











 texto em
texto em  Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO 
 Permalink
PermalinkIntroducción
La madera es un grupo importante del carbono orgánico terrestre, cuyos componentes estructurales de la pared celular incluyen celulosa, hemicelulosa, y lignina. Esta última forma una matriz densa que protege los carbohidratos del ataque enzimático; la celulosa también es resistente a la descomposición, en particular en su forma cristalina, la cual es altamente ordenada. Aunque hay muchos agentes, tanto bióticos como abióticos que atacan la madera, los hongos son los principales descomponedores, por ser más efectivos durante el proceso de degradación, pero no se descarta a los insectos, bacterias, algas y xilófagos marinos; mientras que de los agentes abióticos se pueden mencionar la acción de la lluvia, el viento y la radiación solar (Trevisan et al., 2008; Hibbett et al., 2016).
Ninguna especie maderable, ni siquiera aquéllas de reconocida durabilidad natural son capaces de resistir en forma indefinida el ataque de microorganismos, ya que la madera por ser un material de naturaleza orgánica, y en el estado en que se utiliza, ya no es fisiológicamente funcional, por lo que está sujeta a la siguiente etapa de la secuencia natural de cualquier ser vivo: el deterioro y la descomposición (De Oliveira et al., 2005).
Los hongos xilófagos de pudrición blanca, marrón y blanda se han estudiado ampliamente, debido a su capacidad para degradar las paredes celulares de la madera, lo que provoca grandes daños y fallas en los productos en servicio; sin embargo, estos organismos juegan un papel importante en el ecosistema forestal, donde contribuyen significativamente al reciclaje del carbono, ya que los residuos del aprovechamiento son atacados y degradados, por dichos hongos (Singh y Singh, 2014).
La propiedad de la madera que le confiere resistencia al ataque de agentes degradadores químicos, físicos y biológicos, sin tratamiento ajeno de preservación, se denomina durabilidad natural (Paes, 2002). Dichos agentes, en conjunto o por separado, aceleran su proceso de deterioro, por lo que la durabilidad de la madera está influenciada por la interacción de la composición química de sus protectores naturales (extractivos) con las condiciones ambientales (Brischke et al., 2013).
Las comunidades de organismos que destruyen la madera varían de un lugar a otro y en su actividad inciden el pH, la materia orgánica, la microflora y microfauna del suelo (Brischke et al., 2014); la humedad de la pared celular en la madera, puesto que su descomposición no progresa por debajo de 25 % (Thybring, 2013); y por las variaciones en la temperatura ambiente, así como la humedad relativa que crean condiciones favorables o desfavorables para su desarrollo (Johansson et al., 2013). La humedad relativa critica para los microorganismos descomponedores de la madera es de 80 a 95 %, la que a su vez responde a otros elementos como la temperatura ambiente, el tiempo de exposición, además del tipo y las condiciones de la superficie de la madera (Viitanen et al., 2010).
A pesar de los avances en la tecnología para la protección de la madera, los consumidores todavía dependen de las naturalmente durables para la construcción de diversas estructuras, pero desconocen la durabilidad inherente de las especies bajo diferentes condiciones ambientales, por lo que los estudios sobre la durabilidad natural de la madera al daño biológico son de primordial importancia; puesto que, en función de ello, serán las recomendaciones para su óptima utilización, a partir de las condiciones de riesgo en su instalación final, de esta manera se evitan gastos innecesarios para su reposición y el empleo excesivo de madera, lo que impacta positivamente la permanencia de los bosques, al reducir el corte de árboles y, en consecuencia, la amenaza de la deforestación (Paes et al., 2005; Calonego et al., 2010; Sundararaj et al., 2015).
Dada la poca información sobre la durabilidad natural de las maderas locales de la región de El Salto, Durango, este trabajo está encaminado a proporcionar información sobre el nivel de durabilidad, mediante evaluaciones rápidas de laboratorio.
