











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
Los hongos ectomicorrizógenos (HECM) son componentes importantes de la biota edáfica, ya que tienen un papel preponderante en la movilización de nutrimentos entre el suelo y las plantas, principalmente de nitrógeno y fósforo, así como en el transporte de agua (Smith y Read, 1997). El establecimiento de la asociación ectomicorrícica es fundamental para el buen desarrollo de muchas especies de árboles de zonas templadas, entre las que destacan miembros de la familia Pinaceae, como Pinus spp. y Abies religiosa (Kunth) Schltdl. et Cham. Estos taxa son de gran importancia económica, y sus rodales aportan 2.6 687 millones m3 r (metros cúbicos de madera en rollo) (Semarnat, 2017) de la madera que se comercializa en México procedente de bosques con programas de manejo forestal.
Las poblaciones de HECM son sensibles a cambios en los factores abióticos (la precipitación, la altitud, la exposición); y en las propiedades edáficas como pH, textura, capacidad de retención de humedad, concentración de carbono, sodio, magnesio, calcio, y la capacidad de intercambio catiónico (Martínez-Peña et al., 2012; Taylor et al., 2014; Nadeau y Khasa, 2016; Smith et al., 2017). Dado que la extracción de madera y otras prácticas silvícolas modifican las propiedades físicas y químicas del suelo forestal, como consecuencia del aumento en la insolación, la tasa de evapotranspiración y la temperatura, se espera que estas incidan sobre la composición de la estructura de las comunidades ectomicorrícicas (Zamora-Martínez, 2010), tanto en la emergencia de esporomas como en la presencia de micorrizas en los sistemas radicales (Perry et al., 1984; Wright et al., 1997; Pilz et al., 2003; Valdés et al., 2003; Bonet et al., 2004; Pilz et al., 2006; Valdés et al., 2009; de-Miguel et al., 2014). Por ejemplo, se ha documentado que la compactación y remoción de suelo, producto de la extracción de la madera en plantaciones de Pseudotsuga menziesii (Mirb.) Franco, disminuye su abundancia y diversidad (Amaranthus et al., 1996).
La influencia de las propiedades del suelo en la composición taxonómica y en la estructura funcional de las comunidades ectomicorrizógenas, a nivel de esporomas y de las micorrizas, ha sido sujeto de estudio por diversos autores (Courty et al., 2010; Martínez-Peña et al., 2012). Sin embargo, en México la información relacionada con esa temática ha sido poco abordada, y en general se le refiere de manera colateral, o bien complementaria en investigaciones relacionadas con el efecto de los tratamientos silvícolas en la diversidad fúngica (Valdés et al., 2003; Valdés et al., 2009), así como en trabajos referentes a la distribución potencial de algunos macromicetos (Zamora-Martínez et al., 2014).
En este contexto, se planteó el objetivo de establecer la relación existente entre las propiedades físicas y químicas del suelo y la aparición de los esporomas de especies ectomicorrizógenas en rodales sujetos a diferentes prácticas silvícolas. Se partió de la premisa de que la medición de variables relacionadas directamente con esos factores son un elemento importante para la zonificación de la productividad, en particular de especies no maderables, como es el caso de los hongos, que tienen presencia y valor ecológico, económico y cultural en los ecosistemas forestales de clima templado y templado frío de México.
Materiales y Métodos
El estudio se llevó acabo en el ejido Rancho Nuevo Nanacamila, municipio Zacatlán de Las Manzanas, Puebla, México; entre 20°02'54.24'' y 20°04'30.00'' N, y 98°04'42.24'' y 98°06'38.88'' O; a una altitud de 2 290 m. La vegetación predominante son bosques de Pinus patula Schiede ex Schltdl. et Cham., con presencia, en menor proporción, de P. teocote Schiede ex Schltdl. et Cham., P. leiophylla Schiede ex Schltdl. et Cham., P. rudis Endl., P. pseudostrobus Lindl., Abies religiosa (Kunth) Schltdl. et Cham., Quercus spp. y Arbutus xalapensis Kunth. Las características físicas del lugar varían en función del tratamiento silvícola, por lo que se advierten contrastes importantes de densidad forestal, asociaciones florísticas, espaciamiento, ingreso de luz, pedregosidad, etcétera.
