











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
En México existe la mayoría de los tipos de vegetación registrados a nivel mundial, además de una amplia gama de climas, mismos que condicionan los procesos de formación del suelo, así como la diversidad e interacción entre las especies (Villaseñor, 2003; Hernández et al., 2011; Dzib-Castillo et al., 2014).
El estado de Oaxaca reúne la mayor biodiversidad, y en gran parte de su territorio predomina el tipo de vegetación de selva baja caducifolia (SBC) (López et al., 2005); estos ecosistemas están siendo modificados, principalmente, por actividades antrópicas que afectan a las poblaciones de Bursera spp., las cuales son importantes en el ámbito biológico y económico, al ser utilizadas como materia prima para la obtención de resinas aromáticas, aceites esenciales y para la elaboración de artesanías.
Los recursos naturales: suelo, agua, flora y fauna no son aprovechados de manera sustentable en muchas partes del país. En el caso particular de la selva baja espinosa y de especies de Bursera, en particular, sus poblaciones están disminuyendo, pues la regeneración natural mediante semillas no contrarresta los niveles de aprovechamiento. Se ha documentado que los árboles de Bursera simplex Rzed. & Calderón tienen porcentajes elevados de semillas vanas, condición generada por posibles cruzamientos con B. biflora Rzed. & Calderón; a esto hay que agregar la reducción notable del número de árboles en etapa reproductiva, lo que induce a la endogamia, que repercute negativamente en las plantas al reducir su viabilidad (Cuevas y Abarca, 2006; Rzedowski y Calderón, 2006; Nora et al., 2011).
Bursera simplex es endémica de Oaxaca, se distribuye únicamente en la porción media de la cuenca del río Tehuantepec (Rzedowski y Calderón, 2006); donde es un componente importante del bosque tropical caducifolio y se le utiliza, cotidianamente, para extraer resina aromática con fines ceremoniales. Tiene potencial para otros usos, a partir de los hallazgos de Gigliarelli et al. (2015) quienes identificaron su contenido de ácido morólico y 3-epi-lupeol, esqueletos de lupano y olaneano r, que de acuerdo con Cano (2013) son componentes utilizados como antitumorales, antiinflamatorios, antiVIH, antimicrobianos, hepa y cardioprotectores, analgésicos, antimicóticos, antiquimiopreventivos, entre otros.
La diversidad estructural de las comunidades vegetales es uno de sus aspectos más relevantes, al estar relacionada con el hábitat de diversos animales y plantas; de igual manera lo está con la estabilidad frente a distintos factores bióticos y abióticos, así como con los beneficios directos (productos) e indirectos (fijación de carbono, paisaje, protección del suelo, entre otras) (Del Río et al., 2003). Por ello, es importante conocer la estructura del ecosistema, que está influenciada, principalmente, por el tipo, número, ordenamiento espacial y temporal de los elementos que lo constituyen (Corral et al., 2005; Ledo et al., 2012), los cuales se pueden medir mediante índices estructurales y variables morfológicas (diámetro, altura, área basal, densidad). La relación de tamaños entre los árboles es importante, debido a que las especies mostrarán diferencias morfológicas que dan lugar a contrastes estructurales (Castellanos et al., 2008; Jiménez et al., 2009).
La variabilidad morfológica de los árboles resulta de diferencias intraespecíficas, pero también al efecto de factores biológicos y físicos, mismos que tienen una estrecha correlación con la reproducción debido a que cualquier factor ambiental que influya en el crecimiento lo hará, de igual manera, con el desempeño reproductivo (Enríquez et al., 2004).
Además, las semillas en una especie pueden variar en tamaño entre individuos y poblaciones, por diferencias genéticas o por diferencias en la historia de vida de cada planta. Desde este punto de vista, Alba y Márquez (2006) mencionan que el potencial de producción de semillas de un taxon forestal está influenciado por las condiciones ambientales de un sitio determinado, así como por sus características fenotípicas, las cuales permiten evaluar y ponderar el potencial para su manejo; o bien, seleccionar individuos sobresalientes dentro de una población determinada (Balcorta y Vargas, 2004; Cerón y Sahagún, 2005); cuando se pretende propagar una especie arbórea con fines de reforestación, es importante manejar criterios rigurosos de la selección de los árboles en los que se colectarán semillas, ya que de ello depende que en la siguiente generación de individuos se logre ganancia genética (Vallejos et al., 2010). Por lo anterior, el objetivo del presente estudio consistió en evaluar el efecto de la estructura arbórea y variables de sitio sobre el peso y características morfológicas de los frutos y semillas de Bursera simplex.
