











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los cambios de uso del suelo en los trópicos modifican los paisajes de bosques y selvas en masas forestales secundarias (FAO, 2010). La superficie de vegetación secundaria se ha extendido a nivel global. En México, se le conoce como “acahuales” y está aumentando como consecuencia de la degradación de más de 300 mil hectáreas de selvas al año; por lo que, últimamente, constituye la capa forestal dominante en las zonas tropicales (Semarnat, 2012; Mukul y Herbohn, 2016).
En general, los acahuales se consideran sistemas de flora que carecen de valor; en consecuencia, las políticas para su conservación denotan poco interés por parte de instancias gubernamentales (Sánchez-Sánchez et al., 2007). En algunos estudios se ha demostrado que dicha vegetación mantienen una variabilidad en su capacidad productiva acorde a su etapa sucesional, que puede ser multifuncional desde la perspectiva de una fuente alimentaria, medicinal y de suministros de productos forestales, además de servicios ambientales importantes, como protección al suelo y almacenamiento de carbono (Alayon-Gamboa et al., 2016), que los hace vitales para la economía rural, ya que en muchos casos logra reemplazar funciones básicas y presiones hacia los bosques primarios (Schulze et al., 2000; del Valle et al., 2011).
Los acahuales, por su extensión y nivel de incremento en biomasa, tienen potencial para mitigar el aumento en las concentraciones de CO2 atmosférico. Por ejemplo, a diferentes edades de abandono funcionan como grandes reservorios de carbono (C) que sobrepasan los 190 Mg C ha-1 (Johnson et al., 2000), pero la poca importancia que se le ha dado a su estudio, a nivel mundial, así como en México, ha resultado en escasa información y desconocimiento de su aporte y potencial en el ciclo global del carbono.
En México, los acahuales con potencial productivo mantienen una cobertura de 3 % (Semarnat, 2012), pero el poco conocimiento cuantitativo y cualitativo de sus atributos impiden determinar su viabilidad de conservación. Por esta razón, se estimaron datos básicos estructurales y de biomasa de acahuales a diferentes edades de abandono para dos componentes: árboles juveniles y árboles maduros de tres regiones ecogeográficas en Tabasco. El objetivo central del trabajo fue evidenciar el potencial de los acahuales con relación al servicio ambiental de captura de carbono; con la finalidad de contribuir a demostrar que esta vegetación puede considerarse en el esquema de pagos por servicios ambientales, que conlleve a mejorar la estrategia de planeación del recurso biomasa como método de conservación integral de la biodiversidad, para minimizar el deterioro del capital natural en el trópico húmedo.
Materiales y Métodos
Área de estudio y toma de datos
La investigación se realizó en la subregión de los Ríos de Tabasco, entre las coordenadas 17°16’00’’ y 18°12’00’’ de latitud norte; 90°56’00’’ y 91°52’00’’ de longitud oeste, en tres unidades ecogeográficas (UE) (Ortiz-Pérez et al., 2005): Laderas Septentrionales de la Sierra del Norte de Chiapas (LSSNCh) en Tenosique; Terrazas en Balancán (T); y Planicies Fluviales de Corrientes Alóctonas (PFCA) en Emiliano Zapata, las cuales presentan un arreglo geológico-estructural con un gradiente altitudinal de 250 m en la parte de la sierra y de 6 msnm hacia la costa del Golfo de México, y en su parte central de 50 msnm (Figura 1). Las precipitaciones en las laderas (3 000 mm) favorecen un clima cálido húmedo con lluvias todo el año (Af(m)); mientras que, en las zonas de altitud media y baja, la precipitación media anual es de 1 500 mm, con clima cálido húmedo y lluvias en verano (Am(fz)) INEGI (1986).
Figura 1 Unidades ecogeográficas (UE) y sitios de muestreo en la subregión Ríos; Laderas Septentrionales de la Sierra Norte de Chiapas (LSSNCh) en el municipio Tenosique, Terrazas (T) en el municipio Balancán y Planicies Fluviales de Corrientes Alóctonas (PFCA) en el municipio Emiliano Zapata, de Tabasco, México.