La masa arbórea sujeta a manejo forestal cubre una superficie de 283.13 ha, e incluye la aplicación de tres prácticas según el Método de Desarrollo Silvícola: 1) corta de regeneración (CR), que consiste en dejar en pie únicamente individuos (árboles Padre o árboles semilleros) con cualidades físicas de interés para el aprovechamiento, por ejemplo fustes rectos; 2) corta de liberación (CL), en la cual ocurre la eliminación de los árboles semilleros; y 3) cortas de aclareo, que se aplican para extraer los individuos arbóreos que no cumplen con las características comerciales determinadas en el programa de manejo; en el presente estudio se evaluaron la primera (PCA) y segunda (SCA) corta de aclareo. El ejido cuenta con certificación para su programa de manejo emitido por la Rainforest Alliance (certificado FSC, RA-FM/COC-006372).
Los sitios de muestreo estuvieron conformados por 10 parcelas de 33 × 33 m, distribuidos de la siguiente manera: tres parcelas para los tratamientos correspondientes a las cortas de regeneración y primera corta de aclareo, dos para la segunda corta de aclareo y de liberación, cuya densidad y área basal después de la intervención se muestran en el Cuadro 1.
Cuadro 1 Información dasométrica de las parcelas de muestreo en Rancho Nuevo Nanacamila, municipio Zacatlán de Las Manzanas, Puebla.
| Tratamiento silvícola | Densidad (Indiv. ha -1 ) |
Área basal (m -2 ha -1 ) |
|---|---|---|
| Corta de regeneración | 187.50** | 11.58 |
| Corta de liberación | 625** | 17.55 |
| Primera corta de aclareo | 672.22* | 37.42 |
| Segunda corta de aclareo | 800** | 29.67 |
*Promedio de tres años de intervenidas = 2006, 2007 y 2012; **Año de aplicación = 2009.
Para estimar la riqueza y la abundancia de HECM, durante los meses de julio a noviembre de 2015 y 2016 se recolectaron todos los esporomas de las especies ectomicorrizógenas presentes; para ello, se hicieron recorridos semanales en cada unidad de muestreo (parcela) siguiendo un esquema en zig-zag hasta cubrir la totalidad del área. El material fúngico de cada parcela se separó por especie para contabilizarlo, y se colocó en bolsas de polipapel o en papel encerado. Posteriormente, las muestras se identificaron a partir de sus características macroscópicas (Largent et al., 1984; Pérez-Silva y Herrera, 1991; Phillips, 1991; Rodríguez-Alcalá et al., 2002; Pérez-Moreno et al., 2010; García-Rodríguez et al., 2012). Los ejemplares se depositaron en el Herbario Nacional Forestal “Biól. Luciano Vela Gálvez”: (INIF) del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP). La riqueza (S) estuvo representada por el número total de taxones identificados por tratamiento, en tanto que la abundancia por la cantidad de esporomas recolectados de cada especie.
El efecto que ejercen las prácticas silvícolas en la producción de HECM, en función de las características físicas y químicas del suelo se determinó a partir de un muestreo de suelo efectuado en 2016. El material se recolectó en cinco puntos por parcela, a una profundidad de 0-30 cm para generar una mezcla compuesta. En total se obtuvieron tres muestras por parcela, para un total de 27, y se procesaron en el Laboratorio de Suelos del Centro de Investigación Regional Centro (CIR-Centro) del INIFAP. La determinación de las propiedades físicas y químicas se realizó con base en los lineamientos de la Norma Oficial Mexicana NOM-021-RECNAT-2000 (Semarnat, 2002).
Con la información de campo se efectuó la comparación de la abundancia (número de esporomas) y la riqueza de especies de HECM entre tratamientos silvícolas por año de recolecta, y entre años para cada práctica, mediante pruebas de ANOVA Factorial Entre Sujetos, y el ajuste para múltiples comparaciones de los p-valores con la prueba de Bonferroni (Zar, 2010).
El análisis físico y químico del suelo incluyó 18 variables, a partir de las cuales se evaluó la presencia de distribución normal. En el caso de ausencia de normalidad en los datos, se utilizó la prueba no paramétrica de Kruskal-Wallis para analizar la existencia de diferencias significativas entre los tratamientos (Zar, 2010). En aquellas comparaciones que hubo diferencias significativas, se realizó un análisis de comparaciones pareadas para muestras independientes, con la prueba U de Mann-Whitney (Quinn y Keough, 2002).
Asimismo, se hizo el análisis de correlación de Pearson para determinar la posible relación que asocia los porcentajes y las concentraciones de los elementos físicos y químicos del suelo con la riqueza y abundancia de los HECM. Finalmente, se llevaron a cabo correlaciones para vincular la frecuencia muestreada de las especies registradas en las parcelas y los datos edáficos. Todos los análisis estadísticos se ejecutaron en el paquete estadístico SPSS (IBM, 2013).