Materiales y Métodos
Área de estudio
El estudio se llevó a cabo durante 2016 en Santa María Zoquitlán, municipio del mismo nombre, una comunidad de los Valles Centrales de Oaxaca, ubicada a 16°34’ N y 96°21’ O, a una altitud media de 1 080 m. La vegetación predominante donde se distribuye B. simplex es la selva baja caducifolia, especie endémica de la cuenca del Río Tehuantepec; los géneros que comparten el hábitat son Cephalocereus, Cercidium, Lysiloma, Ceiba, Neobuxbaumi, Escontria, Acacia, Beaucarnea, entre otras. El clima predominante es el BSo(h')w” (w) i´g (Acosta et al., 2003).
Método de muestreo y registro de datos
Se realizaron muestreos dirigidos con la finalidad de identificar árboles de la especie de interés con características fenotípicas sobresalientes, de apariencia sana y que además presentaran frutos, este último fue el principal criterio para su selección. Durante octubre-diciembre de 2015 se lograron reconocer 27 árboles en el paraje denominado El Paredón, ubicados en un intervalo altitudinal de 1 078-1 406 m; los individuos se identificaron con números progresivos y fueron tomados como punto central de referencia para establecer 23 unidades muestrales (UM) de 100 m2.
En cada sitio de muestreo, se contabilizaron todas los taxa presentes y con el apoyo de una cinta métrica Fiber-glass de 1.52 m y una cinta métrica de 5 m (marca Cadena modelo MGA5020) se midieron y registraron las variables morfológicas: diámetro al pie (DP, cm), altura total (AT, m) y diámetro de copa (DC, m) con el que se calculó el área de copa (AC= 1/4 × πDC2, m2).
En las unidades muestrales establecidas con un GPS Garmin DriveTM, se otuvieron las variables del sitio:exposición, altitud (m) y pendiente (%) con un clinómetro Suunto Finland Pm5/360pc.
De cada uno de los ejemplares de Bursera seleccionados se recolectó parte de los frutos que presentaban, los cuales se introdujeron en bolsas de papel y se trasladaron a las instalaciones del Instituto Tecnológico del Valle de Oaxaca (ITVO).
Se tomó una muestra aleatoria de 50 frutos de cada árbol, a los que se les midió con un vernier Pretul de 0.1 cm de precisión, su diámetro polar (Dp, mm) y su diámetro ecuatorial (DE, mm); se calculó el coeficiente de forma (CF=DE/Dp) y se pesaron (g) de manera individual con una balanza analítica Ohaus Adventurer™ con capacidad máxima de 210 g y precisión de 0.1 g, para obtener el número de frutos kg-1. Posteriormente, para su secado se colocaron bajo radiación solar durante cuatro días; paso seguido, se separó la semilla de los tejidos del fruto, para después medir su diámetro polar y ecuatorial; por último, se les pesó para obtener el número de semillas kg-1.
Con los datos de las variables morfológicas se calcularon índices estructurales y se realizaron histogramas de frecuencia, para ello se utilizaron categorías diamétricas comerciales de 5 cm y categorías de altura a intervalos ≤1.5, 1.5 - 3, 3.1 - 4.5, 4.6 - 6 y >6 (en metros). Se determinaron tres índices de valoración estructural (Zarco et al., 2010; Santiago, 2013):
Donde:
IVI = Índice de valor de importancia
DMr = Dominancia relativa o área basal relativa (ABr)
Dr = Densidad relativa
Fr = Frecuencia relativa
IVF = Índice de valor forestal
DPr = Diámetro al pie relativo (cm)
Ar = Altura relativa (m)
ACr = Área de copa relativa (m2)
IC = Índice de complejidad
s = Número de especies
d = Densidad de árboles mayores a 2.5 cm de DP
b = Área basal (m2)
h = Altura (m) de los tres árboles dominantes
A= Área de la muestra (m2)
Manejo y análisis de datos
En los datos de frutos, semillas y árboles se verificaron los supuestos de normalidad y homogeneidad de varianzas, mediante las pruebas de Shapiro-Wilk y Bartlett. Las variables que los cumplieron se sometieron a un análisis de varianza con un diseño completamente aleatorizado y se realizaron comparaciones de medias aplicando la prueba de Duncan (α = 0.05), en la que se usó como variable clasificatoria el árbol de procedencia, la exposición, altitud y pendiente; para las dos últimas se formaron tres categorías de manera individual.