En cada UE se establecieron seis conglomerados de 1 600 m2, constituidos de cuatro parcelas de 10 × 40 m, con un arreglo de Y invertida (Conafor, 2011), los sitios se seleccionaron con base en la cartografía del estudio regional de la Umafor Ríos (Cámara-Cabrales et al., 2011). In-situ, se establecieron las edades de abandono de los acahuales, de acuerdo con el conocimiento de campo de los dueños de los predios. En las PFCA la edad de abandono fue de 100 años; en T de 15 y 20 años; en LSSNCh de 20 y 30 años. Asimismo, se consideraron dos muestras de selva en LSSNCh, como referencia de una vegetación madura.
Las variables para determinar la estructura fueron nombre común y científico, diámetro normal (DN), altura total y densidad especifica de la madera (Zanne et al., 2009). Con esta información se calcularon los valores de densidad (Ecuación 1), área basal (Ecuación 2) y biomasa (Ecuación 3). De la medición del diámetro normal (DN), se catalogaron dos componentes dentro de los acahuales; árboles maduros (Amad) a los DN ≥10 cm y como árboles juveniles (AJuv) a DN ≥ 2.5 cm y ≤ 9.9 cm; a dicha categorización se le aplicó la ecuación alométrica de biomasa propuesta por Chave et al. (2005) para selvas húmedas (1 500 a 3 500 mm), en conjunto con las ecuaciones alométricas específicas para cada especie disponible en Rojas-García et al. (2015).
Donde:
AB = Área basal por árbol (cm2)
D = Diámetro normal (cm)
Donde:
Y = Biomasa (kg árbol-1)
ln = Logaritmo natural
ρ = Densidad de cada especie (g cm-3)
D = Diámetro normal (cm)
h = Altura (m)
El análisis de varianza y la comparación de medias de Tukey-Kramer HSD se efectuaron con el paquete estadístico JMP 2008®. Las pruebas a posteriori se utilizaron para observar diferencias estadísticas entre áreas basales y biomasa para los tiempos de abandono de los acahuales como la principal fuente de variación (p≤ 0.05).
Resultados
Atributos estructurales y biomasa
Se observó, de manera general, que conforme el tiempo de abandono es mayor, el número de individuos totales disminuye. En las Planicies Fluviales de Corrientes Alóctonas (PFCA), la edad de 100 años registró menor densidad de árboles (406 Ind. ha-1) y 131 Ind. ha-1 en AJuv. La diferencia porcentual para la densidad de árboles entre juveniles y adultos se evidenció más en las PFCA, donde el número de Amad se mantuvo por arriba de 35 %, en comparación a los AJuv; en T la diferencia fue inferior a 2 %; mientras que, en las LSSNCh para los Amad la proporción fue de hasta 28 % (Figura 2).
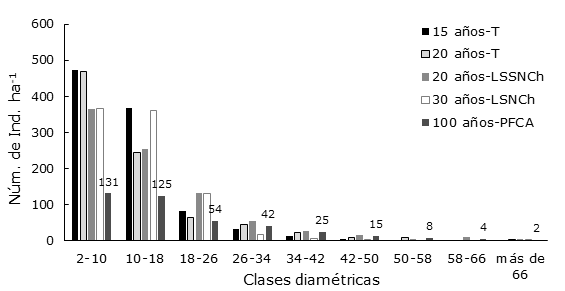
Figura 2 Número de individuos por hectárea (Núm. de Ind. ha-1) por clase diamétrica (cm) en los acahuales a diferentes edades, en cada una de las unidades ecogeográficas (UE) identificadas en la cuenca baja del Usumacinta, Región de los Ríos, Tabasco, México.