Materiales y Métodos
La durabilidad natural se determinó a través de microcosmos terrestre y exposición a hongos xilófagos en madera de Pinus durangensis Martínez, Pinus cooperi C. E. Blanco, Pinus strobiformis Engelm., Pinus teocote Schiede ex Schltdl. & Cham., Juniperus deppeana Steud., Quercus sideroxyla Humb. & Bonpl. y Arbutus spp. Cinco árboles por especie se recolectaron en diversas áreas de corta de la región de El Salto, Durango; de los cuales, se utilizaron en la elaboración de las probetas secciones de 0.5 m a partir de la base del árbol, y se usó madera de Fagus sylvatica L. como especie testigo, para cumplir con los requisitos que marca la norma,
Exposición a microcosmos terrestre
La evaluación de la durabilidad natural de la madera mediante microcosmos terrestre se llevó a acabo de acuerdo a la Pre-norma Europea 807 (CEN, 2001). Las probetas (55 por especie) consistieron en prismas rectangulares con dimensiones de 10 mm × 5 mm × 100 mm en dirección radial, tangencial y longitudinal, respectivamente; sin considerar la separación de madera de albura y duramen; estas fueron sometidas a un proceso de secado en un horno YAMATO ® DNE910 hasta obtener un peso constante, el cual fue el peso seco inicial. Las probetas se distribuyeron al azar en recipientes de plástico que contenían el suelo recolectado para el experimento; se enterraron 80 % de su longitud (80 mm) con una separación de 20 mm entre probetas (Figura 1). El tiempo de exposición a microcosmos terrestre fue de 32 semanas y el monitoreo se hizo en diferentes etapas de descomposición de la madera, específicamente, después de las 8, 16, 24 y 32 semanas de exposición. El suelo se mantuvo de 80 a 95 % de su capacidad de retención de agua. La durabilidad natural de la madera se consideró como la pérdida tanto de masa como del módulo de elasticidad dinámico.

Cálculo de la pérdida de masa
El procedimiento consistió en determinar la reducción de masa de las probetas en porciento, mediante la diferencia de la masa de madera en condición seca al inicio del experimento y la masa registrada al final. La pérdida de masa se calculó con la fórmula siguiente:

Donde:
Pm = Pérdida de masa (%)
Pi = Peso seco al inicio del experimento (g)
Pf = Peso seco al final del experimento (g)
Módulo de elasticidad dinámico (MOE din )
El MOEdin se calculó en probetas a punto de saturación de la fibra, colocadas sobre dos soportes a una distancia de 0.224 veces su longitud (2.24 mm) a partir del centro, a partir de la aplicación de vibraciones producidas por un golpe en el punto medio de la probeta, las cuales fueron capturadas con la ayuda de un micrófono y el programa Vibration program ® (Divos, 2004). De acuerdo con Carrillo-Parra et al. (2011), este método se utiliza para cuantificar la pudrición en probetas de madera, debido a que la pérdida del MOEdin es mucho más sensible que otras pruebas y presenta valores altamente significativos, incluso en etapas iniciales de deterioro cuando la pérdida de masa aún es baja.
Se utilizó la fórmula derivada de Hearmon (1966) sobre la teoría de la vibración en la madera (Machek et al., 2001):

Donde:
MOE din = Módulo de elasticidad dinámico (MPa)
f n = Frecuencia (Hz)
γ n = (n+0.5)2 = 2.267
π = 3.1416
m = Peso de la probeta por encima del punto de saturación de la fibra (g)
L = Longitud de la probeta (mm)
I = Inercia (mm4)
a = Grosor de la probeta (mm)
b = Ancho de la probeta (mm)
Cálculo del módulo de elasticidad estático (MOE est )
La determinación del MOEest se realizó a partir de ensayos de flexión a un contenido de humedad en equilibrio con el ambiente. La prueba se llevó a cabo en una máquina universal de ensayos mecánicos Instron ®, con capacidad de 60 toneladas de presión; la probeta se colocó sobre dos apoyos a una distancia de 0.224 veces su longitud (2.24 mm), y se le aplicó carga a una velocidad de 1 mm min-1 de acuerdo a lo indicado en la Norma ASTM D 143 (ASTM, 1992). El cálculo se hizo con la siguiente ecuación (ASTM, 1992):
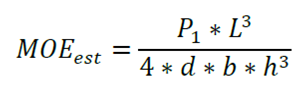
Donde:
MOE est = Módulo de elasticidad estático (kgf cm-2)
P 1 = Carga en el límite de proporcionalidad (kgf)
L = Luz de la probeta, es decir separación entre apoyos (cm)
d = Deformación sufrida por la probeta bajo la carga P 1 (cm)
b = Base de la probeta (cm)
h = Altura de la probeta (cm)
Exposición a hongos xilófagos
Para determinar la durabilidad natural de la madera por exposición a hongos xilófagos, se siguió el procedimiento recomendado en la Norma Europea 113 (CEN, 1996); los hongosutilizados fueron: Coniophora puteana (Schumach.) P. Karst, cuyo número de referencia es BAM 15 (hongo de pudrición parda), y Trametes versicolor (L.: Fr.) Quél. con número de referencia CTB 863A (hongo de pudrición blanca) proporcionados por el laboratorio del Departamento de Biotecnología Molecular de la Madera y Micología Técnica de la Universidad Göttingen, Alemania. Las dimensiones de las probetas (36 por especie) fueron 10 mm × 5 mm × 30 mm en la dirección radial, tangencial y longitudinal, respectivamente; sin considerar la separación de madera de albura y duramen. Posteriormente, se secaron a una temperatura de 103 ± 5 °C hasta obtener un peso constante, el cual constituyó el peso seco al inicio del experimento. Las probetas se esterilizaron en autoclave durante 20 minutos, a una temperatura de 125 °C.
