 | 0.0069 | |||||
| b 3 | 0.8459 | 0.0268 | 31.52 | <0.0001 | |||||
| 5 | b 1 | 40.5627 | 0.8353 | 48.56 | <0.0001 | 1.74 | 0.97 | 0.59 | |
| b 2 | 718.3985 | 491.1000 | 1.46 | 0.1439 | |||||
| b 3 | 2.0036 | 0.0329 | 60.96 | <0.0001 | |||||
| 6 | b 1 | 0.0447 | 0.0014 | 30.95 | <0.0001 | 1.85 | 0.97 | 0.56 | |
| b 2 | -9.4612 | 1.0574 | -8.95 | <0.0001 | |||||
| b 3 | 42.7925 | 3.9189 | 10.92 | <0.0001 | |||||
| Pps | 4 | b 1 | -23.2364 | 5.8717 | -3.96 | <0.0001 | 1.49 | 0.98 | 0.61 |
| b 2 | 163.9336 | 25.7222 | 6.37 | <0.0001 | |||||
| b 3 | 0.6937 | 0.0228 | 30.38 | <0.0001 | |||||
| 5 | b 1 | 26.7838 | 2.0150 | 13.29 | <0.0001 | 1.47 | 0.98 | 0.62 | |
| b 2 | 10 535.2800 | 1 554.6000 | 6.78 | <0.0001 | |||||
| b 3 | 1.8045 | 0.0297 | 60.82 | <0.0001 | |||||
| 6 | b 1 | 0.0368 | 0.0011 | 32.21 | <0.0001 | 1.51 | 0.98 | 0.60 | |
| b 2 | -0.7945 | 0.3176 | -2.50 | 0.0126 | |||||
| b 3 | 10.0448 | 1.2201 | 8.23 | <0.0001 |
EE = Error estándar del estimador del parámetro; REMC = Raíz del error medio cuadrático; R 2 adj = Coeficiente de determinación ajustado; DW = Estadístico de Durbin-Watson; Po = Pinus oaxacana; Pd = Pinus douglasiana; Pp = Pinus patula; Pps = Pinus pseudostrobus.

Figura 2 Sobreposición de las curvas de IS generadas con los modelos de Korf (línea discontinua), Hosffeld (línea punteada) y Bertalanffy-Richards (línea continua).
Las curvas se generaron con el modelo dinámico de Bertalanffy-Richards sin (línea punteada) y con (línea continua) corrección de la autocorrelación mediante una estructura autorregresiva CAR2.
Figura 3 Curvas de índice de sitio a la edad de referencia de 40 años para Pinus oaxacana Mirov (Po), P. douglasiana Martínez (Pd), P. patula Schiede ex Schltdl. & Cham. (Pp) y P. pseudostrobus Lindl. (Pps).
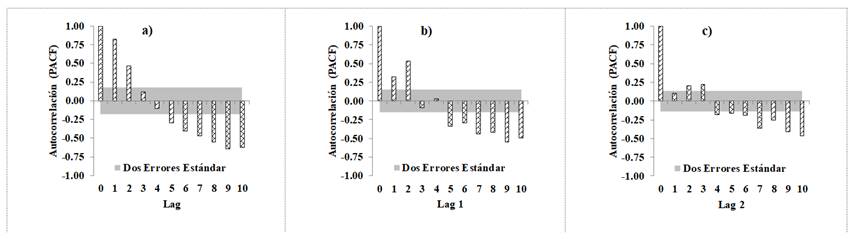
Figura 4 Gráficas de la Función de Autocorrelación Parcial de la ecuación de Bertalanffy-Richards (Ecuación 6): a) ajuste sin corrección de la autocorrelación, b) al considerar un modelo CAR1, y c) con un modelo CAR2.
Los coeficientes de determinación ajustados (R 2 adj ) por el número de parámetros en los modelos de Korf, Hossfeld y Bertalanffy-Richards para las diferentes especies muestran que es posible explicar más de 96 % de la varianza observada en el crecimiento en altura dominante, en función de la edad, con estimaciones de parámetros significativos para la mayoría de las combinaciones de ecuaciones y especies (p<0.01). Los errores medios en los tres modelos oscilan entre 1.18 m (Pd) y 1.85 m (Pp) (Cuadro 3).