En la distribución diamétrica, la mayor concentración de Ind. ha-1 en cada tiempo de abandono se presentó tan solo en dos clases diamétricas: la categoría de 2 a 10 cm, conocida como árboles juveniles y la de 10 a 18 cm de árboles maduros. Ambas son las que están mejor representadas en todos los tratamientos, ya que la primera de las clases mantiene hasta 52 % del total Ind. ha-1 y en la segunda hasta 38 % del total. En las clases diamétricas subsecuentes, el número de Ind. ha-1 disminuye progresivamente conforme los diámetros normales aumentan, con un comportamiento de J invertida para todos los tiempos de abandono. En estos acahuales, la presencia de diámetros superiores a 50 cm es consistente para todos los tiempos de abandono; aunque, también se observa la falta de algunas clases diamétricas en ciertos tiempos. Los acahuales de 20 años en LSSNCh y de 100 años en PFCA presentaron todas las clases diamétricas, hasta un DN de 66 cm (Figura 2).
En las distribuciones de altura se distingue la existencia de tres estratos arbóreos; uno bajo de 2 a 7 m, uno medio de 7 a 12 m y uno alto que comprende de 12 a 17 m de altura. A la clase 2 a 7 m, le correspondió de 36 a 64 % del total de los individuos en cada tiempo de abandono (Figura 3).
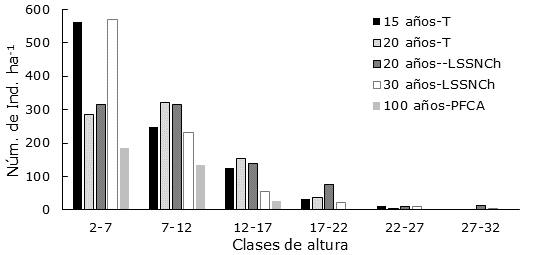
Figura 3 Número de individuos por hectárea (Núm. de Ind. ha-1) por clase de altura (m) en los acahuales a diferentes edades, en cada una de las unidades ecogeográficas (UE) identificadas en la cuenca baja del Usumacinta, Región de los Ríos, Tabasco, México.
El acahual de 30 años en LSSNCh registró el porcentaje más alto de individuos en la clase de altura de 2-7 m, la cual tuvo 64 % del total Ind. ha-1, para este tiempo de abandono. En los acahuales de 20 años y 30 en LSSNCh se observaron árboles emergentes de hasta 30 m de altura. La tendencia fue que a medida que la clase de altura aumenta, el número de individuos disminuye, y en algunas edades estas clases desaparecen. En las Planicies Fluviales de Corrientes Alóctonas (PFCA), la altura alcanza un máximo de 17 m. Al igual que los diámetros, el comportamiento de las alturas después de la clase de 7 a 12 m evidencia una J invertida, en el cual disminuye el número de individuos hacia las siguientes clases superiores en cada tiempo de abandono (Figura 3).
El área basal (AB) de los Amad fluctuó de 12.6 ± 2.10 m2 ha-1 a 25.84 ± 2.34 m2 ha-1. La mayor AB se presentó en el acahual de 20 años (25.84±2.34 m2 ha-1), localizado en la unidad ecogeográfica LSSNCh. El análisis de varianza (ANOVA) mostró diferencia estadística significativa entre estas AB en todas sus etapas sucesionales (p≤0.0016). La prueba de Tukey-Kramer HSD (TKHSD) indicó que solo la edad de 20 años en LSSNCh es significativamente diferente (p≤0.05), en relación con las edades de abandono en las otras UE. En la unidad ecogeográfica T, se observó un aumento sobre la edad sucesional; es decir, con mayores tiempos de abandono, existe un área basal más grande. En LSSNCh este comportamiento se expresó de forma contraria, debido a que su AB no se incrementó con un estado sucesional más avanzado.