Resultados y Discusión
Efecto de los tratamientos silvícolas sobre la abundancia y riqueza de HEMC
El número total de esporomas recolectados en 2015 fue superior al registrado para 2016, con 1 177 y 732 respectivamente. Durante el período de estudio, la mayor abundancia se registró en el tratamiento primera corta de aclareo (Figura, 1). Por su parte, el promedio de la riqueza fue muy similar entre tratamientos, S = 16 para el 2015 y S = 15 para el 2016 (Figura 2).

CR =Corta de regeneración; PCA = Primera corta de aclareo; SCA = Segunda corta de aclareo; CL = Corta de liberación.
Figura 1 Número de esporomas de HECM recolectados en cuatro tratamientos silvícolas durante dos años de muestreo en el ejido Rancho Nuevo Nanacamila, municipio Zacatlán de Las Manzanas, Puebla.
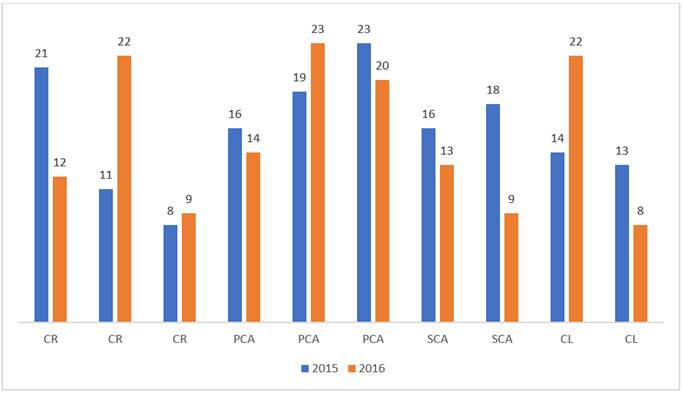
CR = Corta de regeneración; PCA = Primera corta de aclareo; SCA = Segunda corta de aclareo; CL = Corta de liberación
Figura 2 Riqueza de HECM para cuatro tratamientos silvícolas durante dos años de muestreo en el ejido Rancho Nuevo Nanacamila, municipio Zacatlán, Puebla.
La prueba ANOVA Factorial entre Sujetos mostró que en el 2015 el número de esporomas recolectados en el tratamiento corta de regeneración fue significativamente menor (p<0.05) a lo registrado en las otras prácticas silvícolas. Mientras que, en el 2016, las diferencias se detectaron entre la primera corta de aclareo y la segunda corta de aclareo, así como en la corta de liberación (Cuadro 2). No se observaron diferencias cuando se contrastó la riqueza de especies en los distintos tratamientos.
Cuadro 2 Resultados de la prueba ANOVA Factorial Entre Sujetos para la comparación entre número de esporomas, la riqueza (S) y los tratamientos silvícolas en el mismo año de recolecta.
| Tratamiento | Significancia a | |||
|---|---|---|---|---|
| Número de esporomas | Riqueza (S) | |||
| 2015 | 2016 | 2015 | 2016 | |
| CR - 1CA | 0.014* | 0.172 | 0.956 | 1.000 |
| CR - 2CA | 0.030* | 1.000 | 1.000 | 1.000 |
| CR - CL | 0.037* | 1.000 | 1.000 | 1.000 |
| 1CA - 2CA | 1.000 | 0.036* | 0.956 | 0.293 |
| 1CA - CL | 1.000 | 0.043* | 1.000 | 0.833 |
| 2CA - CL | 1.000 | 1.000 | 1.000 | 1.000 |
CR = Corta de regeneración; 1CA = Primera corta de aclareo; 2CA = Segunda corta de aclareo; CL = Corta de liberación. * = Valores estadísticamente significativos (p<0.05). ANOVA Factorial Entre Sujetos: N = 24; g.l. = 3. aAjuste de los p-valores mediante la prueba de Bonferroni.
Cuando la comparación se hizo por tratamiento el número de esporomas recolectados entre 2015 y 2016, la prueba evidenció diferencias significativas para la segunda corta de aclareo y la corta de liberación (Cuadro 3). No hubo diferencias significativas para la riqueza de especies.