Para la rutina de análisis se empleó el programa de cómputo SAS (2004), con el procedimiento GLM. En el caso de las variables que no cumplieron con los supuestos de normalidad y homogeneidad de varianzas, se analizaron mediante la prueba de Kruskal Wallis con el procedimiento NPAR1WAY Wilcoxon, en el paquete estadístico SAS (SAS, 2004).
Resultados y Discusión
Estructura vertical y horizontal
Las diferencias de tamaño de árboles de una misma especie ubicados en sitios de exposición diferente se pueden atribuir a que la magnitud de crecimiento es función de las potencialidades definidas por el genotipo, el ambiente y la interacción de ambas. Ramírez y Rodríguez (2004) comentan que la diferencia del crecimiento de los árboles se atribuye a que en el hemisferio norte la exposición sur es la que recibe mayor cantidad de radiación solar. En este sentido, Ortiz-Pulido y Pavón (2010) indican que la exposición del sitio y la cantidad de radiación solar incidente influye en la diferenciación sexual de los individuos de Bursera, ya que los árboles en sitios orientados al este corresponden con más frecuencia al sexo masculino, y en los grupos orientados al oeste al sexo femenino.
Sánchez y López-Mata (2003) definen a la altitud como un factor físico que influye sobre las condiciones climáticas, a través de variables como la temperatura y la precipitación, que tienen, a su vez, un efecto directo sobre el crecimiento vegetal y están correlacionadas de forma específica en cada sitio. Martínez et al. (2013) mencionan que se han determinado relaciones entre el crecimiento de coníferas y los factores de profundidad de suelo, altitud, pendiente y exposición. Rubio et al. (2011), al estudiar la vegetación de Quercus, atribuyeron las diferencias observadas al estado sucesional vegetal y a que las especies sucesionales tempranas producen semillas más pequeñas.
Las alturas máximas en las unidades muéstrales (UM) fueron de 6.60 m; con fines descriptivos de la vegetación, se identificaron dos estratos: inferior y superior. El estrato inferior está constituido, principalmente, por individuos ≤1.5 m, reunidos, en su mayoría, en la exposición oeste (1 057 árboles ha-1) y 326 individuos ha-1 mayores a 1.5 m. En los sitios ubicados en la exposición suroeste, se contabilizó la menor densidad de ejemplares arbóreos en el estrato inferior (209 individuos ha-1). Los individuos de B. simplex conforman el estrato superior, con alturas ≥ 2 m (Figura 1a).
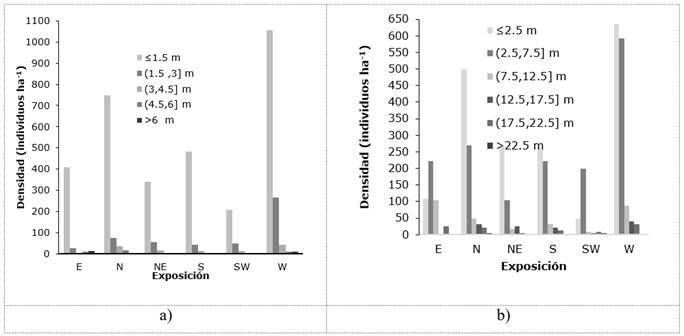
Figura 1 Frecuencia de individuos de acuerdo con sus categorías de altura (a) y diámetro (b) en diferentes exposiciones.
Gutiérrez et al. (2011), en la vegetación de selva baja caducifolia de tres localidades del estado de Yucatán, registraron datos similares, mismos que obtuvieron de agrupaciones con individuos de 1.5 a 4.4 m. Dzib-Castillo et al. (2014), al referirse a la vegetación en las comunidades de Pomuch y San José Carpizo en Campeche, calcularon alturas de 6.3 m en selva baja caducifolia y diámetros de 6.9 cm. En una selva baja caducifolia de Xmatkuil, Yucatán, Gutiérrez y Zamora (2012) midieron individuos de 4 a 12 m de altura y Castillo et al. (2007) describieron tres estratos arbóreos de 1 a 7 m en la vegetación del estado de Veracruz.