Las AB en AJuv correspondieron al intervalo de 0.20 ± 0.06 a 1.07 ± 0.18 m2 ha-1. El análisis de varianza mostró diferencias estadísticas significativas entre las edades (p≤0.0008). La prueba a posteriori denota diferencia significativa solo entre los acahuales existentes en T de 15 y 20 años, con respecto al de 100 años perteneciente a PFCA. En las AB para árboles juveniles existe una tendencia a incrementarse para cada unidad ecogeográfica, porque conforme aumenta la edad de abandono, ocurre lo mismo con el área basal (Figura 4).
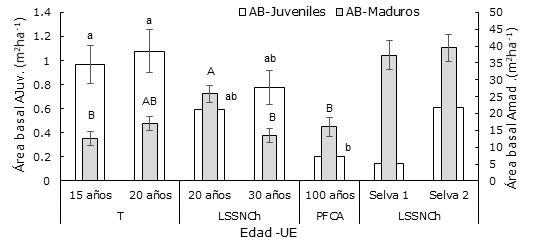
Las líneas verticales representan el error estándar. Las letras diferentes mayúsculas para Amad. y minúsculas para AJuv. indican diferencias significativas entre edades (α ≤ 0.05). Así mismo, las áreas basales de dos muestras de selva mediana.
Figura 4 Del lado izquierdo. - Área Basal (m2 ha-1), en el componente árboles Juveniles (AJuv) para los acahuales en diferentes edades en las unidades ecogeográficas (UE). Del lado derecho. - Área Basal (m2 ha-1), en el componente árboles maduros (Amad.).
La biomasa aérea en los tiempos de abandono con presencia para Amad tuvo registros de 150.90 ± 21.79 a 63.51 ± 24.57 Mg ha-1. La máxima concentración se determinó en la edad de abandono de 20 años, dentro de la unidad ecogeográfica LSSNCh. El ANOVA reveló diferencia significativa (p≤0.0045) entre los contenidos de biomasa aérea para estos tiempos de abandono. La prueba de Tukey-Kramer HSD (α≤0.05) mostró que el acahual de 20 años en LSSNCh es similar al contenido de biomasa con el acahual de 20 años en T y diferente a los otros tiempos de abandono en las unidades ecogeográficas. En T, el incremento de biomasa con respecto al tiempo de abandono aumentó; mientras que, en LSSNCh el comportamiento fue contrario (Figura 5).
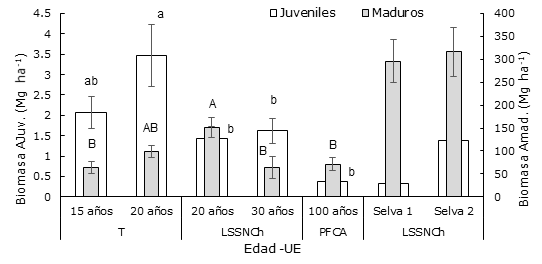
Las líneas verticales representan el error estándar. Las letras diferentes indican diferencias significativas entre edades (α ≤ 0.05), mayúsculas para Amad. y minúsculas para AJuv. Asimismo, la biomasa de dos muestras de selva mediana.
Figura 5 Del lado izquierdo. - Biomasa (Mg ha-1) en componente árboles Juveniles (AJuv) en acahuales a edades diferentes en las unidades ecogeográficas (UE). Del lado derecho. - Biomasa (Mg ha-1) en el componente árboles maduros (Amad).
En árboles juveniles (Figura 5) la biomasa registrada fue de 3.47 ± 0.76 Mg ha-1, como valor máximo y un mínimo de 0.375 ± 0.13 Mg ha-1. El ANOVA indicó diferencia estadística significativa entre los contenidos de biomasa, con respecto a la edad de abandono (p≤0.0004). La prueba a posteriori (α≤0.05) reveló que en T ambos acahuales son similares en sus contenidos de biomasa. El acahual de 20 años en T presentó diferencia significativa para las edades de 20 y 30 años en LSSNCh y para la edad de 100 años en PFCA. En T, el componente árboles juveniles tuvo un comportamiento de aumento, con respecto a un tiempo de mayor abandono y de forma inversa en LSSNCh.