Cuadro 3 Resultados de la prueba ANOVA Factorial Entre Sujetos para la comparación entre el número de esporomas y la riqueza (S) obtenida en dos años de recolecta por cada tratamiento silvícola.
| Tratamiento silvícola | Significancia a | |
|---|---|---|
| Núm. de esporomas |
Riqueza (S) |
|
| Corta de regeneración (2015 vs 2016) | 0.940 | 0.809 |
| Primera Corta de Aclareo (2015 vs 2016) | 0.272 | 0.936 |
| Segunda Corta de Aclareo (2015 vs 2016) | 0.001* | 0.104 |
| Corta de Liberación (2015 vs 2016) | 0.002* | 0.872 |
*Valores estadísticamente significativos (p<0.05). ANOVA Factorial Entre Sujetos: N = 24; g.l. = 3. aAjuste de los p-valores mediante la prueba de Bonferroni.
Los resultados evidenciaron que existe una distribución diferencial de la emergencia de esporomas entre las cuatro prácticas silvícolas evaluadas, lo cual se asocia con la intensidad de corta. Resulta congruente, si se toma como base una perspectiva ecosistémica en la que se considera al tratamiento silvícola como un factor de disturbio que modifica la densidad del bosque, la estructura espacial y el valor de sus existencias (Zamora-Martínez, 2010), así como las interacciones bióticas. Por lo tanto, la magnitud del disturbio estará en función del tipo de corta: parcial (aclareos) o de regeneración (total o árboles semilleros); ya que su aplicación altera, en forma diferente, el área basal, la altura dominante, la cobertura arbórea en el rodal, así como otras variables dasométricas que tienen efectos significativos sobre la producción de HECM (Velasco et al., 2010; Bonet et al., 2012; Martínez-Peña et al., 2012). Por otra parte, las acciones ejecutadas durante la extracción de la madera, ya sea manual o mecanizada, impactan las poblaciones fúngicas, toda vez que originan la compactación del suelo y disminuyen la evapotranspiración (de-Miguel et al., 2014).
La corta de regeneración fue el tratamiento silvícola en el cual se registró el menor número de esporomas en los dos años de evaluación (2015 y 2016) (Figura 1). Dicho tratamiento provoca un alto grado de disturbio que afecta las comunidades de hongos asociados, puesto que implica la extracción de prácticamente todo el arbolado (excepto los árboles Padre o semilleros); propicia el aumento de la radiación solar, la temperatura edáfica y la de los estratos de la vegetación herbácea y arbustiva remanente, así como de la que se desarrolla durante los años posteriores a la intervención (seis años para el presente estudio).
Al respecto, Luoma et al. (2004) indican que la producción de HECM disminuye a medida que aumenta la intensidad de corta; aunque, también se ha documentado en bosques de Pinus pinaster Aiton. sujetos a manejo intensivo, que puede favorecer el desarrollo de taxa particulares, como Lactarius gpo. deliciosus (Bonet et al., 2012). Sin embargo, es importante señalar que otros factores ambientales que inciden en la emergencia de esporomas son la distribución e intensidad de la precipitación pluvial (Martínez de Aragón et al., 2007; Velasco et al., 2010).
Por otra parte, existen evidencias de que el área basal (AB) del rodal afecta significativamente la producción de los HECM (Martínez-Peña et al., 2012). En el caso de las parcelas ubicadas en las áreas sometidas a corta de regeneración, a los seis años de la intervención, el AB fue de 11.58 m2 ha-1, que fue inferior a los valores estimados en los otros tratamientos (Cuadro 1, Figura 1). Por ejemplo, para la primera y segunda corta de aclareo las respectivas áreas basales estimadas fueron de 37.42 m2 ha-1 y 29.67 m2 ha-1, registros que superan el intervalo de 15-20 m2 ha-1 documentado para esta variable como valor relacionado con producciones máximas de HECM en bosques de pino (Bonet et al., 2008; Bonet et al., 2010).
Referente a las diferencias observadas en cuanto al número de esporomas entre la primera corta de aclareo y la segunda corta de aclareo, cabe señalar que están sesgadas por la diferencia en el número de parcelas consideradas, aunque lo esperado era registrar una abundancia superior a medida que la masa forestal adquiriera mayor edad y vigor. Una posible explicación radica en el tiempo transcurrido entre el momento de la intervención y el año de muestreo, el cual fue de diez años para la PCA y de seis para la SCA. Después de ejecutar cualquier tipo de corta, selectiva o de regeneración, se modifican las propiedades físicas y químicas de los suelos, lo que se refleja en la pérdida de nutrimentos, en la alteración de la densidad aparente y de la porosidad, así como de su capacidad para retener agua, por citar algunas (Kishchuk et al., 2014).