En el presente trabajo, en la estructura horizontal se cuantificó una mayor densidad de individuos con diámetros < 2.5 cm en todas las exposiciones, seguidos de los diámetros superiores a 2.5 e inferiores a 7.5 cm, muy relacionadas con las alturas que fueron más pronunciadas en la exposición oeste (Figura 1b); esto se explica por la frecuencia de individuos con diámetros superiores, que disminuye a medida que alcanzan dimensiones más grandes, lo que supone que siempre las clases de diámetro menores serán más abundantes.
Los índices estructurales (IVI, IVF, IC) mostraron diferencias estadísticas significativas (p≤ 0.0001), en su efecto sobre los rasgos morfológicos de los frutos (Cuadro 1). Dichas diferencias se deben, principalmente, a que la estructura está determinada por características morfológicas de los árboles situados en una zona específica, así como por la densidad y la competencia, lo cual implica relaciones dinámicas entre individuos para competir por nutriments o para obtener mejor desarrollo de sus estructuras. Por ello, no siempre una mayor complejidad representará la estabilidad más alta del ecosistema; además, el arreglo espacial de los individuos determina la morfología particular de la planta (Del Río et al., 2003; Perreta y Vegetti, 2005). Alvis (2009) y Zarco et al. (2010) indican que el IVI revela el peso ecológico de una especie dentro de una población.
Cuadro 1 Variables morfológicas de fruto-semilla en relación a los índices estructurales.
| Índice | Variable | |||||
|---|---|---|---|---|---|---|
| Peso fruto (g) |
Cf fruto | Frutos kg -1 |
Peso semilla (g) |
CF semilla |
Semilla kg -1 |
|
| IVF | 673.04** | 585.16** | 637.04** | 656.56** | 259.56** | 256.56** |
| IVI | 695.86** | 690.44** | 695.86** | 802.1** | 282.65** | 802.1** |
| IC | 644.34** | 588.37** | 644.34** | 650.17** | 241.52** | 650.17** |
IVF = Índice de Valor Forestal; IVI = Índice de Valor de Importancia; IC = Índice de Complejidad; CF = Coeficiente de Forma; ** Altamente significativos (Duncan, 0.05).
Características de frutos y semillas
Los frutos y semillas recolectados de 27 árboles de Bursera simplex presentaron diferencias estadísticas significativas en el peso, coeficiente de forma (CF) y en el número de semillas kg-1 (p ≤ 0.0001) en función del árbol de procedencia. La mayor heterogeneidad (CV >65 %) se estimó para el peso de la semilla y número de semillas kg-1; la menor variación se registró para el CF del fruto y el CF de las semillas (CV=7.42 y 13.36 %, respectivamente) (Cuadro 2).
Cuadro 2 Resumen del análisis de varianza de variables morfológicas de frutos y semillas de Bursera simplex Rzed. & Calderón.
| Variable | Fuente de variación |
Grados de libertad |
Cuadrados medios |
Coeficiente de variación (%) |
Desviación estándar |
|---|---|---|---|---|---|
| Peso del fruto | Árbol | 26 | 0.209** | 18.72 | 0.05 |
| Rep | 49 | 0.014** | |||
| Error | 1 274 | 0.003 | |||
| Total | 1 349 | ||||
| Coeficiente de forma del fruto | Árbol | 26 | 0.181** | 7.92 | 0.05 |
| Rep | 49 | 0.008ns | |||
| Error | 1 274 | 0.003 | |||
| Total | 1 349 | ||||
| Peso de la semilla | Árbol | 26 | 0.009** | 65.13 | 0.03 |
| Rep | 49 | 0.001** | |||
| Error | 1 274 | 0.0009 | |||
| Total | 1 349 | ||||
| Coeficiente de forma de la semilla | Árbol | 26 | 0.129** | 13.36 | 0.09 |
| Rep | 49 | 0.008ns | |||
| Error | 1 274 | 0.008 | |||
| Total | 1 349 | ||||
| Número de frutos (kg-1) | Árbol | 26 | 40062159 | 20.55 | 724.24 |
| Rep | 49 | 2527007 | |||
| Error | 1 274 | 524537 | |||
| Total | 1 349 | ||||
| Número de semillas kg-1 | Árbol | 26 | 1955037863** | 22.21 | 5354.54 |
| Rep | 49 | 103220988** | |||
| Error | 1 274 | 28671108 | |||
| Total | 1 349 |
Rep = Repetición; **Altamente significativos (Duncan, 0.05).