Discusión
De manera general, a un menor tiempo de abandono le correspondió una densidad superior de individuos. Lo anterior sugiere que las especies que componen esas etapas sucesionales responden, positivamente, a una alta disposición de recursos que favorecen en cierta manera el crecimiento y supervivencia, al no existir una gran competencia por nutrimentos y luz, cuya característica primordial es mantener un gran número de individuos en clases diamétricas pequeñas (Smith et al., 1997; Ajbilou et al., 2003). Asimismo, Morales-Salazar et al. (2012) y Puc (2014) señalan que, para bosques tropicales secundarios la densidad arbórea tiene una tendencia al aumento al incrementarse el tiempo de abandono, que difiere de lo registrado en este trabajo; ya que la densidad de individuos en los sitios con mayores tiempos de abandono disminuyó progresivamente, esto se explica por la incidencia de algún fenómeno climatológico o por un aprovechamiento selectivo, aunque no se tiene registro de ello.
A lo largo de la cronosecuencia, la densidad de AJuv a un tiempo superior de abandono, se ajustó a lo esperado de manera natural; puesto que conforme avanza el grado de sucesión, por cualidades intrínsecas de historia de vida de las especies, esta disminuye (Chazdon et al., 2007), pero se ha determinado que dicha densidad baja, cuando la apertura del dosel disminuye, lo cual incide en la supervivencia y crecimiento de los árboles de algunos grupos (Muñiz-Castro, 2008). Pero si existiera otro comportamiento que altere dicha generalidad, probablemente, se deba a una perturbación episódica (Ajbilou et al., 2003) que incrementa la regeneración, y, en consecuencia, el número de individuos juveniles proliferen; por ejemplo, el acahual de 30 años y selva 2, donde se observó que el dosel se abrió por la tala selectiva de especies maderables, lo cual hizo que, probablemente, aparecieran especies dependientes de la apertura de claros en el dosel.
En PFCA (100 años) la poca densidad de AJuv y Amad está influenciada por características propias del sitio, más que por la edad de abandono, ya que es una zona de inundación temporal lo que puede incidir en la poca densidad registrada; Cortés-Castelán e Islebe (2005) sugieren que la densidad arbórea tiende a ser menor en estas zonas que en relieves altos. Asimismo, la anegación no permite la regeneración, principalmente, por semillas al descomponerse estas con el agua, hecho que incide en una baja tasa de germinación para el establecimiento de plántulas que enriquezcan las densidades de AJuv y Amad. También, se ha observado que el efecto de la inundación causa reducción en la disponibilidad de oxígeno en las raíces, con la consecuente senescencia y mortalidad (Moreno y Fischer, 2014), y con ello se favorece una baja densidad.
El patrón de distribución de los individuos por clase diamétrica tiene una forma de J invertida, independientemente, de la edad de abandono, con una tendencia hacia la disminución del número de individuos en las categorías diamétricas mayores (Figura 2), en concordancia con lo citado por Morales-Salazar et al. (2012) para bosques secundarios menores y con más de 30 años, producto del abandono de potreros en Costa Rica. Al igual que Carreón-Santos y Valdez-Hernández (2014) y Puc (2014), se observó que gran parte de los individuos se agrupan en dos categorías diamétricas inferiores a los 20 cm de DN, la primera de 2-10 cm (AJuv) y de 10 a 18 cm(Amad); por lo tanto, dichos acahuales están en fase de regeneración óptima (Guariguata, 1998; Ajbilou et al., 2003) que aseguraran la persistencia del bosque y el equilibrio del mismo, a través del proceso de sucesión (Lamprecht, 1990; Higuchi et al., 2008). También se evidenció que en estados sucesionales de edades menores, sin superar los 30 años, existen más individuos por hectárea, que a edades superiores.