En general, las tres ecuaciones ajustadas muestran estadísticos de ajuste satisfactorios para predecir el crecimiento en altura dominante en función de la edad, sin expandir los términos del error para corregir la autocorrelación. Por otro lado, el análisis gráfico muestra un comportamiento biológico realista (predicciones similares a los datos observados). Sin embargo, en Pinus oaxacana el parámetro b 1 fue no significativo en los modelos Korf (4) y Hossfeld (5) por lo que se acepta la hipótesis nula de que los estimadores de los parámetros son iguales a cero; escenario que también ocurrió para el modelo Bertalanffy-Richards (6) en el parámetro b 2 . El parámetro b 2 no fue significativo en el modelo de Hossfeld (5) para Pinus patula Schiede ex Schltdl. & Cham., situación que se presentó para el mismo parámetro en el modelo Bertalanffy-Richards (6) para Pinus pseudostrobus, todos a un nivel de significancia de 5 %.
Los resultados indican que las diferencias entre los estadísticos de bondad de ajuste fueron mínimas entre los modelos evaluados, por lo que la selección del más apropiado se apoyó en el análisis gráfico de sobreponer las familias de curvas de crecimiento de altura dominante a las trayectorias de crecimiento de los datos observados; se consideraron categorías de IS de 15 m a 35 m para Pinus oaxacana, de 10 m a 30 m para Pinus douglasiana, con intervalos de 5 m; y de 14 m a 38 m, para Pinus patula y Pinus pseudostrobus, con intervalos de 6 m a una edad de referencia de 40 años para todos los casos (Figura 2). Lo anterior, con base en que diferentes modelos pueden presentar los mismos estadísticos de bondad de ajuste o de comparación, pero una respuesta distinta a las trayectorias de los datos observados; así, unos subestiman en las primeras edades y sobreestiman en las avanzadas, o viceversa (Diéguez-Aranda et al., 2006).
Se seleccionó el modelo de Bertalanffy-Richards (6) como el más adecuado para describir las trayectorias de crecimiento de los árboles que integraron la base de datos, y para comparar las estimaciones de crecimiento frente al mismo modelo, a partir de la modelación de la estructura del error, con una estructura autocorregresiva de errores continua tipo CAR1 y CAR2, en virtud de que este modelo describió ligeramente mejor las tendencias individuales de crecimiento en altura dominante, principalmente, a edades superiores a 50 años (Figura 2).
La ecuación de Bertalanffy-Richards, en la que se considera una estructura CAR2 fue capaz de modelar las tendencias de crecimiento de los datos experimentales para las especies estudiadas, y describir patrones de variación en el crecimiento de las curvas construidas, así como potencialidades de sitio diferentes. Este resultado es consistente con otros estudios en los que también se recomienda dicho modelo para la construcción de curvas de índice de sitio de otras especies de interés comercial (Kiviste et al., 2002; Rodríguez-Carrillo et al., 2015; Castillo et al., 2013).
Modelación de la estructura del error
En el Cuadro 4 se muestran los estadísticos del ajuste del modelo de Bertalanffy-Richards, en el que se integra un modelo autorregresivo continuo de segundo orden (CAR2), el cual corrigió de manera adecuada la autocorrelación de los errores de la base de datos (valor de DW cercano a 2).
Cuadro 4 Estimaciones de los parámetros y estadísticos de bondad de ajuste de la ecuación de Bertalanffy-Richards, integrando el modelo autorregresivo continúo CAR2.