El período de recuperación puede sobrepasar los 10 años, en función de la intensidad de la corta (Zhou et al., 2015) y estos impactos negativos a nivel edáfico influyen en la abundancia de los HECM, dada la dependencia nutricional que estos organismos tienen de sus hospederos arbóreos, los cuales al haber menos disponibilidad de nutrimentos disminuyen su actividad fotosintética y con ello, la traslocación de carbohidratos hacia los HECM (Nehls, 2008).
Cabe señalar que aún se carece de investigaciones que permitan explicar con precisión el efecto de las prácticas silvícolas en las poblaciones fúngicas presentes en los rodales sujetos a programas de manejo forestal, particularmente, de aquellas especies de hongos que son recolectadas con fines comerciales. Al respecto, Pilz et al. (2006) señalan que la producción de esporomas de Chantarellus spp. disminuye durante el primer año posterior al aprovechamiento maderable, pero seis años después su productividad casi se recupera.
La corta de liberación que consiste en eliminar los árboles de la especie en proceso de crecimiento, pertenecientes a una clase de edad superior, así como los taxa herbáceos y arbustivos (Daniel, 1983) provoca la compactación del suelo, con la consecuente pérdida del inóculo ectomicorríco presente a nivel edáfico y en algunas “plantas refugio” (Wiensczyk et al., 2002; Kennedy et al., 2003; Martínez, 2008), lo que se reflejó en una reducida abundancia de esporomas.
Efecto de las condiciones edáficas sobre la abundancia y riqueza de los HECM
Uno de los retos del sector forestal es aplicar prácticas silvícolas para el manejo sustentable del recurso. Mantener la productividad del suelo resulta clave para este fin, ya que impacta no solo sobre el potencial de conservación de la producción maderable a largo plazo, sino también en otras funciones del ecosistema forestal, como la presencia de la biota asociada, y en particular de los HECM. Al comparar los cuatro tratamientos silvícolas evaluados, se obtuvieron diferencias significativas (p>0.05) para 11 de las variables edáficas consideradas: el porcentaje de materia orgánica, porcentaje de arena, fósforo (P) disponible, potasio (K), calcio (Ca), magnesio (Mg), sodio (Na), hierro (Fe), zinc (Zn), cobre (Cu) y pH (Cuadros 4 y 5).
Cuadro 4 Valores medios de las propiedades físicas y químicas de los suelos con diferencias significativas en el análisis pareado U de Mann Whitney.
| Variable edáfica / Tratamientos silvícolas |
CR | CL | 1CA | 2CA |
|---|---|---|---|---|
| Materia orgánica (%) | 38.25 | 8.15 | 46.46 | 8.04 |
| Arena (%) | 81.59 | 84.01 | 86.29 | 82.26 |
| P bray (mg L-1) | 1.11 | 0.77 | 2.11 | 1.36 |
| K (mg L-1) | 306.18 | 352.80 | 196.6 | 373.79 |
| Mg (mg L-1) | 197.47 | 225.46 | 162.8 | 235.39 |
| Na (mg L-1) | 49.15 | 13.86 | 28.64 | 17.56 |
| Fe (mg L-1) | 84.99 | 41.20 | 39.26 | 29.22 |
| Zn (mg L-1) | 1.89 | 0.37 | 0.65 | 0.72 |
| Cu (mg L-1) | 0.25 | 0.26 | 0.39 | 0.35 |
| pH | 5.45 | 5.76 | 5.65 | 5.93 |
| Ca (mg L-1) | 1 133.08 | 1 236.70 | 1 093.18 | 1 594.20 |
CR = Corta de Regeneración; CL = Corta de Liberación; 1CA = Primera Corta de Aclareo; 2CA = Segunda Corta de Aclareo.
Cuadro 5 Resultados de la prueba de Kruskal-Wallis para comparar las diferencias de las variables físicas y químicas del suelo entre los tratamientos silvícolas.
| Variables fisicoquímicas del suelo | Significancia |
|---|---|
| Materia Orgánica (%) | <0.001* |
| N inorgánico (mg L-1) | 0.857 |
| P bray (mg L-1) | 0.027* |
| K (mg L-1) | 0.001* |
| Ca (mg L-1) | 0.032** |
| Mg (mg L-1) | 0.017* |
| Na (mg L-1) | 0.007* |
| Fe (mg L-1) | <0.001* |
| Zn (mg L-1) | 0.001* |
| Mn (mg L-1) | 0.171 |
| Cu (mg L-1) | 0.004* |
| pH | 0.007* |
| Punto de saturación (%) | 0.071 |
| Capacidad de campo (%) | 0.071 |
| Punto de marchitez permanente (%) | 0.071 |
| Arena (%) | 0.05* |
| Arcilla (%) | 0.585 |
| Limo (%) | 0.060 |
*Resultados que tuvieron una diferencia estadísticamente significativa (p>0.05); **Elemento con diferencia significativa con la prueba Kruskal-Wallis, pero sin ninguna comparación pareada (U de Mann-Whitney) significativa; N = 28; g.l. = 54 para cada caso
La comparación entre pares de tratamientos para las propiedades edáficas evidenció las mismas correlaciones; de ellas, sobresalieron como las más relevantes la materia orgánica y el porcentaje de arena (Cuadro 6).