Del árbol 5 se seleccionaron los frutos de tamaño y peso superior (0.42±0.09 g), por lo que la cantidad de frutos kg-1 es baja (2456±571.46); dichas magnitudes fueron significativamente diferentes (Duncan, 0.05) a los tamaños y pesos de los frutos de los árboles 3, 9 y 16, que pesaron 0.18±0.03 g; y la cantidad de frutos kg-1 fue de 5 743.7±1 138.21, 5 566±998.51 y 5 743.7±1 138.21, respectivamente.
Dada la relación diámetro ecuatorial - diámetro polar, el CF en fruto de los árboles 1, 15, 19 y 23 fue superior (0.81±0.06 cm) al igual que en el árbol 10 (0.81±0.05 cm); esto implica frutos de forma casi esférica.
El mayor CF en las semillas se obtuvo en los frutos del árbol 25 (0.79±0.01 cm), que fue estadísticamente diferente (p = 0.0001). El individuo 21 presentó las semillas de mayor peso (0.08±0.13), que producen 16 179±5 036 semillas kg-1, mientras que las del árbol 3 fueron más pequeñas, por lo que se tienen 35 489±10 209 semillas kg-1 (Cuadro 3).
Cuadro 3 Frutos-semillas de 27 árboles de Bursera simplex Rzed. & Calderón de Santa María Zoquitlán, Oaxaca.
| Árbol | Variable y significancia | |||||
|---|---|---|---|---|---|---|
| Peso de fruto (g) (0.0001) |
Coeficiente de forma de fruto (cm) (0.0001) |
Frutos kg
-1
(0.0001) |
Peso de semilla (g) (0.0001) |
Coeficiente de forma de semilla (0.0001) |
Semillas kg
-1
(0.0001) |
|
| 1 | 0.30±0.05 d | 0.81±0.06 a | 3 439.3±791.20 c | 0.03±0.007 ghi | 0.74±0.12 abc | 29 328±6 694 c |
| 2 | 0.25±0.05 e | 0.70±0.06 e | 4 142.4±991.18 b | 0.04±0.008 defgh | 0.70±0.09 bcd | 22 318± 4 441 ghi |
| 3 | 0.18±0.03 f | 0.78±0.07 bc | 5 743.7±1138.21 a | 0.03±0.030 ghi | 0.68±0.07 ghi | 35 489±10 209 a |
| 4 | 0.30±0.05 d | 0.68±0.04 ef | 3 438.4±704.60 c | 0.03±0.006 hi | 0.67±0.09 ghi | 32 712±7 347 b |
| 5 | 0.42±0.09 a | 0.77±0.04 cd | 2 456.0±571.46 e | 0.06±0.010 bc | 0.75±0.08 abc | 17 283±4 157 klm |
| 6 | 0.35±0.06 c | 0.76±0.05 cd | 2 878.1±544.05 d | 0.05±0.070 bcde | 0.76±0.08 abc | 22 065± 5 000 hi |
| 7 | 0.38±0.06 b | 0.64±0.06 g | 2 678.6±539.12 de | 0.04±0.009 defgh | 0.68±0.13 fgh | 22 585±5 521 ghi |
| 8 | 0.30±0.05 d | 0.80±0.06 ab | 3 430.5±821.21 c | 0.03±0.005 fghi | 0.72±0.08 cdef | 25 476±3 648 ef |
| 9 | 0.18±0.03 f | 0.69±0.09 e | 5 566.4±998.51 a | 0.02±0.005 i | 0.63±0.08 jkl | 36 983±7 395 a |
| 10 | 0.40±0.04 ab | 0.81±0.05a | 2 512.8±292.90 e | 0.03±0.007 fghi | 0.73±0.09 bcde | 26 947±5 694 ed |
| 11 | 0.29±0.07 d | 0.75±0.05 d | 3 615.2±1003.62 c | 0.03±0.007 hi | 0.69±0.09 efg | 32 719±6 556 b |
| 12 | 0.32±0.08 d | 0.66±0.04 fg | 3 279.4±757.19 c | 0.03±0.006 ghi | 0.64±0.09 hijk | 28 658±5 709 cd |
| 13 | 0.30±0.05 d | 0.68±0.05 ef | 3 427.7±686.91 c | 0.04±0.005 fghi | 0.69±0.09 efgh | 24 671±3 465 fg |