La distribución diamétrica, afianzada en dos clases menores a 20 cm de DN, independiente del tiempo de abandono en las UE es posible que responda a situaciones de la historia de uso de la tierra y a los impactos que modifican las distribuciones diamétrica, ya que en los acahuales de T, por ejemplo, se han registrado incendios que favorecen el repoblado de árboles jóvenes; mientras que en LSSNCh (30 años), se registró un aprovechamiento que, eventualmente, modifica el dosel y repercute en la presencia de clases diamétricas pequeñas. Los valores extremos que forman clases diamétricas mayores, que no corresponderían a edades tempranas de abandono (si se supone un incremento medio anual de 1 cm), después de la roza-tumba y quema, se explican por la historia del uso de la tierra, ya que es una práctica común en estas zonas mantener árboles de sombra o de algún valor comercial, por lo que se registraron ejemplares de grandes dimensiones.
La distribución de los individuos por su altura para las distintas edades muestran que tres estratos son los que sobresalen, un comportamiento similar es citado por Díaz et al. (2002) y Carreón-Santos y Valdez-Hernández, (2014). Los árboles tanto de acahuales, como de selvas estuvieron dispuestos en clases de altura de estratos bajos, menores a los 10 m de altura, los cuales representan 80 % sobre las demás clases. Carreón-Santos y Valdez-Hernández (2014), también señalan que la característica de J invertida advierte que se trata de masas forestales jóvenes, con buena repoblación. Ajbilou et al. (2003) documentan que una mayor densidad de individuos jóvenes en estratos inferiores es una característica que revela un nivel de perturbación más alto. Esta condicionante de disturbio se observa en la región de LSSNCh y T para el acahual de 30 años y 15 años, donde aparentemente existe un aprovechamiento relativo de los recursos, así como incendios recientes en T.
En cuanto a las alturas en las UE, se observa que en PFCA no hay más de tres estratos (17 m), comportamiento que corresponde a la presencia de árboles de poca talla que están sujetos a condiciones altas de anegación (Cortés-Castelán y Islebe, 2005); como ocurre en áreas con selvas inundables en Tabasco, donde el dosel no supera los 20 m de altura (Rzedowski, 1979).
Las áreas basales (AB) para Amad y AJuv variaron, respecto al tiempo de abandono. Solo en T, para Amad con un tiempo de abandono mayor, le correspondió un AB superior. En el componente AJuv, en general, en cada una de las unidades ecogeográficas hay una tendencia al aumento, en relación a un mayor tiempo de abandono.
Las AB solo en Amad están por encima de lo considerado en la legislación vigente de México para vegetación secundaria, de 4 m2 ha-1, dicha subestimación puede promover la no conservación o restauración de estos ecosistemas forestales, en lo particular cada tiempo de abandono presenta un AB, cercana a la de bosques primarios, como lo indica Plonczak, (2005) para un bosque seco tropical en Venezuela (DN ≥ 10 cm), cuya AB estuvo alrededor de los 21.4 m2 ha-1; y al de Cuello (2002) para una selva nublada en Venezuela, con 26.6 m2 ha-1 para su valor más bajo de (DN ≥ 2.5 cm). El AB de los acahuales con una fracción de la edad de las selvas medianas en este estudio, podrían en poco tiempo alcanzar un AB similar a los valores estimados para estas selvas, así como a los valores existentes en áreas de selvas con estatus de protección dentro de la unidad ecogeográfica LSSNCh, de 40 m2 ha-1. Las AB estimadas para los acahuales son superiores a las registradas por Puc (2014), para edades similares.
En el presente estudio, las AB no aumentaron al pasar de un tiempo de abandono al subsecuente, debido a factores como el efecto que causa la corta selectiva para uso doméstico que se observa principalmente en LSSNCh, para el acahual de 30 años, donde el AB de AJuv es mayor y en Amad es menor, que para uno de 20 años en la misma UE, lo mismo se registra para selva 2, en el componente AJuv, donde existe una corta selectiva para uso local o doméstico; a diferencia de selva 1, cuya AB en el componente AJuv tiene valores bajos, unidad en la que aparentemente no hay acciones importantes de tala selectiva.