Cuadro 6 Comparaciones pareadas (U de Mann Whitney) entre tratamientos respecto a los elementos físicos y químicos del suelo que registraron diferencias significativas mediante la prueba Kruskal-Wallis.
| Propiedades edáficas /Comparación entre tratamientos |
CR - 1CA | CR - 2CA | CR - CL | 1CA - 2CA | 1CA - 2CA | 2CA - CL |
|---|---|---|---|---|---|---|
| Materia orgánica (%) | 1.000 | 0.120 | 0.039* | 0.004* | 0.012* | 1.000 |
| Arena (%) | 0.049 | 1.000 | 1.000 | 0.372 | 1.000 | 1.000 |
| P bray (mg L-1) | 0.380 | 1.000 | 1.000 | 1.000 | 0.046* | 0.176 |
| K (mg L-1) | 0.589 | 0.504 | 0.467 | 0.008* | 0.005* | 1.000 |
| Ca** (mg L-1) | 1.000 | 0.083 | 1.000 | 0.057 | 0.823 | 1.000 |
| Mg (mg L-1) | 1.000 | 0.571 | 0.372 | 0.094 | 0.046* | 1.000 |
| Na (mg L-1) | 1.000 | 0.346 | 0.014* | 0.673 | 0.037* | 1.000 |
| Fe (mg L-1) | 0.007* | <0.001* | 0.072 | 0.992 | 1.000 | 0.615 |
| Zn (mg L-1) | 0.056 | 0.526 | 0.001* | 1.000 | 0.628 | 0.351 |
| Cu (mg L-1) | 0.006* | 1.000 | 1.000 | 0.467 | 0.022* | 1.000 |
| pH | 1.000 | 0.024* | 0.029* | 0.379 | 0.488 | 1.000 |
CR = Corta de Regeneración; 1CA = Primera Corta de Aclareo; 2CA = Segunda Corta de Aclareo; CL = Corta de Liberación. *Resultados que tuvieron una diferencia significativa (p<0.05) mediante el análisis pareado de la U de Mann Whitney. N=6 y g.l. = 2 para cada caso. **Elemento con diferencia significativa con la prueba Kruskal-Wallis, pero sin ninguna en la correlación significativa con la prueba U de Mann-Whitney.
El efecto que tiene la cantidad y calidad de la materia orgánica en los suelos forestales se basa en que es un componente cuantitativo y funcional del ecosistema que desempeña un papel importante en el ambiente físico y químico. Su descomposición integra elementos de fertilidad y toxicidad al sustrato, intercepta la luz y sirve para regular los gradientes térmicos entre el suelo y la atmósfera; asimismo, aumenta la captación de la lluvia, pero reduce su evaporación. Es por eso, que sirve de barrera física para muchos procesos y elementos químicos (Facelli y Pickett, 1991).
En general, se ha documentado que la materia orgánica disminuye en función de la intensidad de la corta (Zhou et al., 2015). Ello concuerda con las diferencias significativas entre la primera corta de aclareo y la corta de liberación (Cuadro 5); no así entre esta última y la corta de regeneración, probablemente debido a que el material residual de la extracción de la madera (ramas, tocones, corteza, acículas) se deja en el sitio de aprovechamiento, mientras que en las parcelas donde se aplicó la corta de liberación, se retira.
Al contrastar los tratamientos silvícolas aplicados, las diferencias significativas para el pH se registraron entre la corta de regeneración con las de liberación y la segunda corta de aclareo, cuyos valores fueron más altos; aunque los registros resultaron muy similares (Cuadros 4 y 6).