| 14 | 0.40±0.05 ab | 0.80±0.06 ab | 2 492.5±376.73 e | 0.04±0.009 efgh | 0.74±0.06 bc | 23 198±5 262 gh |
| 15 | 0.30±0.05 d | 0.81±0.06 a | 3 438.4±704.60 c | 0.03±0.005 ghi | 0.73±0.09 bcde | 28 966±5 515 cd |
| 16 | 0.18±0.03 f | 0.78±0.07 bc | 5 743.7±1138.21 a | 0.07±0.010 b | 0.66±0.07 ghij | 14 964±4 020 m |
| 17 | 0.30±0.05 d | 0.80±0.06 ab | 3 430.5±821.21 c | 0.03±0.005 hi | 0.72±0.07 cde | 31 849±4 752 b |
| 18 | 0.25±0.05 e | 0.70±0.06 e | 4 142.4±991.18 c | 0.06±0.010 bc | 0.63±0.08 ijk | 17 046±4 198 lm |
| 19 | 0.30±0.05 d | 0.81±0.06 a | 3 438.4±704.60 c | 0.06±0.006 bcd | 0.61±0.08 kl | 16 790±2 022 lm |
| 20 | 0.30±0.05 d | 0.68±0.04 ef | 3 438.4±704.60 c | 0.04±0.010 cdefg | 0.59 ±0.08 l | 22 709±6 809 ghi |
| 21 | 0.35±0.06 c | 0.76±0.05 cd | 2 887.1±544.05 d | 0.08±0.13 a | 0.67±0.08 ghi | 16 179±5 036 lm |
| 22 | 0.30±0.05 d | 0.80±0.06 ab | 3 430.5±821.21 c | 0.04±0.007 defgh | 0.72±0.08 cdef | 21 982±3 572 hi |
| 23 | 0.30±0.05 d | 0.81±0.06 a | 3 439.3±791.20 c | 0.05±0.02 bcde | 0.67±0.09 ghi | 19 385±5 940 jk |
| 24 | 0.30±0.05 d | 0.80±0.06 ab | 3 430.5±821.21 c | 0.06±0.001 bcd | 0.69±0.09 defg | 17 512±4 748 kl |
| 25 | 0.32±0.08 d | 0.66±0.04 fg | 3 279.4±757.19 c | 0.04±0.008 defgh | 0.79±0.01 a | 22 127±4 279 hi |
| 26 | 0.30±0.05 d | 0.68±0.05 ef | 3 427.7±686.91 c | 0.05±0.02 bcde | 0.76±0.09 abc | 19 538±8 972 jk |
| 27 | 0.40±0.05 ab | 0.80±0.06 ab | 2 492.5± 376.73 e | 0.05±0.01 cdef | 0.77±0.08 ab | 20 606±5 160 ij |
Media acompañada±desviación estándar. Letras distintas en columnas representan diferencias estadísticas significativas (Duncan, 0.05).
Posiblemente, las variaciones morfológicas en frutos y semillas de los diferentes árboles resultan del tipo de reproducción propia de Bursera simplex, ya que su condición dioica promueve la polinización cruzada, y, por lo tanto, mayor diversidad en la descendencia (Edwards y Sharitz 2000; Enríquez et al., 2004; Rzedowski y Calderón 2006; Magallán et al., 2009). La variabilidad morfológica observada en frutos y semillas podría tener un notable componente genético además del ambiental (Gómez et al., 2010).
Guzmán y Cruz (2014) refieren que el crecimiento del fruto se relaciona con la humedad disponible, ya que cuando esta baja y las temperaturas elevadas, los frutos son de tamaño menor, además de que varía de acuerdo a la posición que ocupan en el árbol; los frutos grandes se desarrollan en la parte superior de la copa por la mayor disponibilidad de azúcares y proteínas producidas en el área fotosintética, en comparación con los de la porción inferior de la copa. Con base en lo anterior, Rubio et al. (2011) señalan que la variación en el tamaño de las semillas es importante debido a que proporciona diversas capacidades para establecerse en sitios con diferentes condiciones.