A continuación, se procedió al montaje de las probetas en cajas Petri; para ello, se colocaron 50 mL de malta agar solidificado, como medio de cultivo, y sobre este, el inóculo del hongo (1 cm) y seis probetas montadas en una base de acero inoxidable (Figura 2). Las cajas Petri con las probetas se incubaron en un cuarto de cultivo con ambiente controlado durante un periodo de 16 semanas, a una temperatura de 20 ± 2 °C y una humedad relativa de 65 ± 5 %. La durabilidad se consideró solo como la pérdida de masa en el periodo de exposición, la cual se midió con una báscula de precisión OHAUS ® DV214C.

Clasificación de la durabilidad de la madera
La clasificación de la durabilidad natural de la madera se llevó a cabo de acuerdo con el índice “x”, según la Norma Europea EN 350-1 (CEN, 1994) (Cuadro 1). El índice “x” se determinó al dividir la pérdida de masa promedio de la especie en estudio, entre la pérdida de masa promedio de la especie testigo (Fagus sylvatica L.) como sigue:
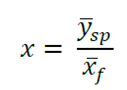
Donde:
x= Índice “x”
y̅ sp =Promedio de pérdida de masa de la especie de interés (g)
x̅ f = Promedio de pérdida de masa de la especie de control (g)
Cuadro 1 Índice de clasificación de la durabilidad de la madera.
| Clase de durabilidad | Descripción | Pérdida de masa según “x” |
|---|---|---|
| 1 | Muy durable | x < 0.15 |
| 2 | Durable | 0.15 > x ≤ 0.30 |
| 3 | Moderadamente durable | 0.30 > x ≤ 0.60 |
| 4 | Poco durable | 0.60 > x ≤ 0. 90 |
| 5 | No durable | x > 0.90 |
Fuente: UNE EN 350-1 (CEN, 1994)
Análisis estadístico
La normalidad de los datos de la pérdida de masa por tiempo de exposición de las especies en estudio, se determinó con la prueba de Shapiro-Wilks modificada, diseñada para el contraste de normalidad. La prueba demostró que las variables pérdida de masa y pérdida de MOEdin no provienen de una población con distribución normal; por tanto, para detectar diferencias estadísticas entre la pérdida de masa por unidad de tiempo de exposición y especie, se realizó la prueba de U de Mann-Whitney para dos muestras independientes, un análisis de varianza no paramétrica y pruebas de comparación de intervalos de la mediana de Kruskal-Wallis (Kruskal y Wallis, 1952), con un nivel de significancia de α=0.05. Los análisis estadísticos se hicieron con el programa InfoStat 2013 (Di Rienzo et al., 2013).
Resultados y Discusión
Exposición a microcosmos terrestre
Pérdida de masa. La pérdida de masa varió en función del tiempo y la especie, con porcentajes promedio de 4 % en J. deppeana, P. cooperi, P. durangensis, P. teocote, F. sylvatica y Pinus strobiformis; de 5 y 10 % para Q. sideroxyla y Arbutus spp., respectivamente (Figura 3).
