











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
De la gran diversidad de tipos de vegetación que existe en México, los bosques templados ocupan 12 % del territorio nacional (Semarnat, 2009) cuya importancia radica, en términos económicos, en los bienes y servicios que suministran (Díaz, 1988).
Los bosques mexicanos se enfrentan a incendios, tala excesiva (Salinas et al., 2010), plagas y enfermedades. Sobresalen los barrenadores de conos y semillas (Cibrián et al., 1998), los defoliadores (Castro, 1981), como los géneros Neodiprion y Zadiprion (González et al., 2014); los muérdagos, que han causado pérdidas anuales por más de dos millones de metros cúbicos de madera (Vázquez et al., 2006) y los descortezadores, principalmente del género Dendroctonus, del cual existen 13 especies (Armendáriz y Zúñiga, 2017); por su agresividad destacan Dendroctonus frontalis Zimmermann, Dendroctonus mexicanus Hopkins, Dendroctonus adjuntus Blandford y Dendroctonus rhizophagus Thomas & Bright, que son fundamentales en la dinámica y renovación natural de los bosques (Salinas et al., 2010).
De 1990 a 2014 se registraron en México 69 mil ha afectadas por barrenadores; 23 mil ha por defoliadores; 383 mil ha por muérdagos y 474 mil ha por descortezadores, de las cuales solo 60 %, en promedio, recibió alguna práctica de control o manejo (Semarnat, 2015); lo que puede explicarse por la incertidumbre sobre el potencial de los bosques afectados por tales agentes.
En ese contexto, los modelos de distribución potencial de especies en conjunto con los sistemas de información geográfica constituyen una herramienta que ayuda a sintetizar y entender la relación entre los taxa y las variables ambientales (Pliscoff y Fuentes, 2011). El software MaxEnt (Phillips et al., 2006) ha sido utilizado para este fin en las últimas décadas y busca determinar la máxima entropía en la que todos los sitios tienen la misma probabilidad de ocurrir (Palma y Delgadillo, 2014).
La modelación de distribución de especies se ha realizado a menudo con variables climáticas (Contreras et al., 2010; Salinas et al., 2010; Palma y Delgadillo, 2014); sin embargo, para plagas forestales, la dasometría también debe ser considerada, puesto que en ocasiones la presencia de una plaga no es únicamente dependiente del clima, sino también de la estructura de los rodales (Raffa et al., 2008; Cuellar et al., 2013).
Con la intensificación del cambio climático, se prevé que los insectos descortezadores tengan condiciones favorables para su fisiología (Raffa et al., 2008) y por ende en su desplazamiento; por lo tanto, es necesario generar información para conocer su distribución potencial y su relación con variables climáticas, además de sus preferencias en el bosque.
Con base en todo lo anterior, se planteó el objetivo de modelar la distribución potencial de barrenadores de conos y semillas, defoliadores, descortezadores y muérdagos en bosques de coníferas en México, mediante el uso de variables bioclimáticas, dasométricas y el algoritmo MaxEnt.
Dado que, generalmente, las brigadas de campo del Inventario Nacional Forestal y de Suelos no están capacitadas para la identificación correcta de las plagas forestales, y ante la escasez de estudios a nivel especie que pudiesen aportar información sobre la distribución nacional de esos agentes, en este trabajo se ha modelado a nivel de tipo de plaga. Con ello se amplía el área de oportunidad para realizar estudios posteriores y más específicos. Así, se espera contribuir a la detección temprana de estos organismos y evitar su proliferación, la cual es información básica para la planificación, monitoreo, manejo y conservación de los bosques del país.
Materiales y Métodos
El área de estudio
Como ámbitos de estudio, se seleccionaron el bosque de pino, el bosque de pino-encino y el bosque de oyamel (INEGI, 2013) con un buffer de 1 km, que se ubican principalmente, en la Sierra Madre Occidental y en la Sierra Madre Oriental, entre otros relieves montañosos En estos tipos de vegetación, el clima es templado subhúmedo con temperaturas promedio de 10 a 22 °C; la precipitación varía de 600 a 1 000 mm anuales (Conagua, 2012). El intervalo altitudinal parte de los 400 a 4 000 m.