Gonzáles y Cruz (1995) citan que los frutos con CF cercanos a la unidad poseen diámetros (polar y ecuatorial) homogéneos y son de forma esférica, mientras que los valores que se alejan de ella describen frutos de forma alargada. Las condiciones del sitio, como la altitud, la exposición y la pendiente del terreno en que se desarrolla el árbol influyen en la magnitud de su crecimiento, lo que a su vez incide en las características morfológicas de los frutos y las semillas (p ≤ 0.01). Los frutos de mayor peso 0.34±0.07 g se recolectaron en árboles localizados en sitios con exposición sur; y las semillas de los individuos con exposición suroeste pesaron 0.06±0.07 g; en contraste, las procedentes de sitios con exposiciones norte y noreste pesaron 0.03 g.
El CF de frutos y semillas refleja diferencias estadísticas significativas (p ≤ 0.0002); de esta manera se confirma que los frutos con mayor CF fueron los recolectados de árboles localizados en sitios con exposición este, con 0.81±0.06 cm, lo que contrasta con las semillas procedentes de árboles en exposición sur, cuyo CF fue de 0.72±0.10 cm; las semillas recolectadas de árboles en lugares con exposición suroeste tuvieron un CF mayor (p=0.72±0.09 cm). La cantidad de frutos y semillas kg-1 fue estadísticamente diferente (p=0.0001), pues las semillas de árboles situados en exposición noreste acumularon 4 235±1 338 frutos kg-1, cifra superior a la de la exposición sur, 3 054±787 frutos kg-1; caso similar ocurre en las semillas de árboles con exposición noreste, que dieron un total de 30 124±8 746 (Cuadro 4).
Cuadro 4 Variables evaluadas en función de la exposición, pendiente y altitud.
| Variable de sitio | Variables de fruto-semilla (probabilidad) |
|||||
|---|---|---|---|---|---|---|
| PF (g) (0.0001) |
CFF (cm) (0.0001) |
Frutos kg
-1
(0.0001) |
PS (g) (0.0001) |
CFS (cm) (0.0002) |
Semilla kg
-1
(0.0001) |
|
| Exposición | ||||||
| Sur | 0.34±0.07 a | 0.72±0.10 c | 3 054.6±787.66 d | 0.05±0.01 b | 0.69±0.11 bcd | 20 048.4±5 722.6 d |
| Oeste | 0.33±0.09 ab | 0.75±0.56 b | 3 236.6±1 111.22 dc | 0.05±0.02 b | 0.70±0.11 abc | 22 186.3±6 908.4 c |
| Suroeste | 0.32±0.06 bc | 0.75±0.07 b | 3 248.4±734.85 dc | 0.06±0.07 a | 0.72±0.09 a | 19 233±6 686.4 d |
| Este | 0.30±0.05 c | 0.81±0.06 a | 3 439.3±791.20 c | 0.05±0.02 ab | 0.67±0.09 d | 19 385.1±5 940.5d |
| Norte | 0.27±0.07 d | 0.75±0.09 b | 3 879.8±1 168.08 b | 0.03±0.01 c | 0.68±0.09 cd | 28 146.7±8 076.2 b |
| Noreste | 0.25±0.07 e | 0.76±0.07 b | 4 235.2±1 338.91 a | 0.03±0.01 c | 0.71±0.01 ab | 30 124.4±8 746.8 a |
| Pendiente (%) | ||||||
| ≤25 | 0.30±0.05 a | 0.81±0.06 a | 3 438.4±704.60 b | 0.06±0.006 a | 0.61±0.08 c | 16 789±2 022.8 c |
| (25,40] | 0.31±0.08a | 0.75±0.08 b | 3 421.4±1 131.96 b | 0.04±0.03 b | 0.71±0.10 a | 23 898.8±8 082.0 b |
| >40 | 0.26±0.08 b | 0.71±0.08 c | 4 104.7±1 271.68 a | 0.04±0.01 b | 0.66±0.09 b | 27 040±8 962 a |
| Altitud(m) | ||||||
| ≤1 178 | 0.24±0.07 c | 0.76±0.08 a | 4 441.81±1 375.15 a | 0.03±0.01 b | 0.72±0.10 a | 29 259.4±8 974.6 a |
| (1 178-1 278] | 0.36±0.08 a | 0.74± 0.07 b | 3 531.67±725.56 b | 0.04±0.03 a | 0.72±0.09 a | 23 985.7±7 212.7 b |
| >1 278 | 0.30±0.08 b | 0.75±0.08 ab | 2 940.33±1 128.83 c | 0.04±0.03 a | 0.69±0.10 b | 23 316.9±8 140.1 b |
Media acompañada ±desviación estándar. Letras distintas en columnas representan diferencias estadísticas significativas (Duncan, 0.05); PF = Peso de fruto; DPF = Diámetro polar de fruto; CFF = Coeficiente de forma de fruto; PS = Peso semilla; CFS = Coeficiente de forma de semilla.