| Ecuación | Especie | Parámetro | Valor estimado | EE. | Valor-t | Valor-p | REMC (m) | R 2 adj | DW |
|---|---|---|---|---|---|---|---|---|---|
| 6 | Po | b 1 | 0.059830 | 0.00214 | 27.98 | <0.0001 | 0.87 | 0.99 | 1.82 |
| b 2 | -2.475350 | 0.94550 | -2.62 | 0.0091 | |||||
| b 3 | 17.211490 | 3.42410 | 5.03 | <0.0001 | |||||
| p 1 | 0.977894 | 0.00973 | 100.45 | <0.0001 | |||||
| p 2 | 0.906721 | 0.01090 | 83.15 | <0.0001 | |||||
| 6 | Pd | b 1 | 0.027946 | 0.00127 | 22.00 | <0.0001 | 0.79 | 0.99 | 1.81 |
| b 2 | -2.135910 | 0.40310 | -5.30 | <0.0001 | |||||
| b 3 | 13.592790 | 1.49220 | 9.11 | <0.0001 | |||||
| p 1 | 0.997808 | 0.00469 | 212.85 | <0.0001 | |||||
| p 2 | 0.935772 | 0.00681 | 137.36 | <0.0001 | |||||
| 6 | Pp | b 1 | 0.044495 | 0.00170 | 26.15 | <0.0001 | 1.18 | 0.99 | 1.66 |
| b 2 | -9.225630 | 1.31600 | -7.01 | <0.0001 | |||||
| b 3 | 41.266710 | 4.85080 | 8.51 | <0.0001 | |||||
| p 1 | 1.001470 | 0.00474 | 211.28 | <0.0001 | |||||
| p 2 | 0.898239 | 0.00906 | 99.17 | <0.0001 | |||||
| 6 | Pps | b 1 | 0.035901 | 0.00138 | 26.01 | <0.0001 | 1.00 | 0.99 | 1.73 |
| b 2 | -1.304410 | 0.41420 | -3.15 | 0.0017 | |||||
| b 3 | 11.565630 | 1.57430 | 7.35 | <0.0001 | |||||
| p 1 | 0.978021 | 0.00671 | 145.79 | <0.0001 | |||||
| p 2 | 0.917932 | 0.00803 | 114.28 | <0.0001 |
EE = Error estándar del estimador del parámetro; REMC = Raíz del error medio cuadrático, R 2 adj = Coeficiente de determinación ajustado; DW = Estadístico de Durbin-Watson; Po = Pinus oaxacana; Pd = Pinus douglasiana; Pp = Pinus patula; Pps = Pinus pseudostrobus.
Los errores promedio de la REMC del modelo 6, con la estructura del error oscilaron de 0.79 m (Pd) a 1.18 m (Pp) (Cuadro 4), y explicaron más de 98 % de la varianza del crecimiento en altura dominante. El modelo autorregresivo continuo de segundo orden (CAR2) garantizó la independencia de los residuales en el proceso de ajuste.
En la Figura 3 se observan las curvas de IS obtenidas con la ecuación de Bertalanffy-Richards sin considerar la estructura del error (línea punteada), así como aquéllas en las que se tomó en cuenta la estructura del error CAR2 (línea continua) a una edad de referencia de 40 años, sobrepuestas a los datos experimentales que se utilizaron en su ajuste. Las curvas de IS cubren las diferentes edades y describen la relación entre crecimiento en altura dominante y edad con un comportamiento lógico y biológico (polimorfismo complejo) a edades mayores a los 50 años (Cieszewski, 2003; Álvarez-González et al., 2010). Se aprecia, gráficamente, que las especies estudiadas presentan distintos patrones de crecimiento en altura dominante, lo cual indica que, el manejo de rodales con dominancia de las cuatro especies de pino requiere de esquemas silvícolas distintos (Attis et al., 2015).
En los gráficos de la función de autocorrelación parcial (PACF), obtenida del ajuste de la ecuación Bertalanffy-Richards con 10 retardos (Lags) de los residuales para cada árbol (Figura 4), se observa que se pasa de un correlograma con autocorrelación positiva de segundo orden, a un correlograma sin evidencia de autocorrelación, estimada con el ajuste de la ecuación dinámica con una estructura autocorregresiva de errores continua tipo CAR2; resultados que son similares a los registrados por Vargas-Larreta et al. (2010), Vargas-Larreta. (2013) y Quiñonez-Barraza et al. (2015), para especies forestales en diferentes regiones de México.
Al corregir la autocorrelación, se determina el patrón aleatorio de los residuales alrededor de la línea de cero, con varianza homogénea y sin detectarse una tendencia definida (Figura 5). También, se observa la evolución del sesgo y RCME, y se nota que a partir de la categoría de edad mayor a 50 años, se presenta un aumento considerable en la dispersión y alturas predichas debido, principalmente, a la falta de pares de datos de altura-edad de esas clases de edad.

Conclusiones
La ecuación dinámica de Bertalanffy-Richards es una opción adecuada para modelar el crecimiento en altura dominante e índice de sitio para las cuatro especies de pino estudiadas en la Umafor 2001. Las ecuaciones obtenidas son polimórficas, con múltiples asíntotas e invariantes con respecto a la edad de referencia; además, permiten estimar de manera directa la altura dominante y el índice de sitio a cualquier altura y edad de referencia. El uso de estas ecuaciones coadyuvará a mejorar la estimación del potencial productivo de las unidades de manejo donde dominan P. oaxacana, P. douglasiana, P. patula y P. pseudostrobus y serán de utilidad para seleccionar regímenes de manejo orientados a incrementar la productividad, a través de su incorporación dentro de los programas de manejo forestal.