Registros de plagas forestales
Los datos de plagas forestales fueron proporcionados por la Comisión Nacional Forestal (Inventario Nacional Forestal y de Suelos 2009-2014, INFyS). A dichos datos se les depuró para verificar que estuvieran ubicados dentro del polígono de área de estudio, que corresponde a todos los bosques de coníferas del país; lo cual se realizó mediante el programa QGIS (QGIS, 2016) y se eliminaron aquéllos que no cumplieron con esa condición. Se obtuvieron 4 004 registros únicos (coordenadas geográficas) de cuatro tipos de plaga: 316, 546, 1 527 y 1 615, para barrenadores (B), defoliadores (d), descortezadores (D) y muérdagos (M), respectivamente; en la información del INFyS no se precisan las especies de plagas observadas en campo, por lo que se optó por organizar los registros de presencia en los cuatro grupos mencionados anteriormente. Se asume, además, que la agrupación de los muérdagos se refiere a los enanos, puesto que la información de referencia se reduce solo a observaciones de dichos organismos.
Variables y modelos de distribución
Se utilizaron las variables bioclimáticas propuestas por Hijmans et al. (2005) que son variantes de la precipitación y temperatura, y tienen una resolución aproximada de 1 km2 (http://www.worldclim.org). Adicionalmente, se generaron variables dasométricas (Aba: área basal ha-1; Arb: número de árboles ha-1; Dnp: diámetro normal promedio ha-1; Nsp: número de especies), con datos del INFyS, mismos que se tabularon y se extrapolaron a hectárea para su posterior interpolación al área de estudio, mediante el método de Interpolación de Distancia Inversa (IDW), porque se ajustó mejor a los datos reales.
De las variables bioclimáticas y dasométricas se eliminaron aquéllas con correlación mayor a 0.6 (Contreras et al., 2010), para lo que se usó la función corselect del paquete fuzzySim (Barbosa, 2015) del programa R (R Core Team, 2017), con el fin de minimizar la colinealidad en los modelos resultantes (Contreras et al., 2010). Las variables bioclimáticas seleccionadas para la modelación fueron: temperatura promedio anual (°C), B01; isotermalidad de la temperatura, B03; estacionalidad de la temperatura, B04; temperatura mínima promedio del periodo más frío (° C), B06; temperatura promedio del cuarto anual más húmedo (°C), B08; temperatura promedio del cuarto anual más cálido (° C), B10; precipitación anual (mm), B12; precipitación del periodo más seco (mm), B14; estacionalidad de la precipitación (mm), B15; precipitación del cuarto anual más cálido (mm), B18; precipitación del cuarto anual más frío (mm), B19.
Los registros y las variables seleccionadas de cada tipo de plaga se utilizaron para generar modelos de distribución potencial en el programa R (R Core Team, 2017) con ayuda de MaxEnt versión 3.4.1 (Phillips et al., 2006) y el paquete dismo (Hijmans et al., 2016) de R (R Core Team, 2017). Para generar los modelos de distribución se dividieron los datos de presencia por tipo de plaga en cinco grupos aleatorios(k-fold), con los que se hace el mismo k número de iteraciones, en cada una de las cuales se toma un grupo definido para verificación y el resto de ellos como entrenamiento del modelo; el proceso se repite de acuerdo al número de k-folds (Hijmans et al., 2016).
El estadístico AUC (Área Bajo la Curva, por sus siglas en inglés) se usó para conocer la precisión de los modelos (Elith, 2000) en la predicción de la distribución potencial de los tipos de plaga. Los modelos de distribución se proyectaron al espacio geográfico y se definió la probabilidad de presencia por arriba del umbral mínimo de presencia conocida sugerido por Pearson et al. (2007).
Perfil bioclimático de plagas forestales
Con MaxEnt se realizó una prueba de Jackknife que ayudó a identificar las variables más importantes en la prueba de los modelos de distribución potencial; de estas variables se extrajeron los valores máximos y mínimos correspondientes a los registros de presencia para generar el perfil bioclimático de cada tipo de plaga.
Resultados y Discusión
Ajuste de modelos de distribución potencial
Los modelos generados se consideran adecuados para predecir la distribución de plagas forestales, por sus valores de AUC mayores a 0.85, que de acuerdo con Elith (2000), cuando estos son superiores a 0.75 son útiles para predecir la distribución potencial. El AUC más alto fue para defoliadores y el menor para descortezadores, mientras que el umbral mínimo de presencia conocida (Pearson et al., 2007) fue de 0.05 en promedio para los tipos de plaga estudiados (Figura 1).