La acumulación de biomasa, en general, fue variable y se relacionó con las edades de abandono (del Valle et al., 2011). Estos valores de biomasa correspondieron con los 190 Mg C ha-1, para masas forestales secundarias menores a 100 años (Johnson et al., 2000; Puc, 2014). Read y Lawrence, (2003) y Puc, (2014) indican que acahuales de 5 a 80 años aportan entre 20 Mg ha-1 y 39.75 Mg ha-1, con diámetros menores a 7.5 cm (juveniles); mientras que, de 179 ± 7 Mg ha-1, en acahuales con DN ≥ 7.5 cm, conceptualizados como árboles adultos para el sur de Quintana Roo. Por tanto, la biomasa en los dos componentes estudiados está en el intervalo esperado para los contenidos de biomasa a distintas edades de abandono.
La cantidad de biomasa en los acahuales para la edad de abandono de 15 años fue superior a las registradas por Urquiza-Haas et al. (2007) para una edad similar en bosques estacionalmente secos de Yucatán y subhúmedos de Quintana Roo; aunque para rodales maduros de 100 años en PFCA (70.9 Mg ha-1) es bajo, en comparación a los de estos autores (191 Mg ha-1).
En general, se observa que la biomasa para tiempos de abandonos menores a los 30 años, tiene un potencial de acumulación de 100 a 150 Mg ha-1, que supera los 100 Mg ha-1 considerados por Brown y Lugo, (1990) y Silver et al. (2000) para edades menores de 20 años; asimismo, si se mantienen. con el tiempo podrían alcanzar valores similares de biomasa a los de las selvas en la mitad del tiempo, ya que estas tuvieron hasta 300 Mg ha-1 de biomasa, la cual está dentro del intervalo general señalado para distintos tipos de bosques tropicales primarios, de 200 a 400 Mg ha-1 (Sarmiento et al.. 2005; Yepes et al., 2010).
La variabilidad en los contenidos de biomasa a edades similares que se han documentado responden a factores propios de cada ecosistema, lo cual permite expresar como máximos o mínimos dichos contenidos de biomasa alrededor del mundo (Ngo et al., 2013; Rutishauser et al., 2013; Puc, 2014). Aunque el aporte de biomasa entre Amad y AJuv presentó una diferencia de 95 %, son similares a los de Read y Lawrence, (2003), para el sur de México. Dupuy et al. (2012) citan que esa proporcionalidad mayor del componente Amad, se debe a que, en principio, AJuv tienen diámetros y alturas pequeñas a pesar de su abundancia, lo cual, por razones de competencia, mantiene esta desproporcionalidad en todas las etapas sucesionales.
Conclusiones
Los acahuales estudiados presentan diferencias significativas en sus atributos estructurales, dependientes de la edad de abandono, así como de factores propios de cada UE. Aunque, la cronosecuencia en las unidades ecogeográficas no sigue un patrón de incrementos generalizados conforme avanza la edad, en algunas parecen ajustarse a lo señalado por diversos autores con respecto a los incrementos estructurales y de biomasa. Que no se ajuste, se debe a la relación de la vegetación con fenómenos naturales y antrópicos que condicionan las características intrínsecas de vida de los acahuales, que propician la variabilidad estructural, lo cual incide en la acumulación de biomasa. Los acahuales, al ser una tendencia mundial por los cambios de uso de suelo, pueden mantener un potencial de secuestro de C por su acelerada acumulación de biomasa, signo positivo para la mitigación del cambio climático global, derivado de los gases de efecto invernadero.
Los acahuales con menor tiempo de abandono tienen un potencial de acumulación de biomasa, en gran medida también son los más vulnerables, ya que la poca importancia que tienen para los dueños de la tierra da pauta para ser intervenidos continuamente. Los acahuales aquí estudiados, con menos de la mitad de edad de las selvas, presentan atributos estructurales similares a ellas con una considerable acumulación de carbono, si se manejaran de manera adecuada podrían ser significativamente benéficos para ayudar a mitigar el incremento de CO2 atmosférico.