Las pendientes mostraron diferencias significativas (p ≤ 0.001); de los árboles ubicados en sitios con 25 y 40 %, se recolectaron los frutos de tamaño mayor, con una media de 0.31 g y la producción de frutos kg-1 es favorable en pendientes >40 %, puesto que presentan un promedio de 4 104 frutos kg-1, con CF de 0.71±0.008 cm. De árboles localizados en lugares con poca pendiente se recolectaron frutos que tienden a ser redondos, a diferencia aquéllos procedentes de pendientes más pronunciadas, que perfilan forma alargada.
No sucede lo mismo con las semillas recolectadas de árboles situados en sitios pendientes <25 %, pues entre 25 y 40 % de pendiente, alcanzaron un peso superior (0.06 g), con coeficientes de forma de 0.71; mientras que, las semillas procedentes de árboles localizados en sitios con pendientes >40 % fueron más pequeñas, pero más abundantes (27 040 semillas kg-1); en contraste con las obtenidas en árboles ubicados en pendientes <25 % que fueron más grandes, con un total de 16 789 semillas kg-1 (Cuadro 4).
La altitud de los sitios en que crecen los árboles también fue un factor con efecto significativo (p≤0.01), pues en el intervalo altitudinal entre 1 178 y 1 278 m, el peso de los frutos fue de 0.36 g, con los menores CF (0.74); estos datos contrastan con los observados en altitudes ≤1 178 m, cuyo CF fue de 0.76; a la misma altitud, la producción de frutos fue de 4 441.
La semilla tuvo una respuesta similar con 0.04 g en altitudes >1 178 m; sin embargo, la producción de semillas fue mayor en altitudes ≤1 178 m (29 259.4) con coeficientes de forma de 0.72 cm en altitudes ≤1 278 m (Cuadro 4).
Las diferencias en la morfología de los frutos y semillas, en relación a las características propias del sitio (exposición, pendiente y altitud), se explican porque son componentes influyen en la radiación solar que incide sobre la planta, la temperatura y la humedad ambiental; condiciones que afectan la productividad primaria de la especie. En su estudio realizado en Quercus, Márquez et al. (2005) refieren que las diferencias de tamaños se deben a factores biológicos y físicos, además de que el tamaño de las semillas en una especie puede variar entre poblaciones o entre individuos, por razones genéticas o por la historia de vida de cada planta (Griffiths et al., 2009; García et al., 2015).
Conclusiones
La estructura poblacional de los árboles seleccionados como semilleros de Bursera simplex se caracteriza por tener una forma irregular, la cual es común en la selva baja caducifolia; de acuerdo con los índices evaluados, la estructura en que se desarrollan los árboles de B. simplex afecta el peso y la características morfología de los frutos y semillas.
En los sitios con exposición oeste se contabilizó el número mayor de individuos, además es donde existe mayor densidad de especies arbustivas, principalmente, con alturas ≤ 1.5 m, así como individuos de mayor tamaño como las burseras con 265 individuos ha-1. El árbol 21 es el mejor individuo semillero, ya que produce las semillas de peso más alto (0.08±0.13 g), con 16 179±5 036 semillas kg-1; mientras que los frutos de mayor peso estuvieron asociados al árbol cinco (0.42 g) y 2 456 frutos kg-1 con CF bajos y de forma elíptica. Las pendientes y altitudes de los sitios influyen en las características de los frutos y semillas, los árboles localizados en sitios con pendientes ≤ 40 % producen los frutos de mayor peso.