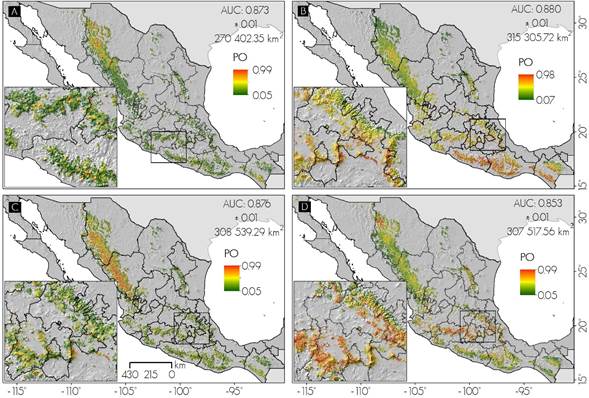
PO = Probabilidad de ocurrencia; AUC = Área Bajo la Curva
A =Barrenadores; B = Defoliadores; C = Muérdagos; D = Descortezadores
Figura 1 Distribución potencial de las plagas que afectan los bosques de coníferas de México.
En cuanto al ajuste, el valor de AUC (0.97) calculado por Mendoza et al. (2011) para la distribución de Dendroctonus rhizophagus en la Sierra Madre Occidental es superior al de esta investigación (0.85), lo que se debe, principalmente, a la especificidad de ambos estudios; en la presente investigación se trabajó a nivel género, mientras que otras lo hacen con especies, y en la medida en que esté localizada, la predicción de los modelos es mejor (Allouche et al., 2006). Asimismo, Salinas et al. (2010) predijeron la distribución de 11 especies de descortezadores en México, con apoyo en el algoritmo BIOCLIM; sin embargo, no muestran el ajuste de dichas predicciones que permita formular una discusión sobre el correspondiente al modelo aquí presentado para descortezadores, aunque no se documenta información acerca de una especie de descortezador en particular.
Distribución potencial de plagas forestales
Barrenadores. Los estados del país con mayor superficie de distribución potencial para barrenadores (probabilidad ≥ 0.85) fueron Chihuahua (23 %), Durango (16.9 %) y Oaxaca (10.2 %) de 270 402.35 km2) (Figura 1), mientras que en la sierra de San Pedro Mártir (Baja California), donde Díaz (1988) ha descrito barrenadores del género Cydia sobre Pinus jeffreyi Balf. y P. ponderosa Douglas ex C. Lawson, el modelo predice probabilidad de presencia de hasta 0.88. Galindo et al. (2011) hicieron el primer registro de la presencia del género Megastimus, causante de importantes daños a semillas de Abies religiosa (Kunth) Schltdl. et Cham. y A. hickeli Flous & Gaussen en el Cofre de Perote (Veracruz), donde se predice la probabilidad de presencia de 0.51. Lo anterior sugiere que en los sitios señalados por el modelo podrían existir barrenadores, y el hecho de que no hayan sido consignados o descritos solo indica la deficiencia en los estudios de campo. Se observan, también, en el Eje Neovolcánico algunas zonas de distribución discontinua, al igual que en Chiapas, Coahuila y Nuevo León (Figura 1A).
Defoliadores. MaxEnt estimó que en México existen 315 305.72 km2 (Figura 1B) que reúnen condiciones climáticas y dasométricas favorables para su presencia. La mayor probabilidad ocurre en los bosques del centro y sur del país (Figura 1B); con la superficie más grande en Chihuahua (66 367.47 km2), estado en el que Castro (1981) registró defoliadores del género Neodiprion en bosques de Pinus arizonica Engelm. en el municipio Bocoyna, Chihuahua; sitio en el cual, el modelo predijo probabilidades altas (> 0.90, Figura 1B). Otras entidades con amplia zona favorable para el desarrollo de defoliadores son Durango (51 655.29 km2), Oaxaca (32 351.08 km2), así como Chiapas, Guerrero y Michoacán con cerca de 19 801.3 km2, en promedio, para cada uno (Figura 1B).