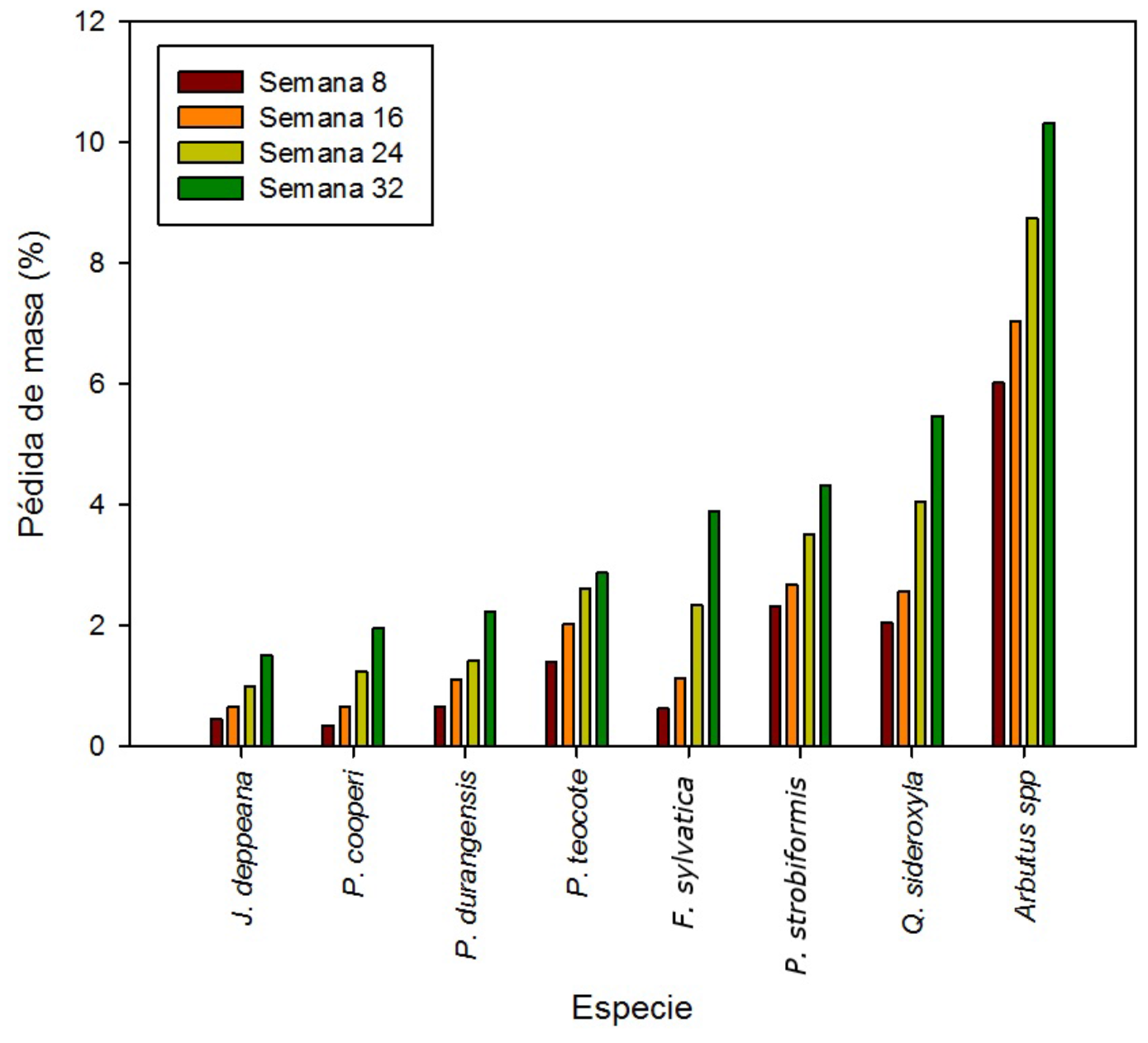
La menor pérdida de masa en la madera de Juniperus deppeana se debe al contenido de extractivos (Eller et al., 2010; Shortle et al., 2010; Kirker et al., 2016) que son los componentes no estructurales de la madera que, a menudo, se concentran en el duramen y actúan como líneas de defensa ante ataques biológicos. Sin embargo, el contenido de extractivos es muy variable, no solo de un árbol a otro, sino dentro de unindividuo. Kirker et al. (2013) registran que la madera de Juniperus virginiana L. tiene una durabilidad natural de moderada a muy durable, pero cuando a la madera se le eliminan los extractivos su durabilidad disminuye hasta clasificarse como poco durable. Existen registros de postes elaborados con madera de Juniperus en el estado de Oregon con 56 años de servicio, en contacto con el suelo (Morrell et al., 1999), lo cual se debe a su valiosa fuente de extractos que proporcionan un alto efecto inhibidor sobre el crecimiento de Trametes versicolor y Gloeophyllum trabeum (Pers.) Murrill (Mun y Prewitt, 2011).