Las diferencias significativas entre las cortas de aclareo para las concentraciones de K y P coinciden con lo citado en la literatura respecto a que dichos elementos disminuyen con la apertura del dosel (Cuadro 1) (Kishchuk et al., 2014; Zhou et al., 2015). Sin embargo, al comparar el tratamiento correspondiente a la primera corta de aclareo con la de liberación, el valor de K fue superior en esta última (Cuadro 4), aun cuando su AB es menor (Cuadro 1). Una posible causa es la presencia de abundantes arbustos y herbáceas que aportan hojarasca al piso forestal, el cual tiene un pH más alto y mayor concentración de cationes básicos, como el K (Kishchuk et al., 2014).
Los principales factores edáficos que se relacionaron con la abundancia de esporomas fueron el porcentaje de materia orgánica, de arena y limo. La textura del suelo, determinada por el porcentaje de limo y arena, afecta la riqueza de especies y la emergencia de esporomas de los HEMC (Martínez-Peña et al., 2012; Taylor et al., 2014). Los suelos con porcentajes altos de arena disminuyen la retención de agua, en tanto que los suelos limosos la aumentan y con ello se favorece la producción de esporomas. Los resultados que aquí se documentan difieren de lo antes citado, ya que la correlación fue negativa entre el porcentaje de limo y la abundancia de los HECM; mientras que para el de arena fue positiva, al igual que el contenido de materia orgánica (Cuadro 7), la cual desempeña el papel de amortiguador de la evapotranspiración y ayuda a mantener la temperatura edáfica, acciones que, probablemente contribuyeron a disminuir la pérdida de agua, lo que pudo favorecer la emergencia de los esporomas.
Cuadro 7 Análisis de correlación de Pearson entre la riqueza y abundancia de los HECM y los resultados del análisis físico y químico del suelo.
| Fertilidad del suelo | Riqueza (S) | Abundancia | ||
|---|---|---|---|---|
| Coeficiente de Correlación de Pearson |
p <0.05 | Coeficiente de Correlación de Pearson |
p <0.05 | |
| Materia Orgánica Muerta (%) | 0.351 | 0.067 | 0.634* | <0.001* |
| N inorgánico (mg L-1) | -0.142 | 0.471 | -0.083 | 0.674 |
| P bray (mg L-1) | -0.158 | 0.421 | 0.020 | 0.918 |
| K (mg L-1) | -0.244 | 0.211 | -0.640* | <0.001* |
| Ca (mg L-1) | -0.408* | 0.031* | -0.564* | 0.002* |
| Mg (mg L-1) | -0.231 | 0.237 | -0.647* | <0.001* |
| Na (mg L-1) | 0.057 | 0.775 | 0.127 | 0.521 |
| Fe (mg L-1) | -0.007 | 0.971 | -0.138 | 0.484 |
| Zn (mg L-1) | -0.214 | 0.274 | -0.190 | 0.332 |
| Mn (mg L-1) | -0.475* | 0.011* | -0.397* | 0.036* |
| Cu (mg L-1) | 0.089 | 0.653 | 0.275 | 0.157 |
| pH | -0.111 | 0.572 | -0.197 | 0.315 |
| Punto de Saturación (%) | 0.248 | 0.203 | 0.292 | 0.132 |
| Capacidad de Campo (%) | 0.248 | 0.204 | 0.292 | 0.132 |
| Punto de marchitez permanente (%) | 0.248 | 0.204 | 0.294 | 0.130 |
| Arena (%) | 0.277 | 0.154 | 0.404* | 0.033* |
| Arcilla (%) | 0.138 | 0.485 | 0.097 | 0.625 |
| Limo (%) | -0.335 | 0.082 | -0.453* | 0.015* |
*Resultados que tuvieron una diferencia estadísticamente significativa (p<0.05).
Respecto a los nutrimentos minerales, los elementos que exhibieron diferencias significativas fueron K, Ca, Mg y Mn. El Ca y Mn se correlacionaron negativamente con la riqueza de especies (Cuadro 7); lo anterior se explica por la afinidad que, en general, tienen los HECM por los ambientes ácidos predominantes en los bosques de coníferas. Por lo tanto, al aumentar el contenido de Ca, el sustrato se alcaliniza, de tal manera que resulta menos propicio para el desarrollo de los hongos y en consecuencia disminuye el número de especies fúngicas.
El análisis de Pearson mostró una correlación positiva entre la abundancia de esporomas, el porcentaje de materia orgánica y el porcentaje de arena; en contraste, para K, Mg y el porcentaje de limo, la correlación fue negativa. Por otra parte, tanto la riqueza de especies como la abundancia de esporomas se correlacionaron negativamente con el Ca y el Mg (Cuadro, 7).