Descortezadores. López et al. (2015) los identificaron en bosques de Abies religiosa en la Reserva de la Biósfera Mariposa Monarca; al igual que Rodríguez et al. (2013), quienes citan a Dendroctonus adjuntus en el ejido Los Pescados, Veracruz, localidades en las que el modelo pronosticó 0.68 de probabilidad de ocurrencia, en promedio, para ambas; aunque en áreas aledañas a la última predijo hasta 0.99. Chihuahua y Durango son los estados con el área potencial más amplia para descortezadores del género Dendroctonus (20.96 y 16.56 % del total nacional), principalmente, por tratarse de los estados con la mayor extensión de bosques, y que poseen la diversidad más alta de especies de Dendroctonus (Salinas et al., 2010). Sin embargo, en el Eje Volcánico Transmexicano el problema se incrementa (Figura 1D), lo que podría deberse a que en ese sistema montañoso se concentra un elevado porcentaje de Pinus colonizados por descortezadores (Salinas et al., 2010).
Muérdagos. El mayor número de registros se observó en Durango y Chihuahua (601 y 452), donde se verifica la extensión más grande de su distribución potencial (65 986.49 y 51 617.97 km2); en contraste, Aguascalientes y Baja California Sur albergan el área potencial más baja, con 27.7 y 161.01 km2 (Figura 2C), debido principalmente a lo limitado que son los bosques de coníferas en esas entidades. Se predijeron 308 539.29 km2 con características para la proliferación de alguna especie de muérdago. Estos organismos han sido consignados por Ramírez y Porcayo (2010) en el Parque Nacional Nevado de Toluca sobre Pinus hartwegii Lindl. y P. montezumae Lamb., en cuya zona se calculan probabilidades máximas de 0.60 (Figura 1C).
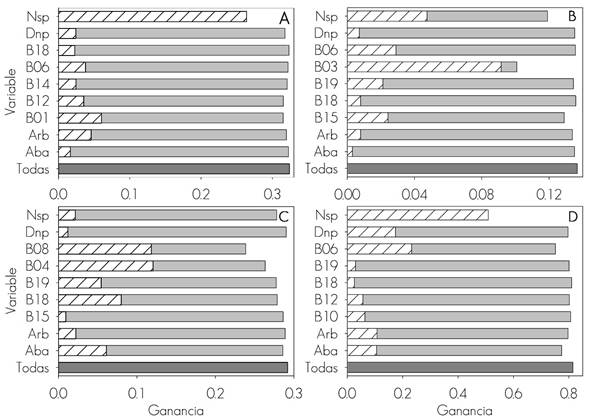
B01= Temperatura promedio anual (°C); B03 = Isotermalidad de la temperatura; B04= Estacionalidad de la temperatura; B06 = Temperatura mínima promedio del periodo más frío (°C); B08 = Temperatura promedio del cuarto anual más húmedo (°C); B10 = Temperatura promedio del cuarto anual más cálido (°C); B12 = Precipitación anual (mm); B14 = Precipitación del periodo más seco (mm); B15 = Estacionalidad de la precipitación (mm); B18 = Precipitación del cuarto anual más cálido (mm); B19 = Precipitación del cuarto anual más frío (mm); Aba = Área basal ha-1; Arb = Número de árboles ha-1; Dnp = Diámetro normal promedio ha-1; Nsp = Número de especies.
A = Barrenadores; B = Defoliadores; C = Descortezadores; D = Muérdagos
Figura 2 Importancia de las variables (Prueba de Jackknife) en modelos de distribución potencial de las plagas que afectan los bosques de coníferas de México.
En general, el área potencial para cada tipo de plaga fue superior a 270 mil km2, la menor de las cuales corresponde a los barrenadores y lo contrario para defoliadores (Figura 1). Durango reúne la superficie boscosa más extensa bajo condiciones óptimas para defoliadores, descortezadores y muérdagos, seguido por Chihuahua, mientras que para otros estados como Nuevo León, Coahuila y Nayarit la mayor susceptibilidad se verifica para descortezadores, y para barrenadores en los bosques de la Ciudad de México (Figura 2).
Ante el alto riesgo de que sus bosques sean atacados, en Oaxaca, Guerrero y Chiapas (para defoliadores); el Estado de México, Michoacán y Nuevo León (para descortezadores); Chihuahua y Durango (para muérdagos y barrenadores) se debería centrar la atención y el diseño de sistemas de monitoreo para la detección oportuna de brotes de alguna plaga (Figura 1).
Variables ambientales de importancia
Las variables ambientales que no presentaron correlación superior a 0.6 fueron cinco bioclimáticas (variables para cada TP) y cuatro dasométricas (Figura 3). De acuerdo con los resultados de la prueba de Jackknife, las que disminuyen la ganancia del modelo cuando se excluyen (barra gris claro de la Figura 2) son igual de importantes que las que aportan la mayor ganancia, cuando se modelan individualmente (barra blanca de la Figura 2); en ese sentido, el parámetro más relevante en la predicción de la distribución de barrenadores, defoliadores y muérdagos fue el número de especies (Nsp), mientras que para descortezadores, el área basal (Aba) (Figura 2D).
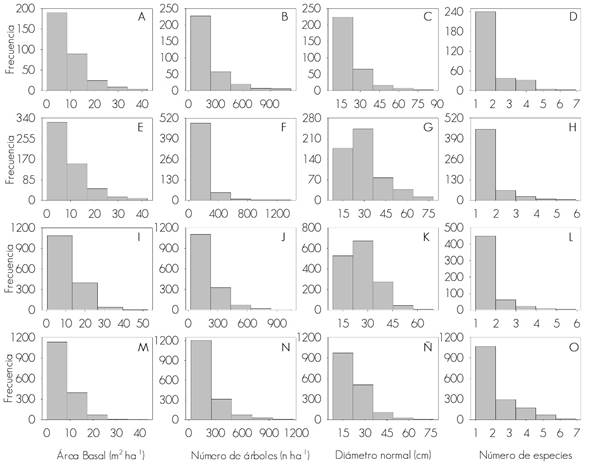
A – D = Barrenadores; E – H = Defoliadores; I – L = Descortezadores; M – O = Muérdagos.
Figura 3 Distribución de registros de presencia de plagas; con respecto a variables dasométricas.
De las variables climáticas, la isotermalidad (B03) fue más relevante para los defoliadores, la temperatura promedio anual (B01) para barrenadores y para muérdagos la temperatura mínima promedio del periodo más frío (B06) del año (Figura 2). En general, las que se asocian con la temperatura son las más significativas para los cuatro tipos de plaga (Figura 2), lo que quizás se deba a que condiciona fenómenos como la evapotranspiración o la severidad de las sequías, que mantienen al árbol en estrés y lo hacen vulnerable al ataque de los insectos (Raffa et al., 2008; Del Val y Sáenz, 2017).
Específicamente para descortezadores, las temperaturas bajas (B08: temperatura promedio del cuarto anual más húmedo del año (Figura 2) suponen un elemento importante en su distribución, al igual que otros efectos directos sobre los insectos, al propiciar la mortalidad de adultos y larvas, así como la alteración del tiempo requerido para completar una generación (Raffa et al., 2008); lo anterior repercute en su dinámica poblacional (Bentz et al., 2010). Sin embargo, por el cambio climático y el aumento de la temperatura, la relación limitante hacia los insectos podría decrecer paulatinamente y ocasionar una explosión poblacional (Hernández et al., 2017).
En general, el panorama para los bosques templados de México es poco alentador en el contexto de plagas. Por una parte, los insectos descortezadores son positivamente influenciados por las temperaturas ascendentes (Del Val y Sáenz, 2017), lo que da lugar a un incremento en el número de generaciones por año (Bentz et al., 2010); con ello, se multiplican sus posibilidades de adaptarse a las nuevas condiciones ambientales. Los árboles, por el contrario, no tienen la misma capacidad para ajustarse a los cambios que supone la presencia masiva de insectos, ni a los de su entorno.
Perfil ecológico-climático de plagas forestales
El porcentaje de contribución de las variables climáticas y dasométricas en los modelos de distribución no fue constante para los cuatro tipos de plaga. Para barrenadores y muérdagos, las variables dasométricas aportaron al modelo más de 50 %, mientras que las climáticas contribuyeron arriba de 70 % a la construcción de los modelos para descortezadores y defoliadores De acuerdo con la prueba de Jackknife, las variables más importantes (Figura 3) fueron también las que tuvieron mayor porcentaje de contribución en la construcción de los modelos (Cuadro 1).