En particular, Taylor et al. (2014) destacan la existencia de una variación a escala fina en los nichos-hongos que se relaciona, principalmente, con la evolución del suelo, ya que la textura está muy ligada con el contenido de microelementos asimilables. Además de que el valor del cociente entre los solubles y los totales disminuye a medida que aumenta el contenido de elementos gruesos en la textura del suelo.
Respecto al pH, es un factor importante para la asimilación de los macro y microelementos; el aumento en el pH reduce la solubilidad y absorción de algunos de ellos, como el cobre, hierro, zinc (Cuadros 4 y 7) y, muy marcadamente, del manganeso. Sin embargo, el incremento en el pH aumenta la solubilidad y absorción del molibdeno (Mo), aunque la concentración de este último no fue significativa para la abundancia y riqueza de los HECM (Cuadro 7).
En relación con la producción de HECM existen registros del efecto significativo del pH sobre el rendimiento de ciertas especies fúngicas, en particular; por ejemplo, Lactarius gpo. deliciosus, que muestra marcada preferencia por los suelos ácidos (Bonet et al., 2012). En la investigación llevada a cabo en el ejido Rancho Nuevo Nanacamila, Lactarius indigo (Schwein.) Fr. y Boletus aestivalis (Paul) Fr. solo se recolectaron en el tratamiento correspondiente a corta de liberación, cuyo pH fue de 5.76; Tylopilus sp. se registró, igualmente, en sitios con valores menos ácidos (5.93), en rodales sujetos a segunda corta de aclareo.
Investigaciones recientes señalan que Boletus loyo Phil. ex Speg. se desarrolla en suelos con altos contenidos de materia orgánica, concentraciones de medias a bajas de N, P y Ca y valores elevados de K, Mg, Fe, Mn, Zn y Cu, los cuales favorecen la producción de sus esporomas (Pereira et al., 2016). Sun et al. (2017) resaltan la importancia de diversos elementos, entre ellos K y Mg, en la emergencia de los esporomas de HECM en una asociación de Cedrus deodara (Roxb.) G. Don, lo que coincide con la información aquí documentada. Cabe señalar que en el presente estudio tanto el K como el Mg tuvieron, en general, correlaciones negativas con la abundancia de HECM (Cuadro 7), pero positivas entre tratamientos (Cuadro 6).
Los resultados son congruentes con lo antes expresado, al existir diferencias significativas en la disminución de algunos de los elementos minerales del suelo, en función de las prácticas silvícolas evaluadas; por ejemplo, en la primera corta de aclareo disminuyen en forma notable K, Ca y Mg; y en la corta de liberación las menores concentraciones corresponden a P, Na y Zn (Cuadro 4). Cabe destacar que los nutrimentos citados tuvieron correlaciones negativas con la abundancia de esporomas, excepto el fósforo disponible y el sodio (Cuadro 7).
A partir de lo antes expuesto, en particular del porcentaje de materia orgánica como un elemento indicador de la calidad del suelo, y las diferencias estadísticamente significativas obtenidas para dicha variable es evidente que cualquiera de las prácticas silvícolas altera su tasa anual de producción, lo que incide sobre el resto de las propiedades físicas y químicas del suelo, así como en la abundancia y riqueza de los taxa de HECM (Martínez-Peña et al., 2012).
Conclusiones
Las prácticas silvícolas corta de regeneración, corta de liberación y cortas de aclareo generan cambios en la composición y estructura de la masa arbórea que incide en la abundancia y riqueza de los taxa de hongos ectomicorrizógenos. Esto es particularmente notorio en la corta de regeneración; es decir, en sitios donde la extracción de la masa arbórea es más intensiva. Las diferencias registradas entre las cortas de aclareo responden al tiempo transcurrido entre el año de intervención y el de muestreo, el cual fue mayor en la primera corta de aclareo y correspondió con una abundancia superior de esporomas.
Las propiedades físicas y químicas del suelo, particularmente el porcentaje de materia orgánica y de arena registraron una correlación positiva respecto a la abundancia de esporomas, a diferencia de las concentraciones de K, Ca, Mn y Mg. En cambio, entre tratamientos silvícolas, las diferencias significativas fueron negativas con un efecto directo en la abundancia y riqueza de los HECM.
Lactarius indigo, Boletus aestivalis y Tylopilus sp solo se registraron en rodales con valores de pH menos ácidos (5.76 a 5.93), que corresponden a las cortas de aclareo y de liberación.
Es importante realizar estudios más detallados, a nivel de especie, para establecer las respuestas fúngicas específicas a los factores edáficos, ya que los requisitos nutricionales de los HECM varían entre taxa.