Cuadro 1 Perfil ecológico con mínimos, máximos y porcentajes de contribución de variables bioclimáticas y dasométricas usadas en modelos de distribución de cuatro tipos de plagas forestales en México.
| Variable | Barrenadores | Defoliadores | Descortezadores | Muérdagos |
|---|---|---|---|---|
| B01(°C) | 9.5-25.0 (14.3) | - | - | - |
| B03 (°C) | - | 0.4-0.8 (58.2)† | - | - |
| B04 (Sd) (°C) | - | - | 5.1-63.2 (12.4) | - |
| B06 (°C) | 0.0-16.2 (0.5) | 0.0-17.5 (1.5) | - | 0.0-16.8 (34.5) |
| B08 (°C) | - | - | 3.3-26.2 (42.4)† | - |
| B10 (°C) | - | - | - | 7.9-27.9 (1.1) |
| B12 (mm) | 371.0-2595.0 (5.8) | - | - | 19.1-311.3 (1.3) |
| B14 (mm) | 1.0-54.0 (1.2) | - | - | - |
| B15 (mm) | - | 50.0-114.0 (10.1) | 48.0-118.0 (3.4) | - |
| B18 (mm) | 94.0-864.0 (0.3) | 51.0-908.0 (0.3) | 74.0-875.0 (9.5) | 50.0-967.0 (0.1) |
| B19 (mm) | - | 14.0-278.0 (1.8) | 17.0-370.0 (7.3) | 13.0-370.0 (2.0) |
| Aba (m2 ha-1) | 0.2-42.2 (1.2) | 0.1-42.2 (1.1) | 0.4-65.6 (15.0) | 0.1-43.3 (7.7) |
| Arb (n ha-1) | 25.0-1134.2 (4.0) | 25.0-1389.5 (1.8) | 25.0-1040.0 (2.1) | 25.0-1170.0 (3.8) |
| Dnp (cm) | 8.8-86.9 (1.6) | 8.0-78.2 (0.6) | 9.2-69.4 (0.3) | 8.1-76.7 (1.3) |
| Nsp (n) | 1.0-6.9 (66.7)† | 1.0-5.9 (24.6) | 1.0-7.9 (7.4) | 1.0-6.9 (46.8)† |
B01 = Temperatura promedio anual; B03 = Isotermalidad; B04 = Estacionalidad de la temperatura; B06 = Temperatura mínima promedio del periodo más frío; B08 = Temperatura promedio del cuarto anual más húmedo; B10 = Temperatura promedio del cuarto anual más cálido; B12 = Precipitación anual; B14 = Precipitación del periodo más seco; B15 = Estacionalidad de la precipitación; B18 = Precipitación del cuarto anual más cálido; B19 = Precipitación del cuarto anual más frío; Aba = Área basal por hectárea; Arb = Número de árboles por hectárea; Dnp = Diámetro normal promedio por hectárea; Nsp = Número de especies; † = Variable más importante de acuerdo a la prueba de Jackknife; Sd = Desviación estándar.
Los porcentajes de contribución están entre paréntesis.
Gan (2004) indica que el riesgo de infestación de Dendroctonus frontalis aumenta cuando las temperaturas oscilan de 8.79 a 14.87 °C; en la presente investigación el intervalo de temperatura promedio es más amplio para descortezadores (Cuadro 1); esto se debe, principalmente, a que los bosques mexicanos están a menor latitud que la región mencionada por el autor, y a que no se trabajó a nivel de especie.
Mendoza et al. (2011) aseveran que la temperatura media contribuyó con 30.1 % y la precipitación con 10.5 % en la distribución de Dendroctonus rizophagus en la Sierra Madre Occidental; en este trabajo se tiene el mismo comportamiento (54.9 % para temperaturas y 20.26 % para precipitación (Cuadro 1), lo que indica que la distribución de descortezadores se rige, en gran medida, por la temperatura (Figura 3D). Sin embargo, no solo la distribución de los insectos es afectada por la temperatura, sino también su explosión demográfica (Raffa et al., 2008), mortalidad (Bentz et al., 2010) y fisiología (Hernández et al., 2017).
Cuellar et al. (2012) señalan que, en el noreste de México, la precipitación y la temperatura (media y máxima) mensual no determinan la abundancia de Dendroctonus mexicanus, lo que a su vez demostraría que en algunas regiones, la distribución y abundancia de descortezadores no están condicionadas a cambios mensuales en el clima, pero sí a variaciones estacionales o incluso anuales (Mendoza et al., 2011).
En México, Queijeiro y Cano (2015) observaron que la incidencia de muérdagos (Arceuthobium vaginatum Humb. & Bonpl. ex Willd. J. Presl. y Arceuthobium globosum Hawksw. & Wiens) está más ligada a procesos estocásticos como incendios o tala de árboles; sin embargo, la información que aquí se documenta no se manejó a nivel especie, ni se incluyeron variables que representen algún tipo de disturbio. El análisis indica que la temperatura mínima promedio del periodo más frío (B06) del año fue una de las más importantes en el modelo (Figura 2D, Cuadro 1), lo que se confirma con los resultados de Brandt et al. (2005) quienes mencionan que las temperaturas bajas reducen 95 % la capacidad de dispersión de la semilla de muérdago enano (Arceathobium americanum Nutt. ex Engelm.).
Bosques y plagas forestales
De acuerdo con los datos del INFyS, la observación de plagas forestales en bosques de México ha sucedido en sitios con menos de 15 m2 de área basal por hectárea (Figura 3 B, F, J, N) y menos de tres especies (Figura 3D, H, L, O).
La homogeneidad en estructura y edad de los bosques son determinantes para la incidencia de insectos descortezadores (Raffa et al., 2008); desafortunadamente, en este estudio no se analizó la variabilidad entre sitios para ese tipo de plaga, lo que permitiría corroborar tal hipótesis, aunque con los datos disponibles se observa que los sitios con categorías diamétricas pequeñas son preferidos por barrenadores y muérdagos (Figura 3C y Ñ); mientras que defoliadores y descortezadores se presentan, en mayor medida, en bosques más maduros (Figura 3G y K), lo que se podría deber a una menor resistencia de los árboles a ser atacados, en especial si se trata de los del segundo tipo.
Las altas densidades del arbolado no parecen ser los sitios favoritos para los tipos de plaga aquí estudiados (Figura 3B, F, J, N), puesto que la mayor parte de la presencia de descortezadores se ha observado en sitios con densidades menores de 30 m2 ha-1 y de 500 individuos (de coníferas) ha-1 (Figuras 3I y J), como lo registrado por Íñiguez (1999) quien analizó la densidad arbolada de sitios sanos e infestados en Nuevo León y consignó una densidad media de 455 árboles ha-1; el autor concluyó que no existen diferencias significativas en la densidad de ambos sitios. En contyraste, Cuellar et al. (2013) establecieron que, a mayor densidad de arbolado, aumenta el riesgo de ataque de descortezadores en el sur de dicha entidad.
La contradicción con los resultados que aquí se exponen puede explicarse, principalmente, porque la toma de datos del INFyS no se realiza sobre sitios donde dichos escarabajos existen en condición de plaga, puesto que son un elemento natural de los bosques (Salinas et al., 2010). En el INFyS solo se consigna la presencia o ausencia de los insectos y no se especifican datos sobre su demografía; por lo tanto, sería conveniente hacer estudios en sitios donde se tenga la seguridad de una infestación masiva de plagas forestales.
El análisis de los datos no aporta información sobre alguna especie de plaga forestal en particular, lo que amplía el área de oportunidad para desarrollar estudios posteriores y que, a la vez, representa una limitación. Sin embargo, en ausencia de trabajos de distribución sobre la mayoría de plagas forestales y la inclusión de parámetros dasométricos del bosque, se ofrece una aproximación sobre las zonas forestales que deberían recibir mayor atención por parte de las instancias correspondientes.
Conclusiones
La temperatura es el factor más influyente en la distribución de los defoliadores y descortezadores. La mayor superficie forestal con condiciones climáticas y dasométricas óptimas es Chihuahua para la presencia de barrenadores, que comparativamente, es la más reducida; Oaxaca, Guerrero y Chiapas para defoliadores, que corresponde al territorio más extenso; Chihuahua y Durango para muérdagos; Chihuahua, Michoacán, Estado de México, Nuevo León y Jalisco para descortezadores, principalmente. Las características dasométricas son un componente importante de la distribución de plagas forestales; sin embargo, parece influir en la incidencia de barrenadores y muérdagos, en particular. Es importante observar con cautela la información sobre la distribución de plagas forestales presentada en este documento, puesto que no se refiere a una especie sino a un conjunto de ellas agrupadas por afinidad.