El comportamiento inverso, de menor durabilidad, en Arbutus spp. y Quercus sideroxyla puede explicarse por la ausencia de elementos tóxicos en los extractivos para hacer frente a los agentes xilófagos (Carvalho et al., 2016). Brischke y Rolf-Kiel (2010) señalan que la durabilidad del roble europeo (Quercus spp.) está en debate desde hace mucho tiempo por su clasificación como madera "duradera", ya que los resultados de diferentes estudios comparativos han demostrado, claramente, una menor durabilidad, en especial cuando es expuesta al suelo, pero identificaron como factor más importante el papel de las grietas radiales en su madera, donde realmente inicia su descomposición.
Herrera et al. (2006) citan que la madera, como todo material orgánico, está sujeto a la destrucción por diversos agentes, y que desde el momento en que un árbol se corta y convierte en tejido muerto es codiciado por diversos organismos, los cuales influyen en su degradación. Asimismo, Nájera-Luna et al. (2010) evaluaron la pérdida de masa en la madera de cuatro especies forestales de la región de El Salto, Durango por exposición prolongada a dos tipos de sustrato, y registraron valores promedio de pérdida de masa en la madera de Pinus strobiformis de 7.26 %, en Pinus durangensis de 4.44 %, en Quercus sideroxyla 5.71 % y 5.95 % para Quercus laeta Liebm., resultados aproximados a los obtenidos en el presente estudio.
El análisis de varianza no paramétrica de Kruskal-Wallis para la pérdida de masa de las especies evaluadas fue significativo (p<0.0001), con diferencias estadísticas entre especies. La prueba de comparación de intervalos de la mediana evidenció en Juniperus deppeana la menor pérdida de masa y para Arbutus spp. la más alta. De acuerdo a la clasificación de la Norma Europea EN 350-1, la madera de J. deppeana, P. cooperi y P. durangensis se ubicaron dentro de la clase 3 (moderadamente durable); en la clase 4 (poco durable) se colocó la madera de P. teocote, y se consideraron como no durables (clase 5), las de Pinus strobiformis, Quercus sideroxyla y Arbutus spp. (Cuadro 2).
Cuadro 2 Análisis de varianza no paramétrica para la pérdida de masa en microcosmos terrestre.

“x” = Índice de durabilidad de la madera; Na = No aplica; *Intervalos con la misma letra no son significativamente diferentes, Kruskal Wallis α=0.05; **Significativo al 5 %. GL = Grados de libertad; H = Valor estadístico de la prueba Kruskal Wallis.
La menor pérdida de masa en la madera del género Pinus spp. expuesta al suelo, se explica porque las coníferas tienen solo lignina de monómeros de guayacil (Blanchette et al., 1990). Típicamente, para Abies alba Mill., Pinus sylvestris L., Pseudotsuga menziesii (Mirb.) Franco y Taxus baccata L., la lignina guayacil predomina en sus paredes celulares, en concentraciones superiores en las traqueidas de madera tardía (Baum, 2001). Las ligninas guayacil son, predominantemente, productos de polimerización de alcohol coniferílico; mientras que, las guayacil-siringil están compuestas de partes variables de núcleos aromáticos guayacil y siringil, junto con pequeñas cantidades de unidades p-hidroxifenilo (Fengel y Wegener, 1989). Las proporciones de estos monómeros varían entre tipos de células individuales y capas de la pared celular e influyen fuertemente en la durabilidad de la madera ante hongos de desintegración (Fukazawa, 1992).
Pérdida del MOEdin. El análisis de varianza no paramétrica de Kruskal-Wallis de la pérdida de MOEdin en la madera expuesta al suelo, mostró diferencias significativas (p<0.0001). Pinus spp. y Juniperus deppeana presentaron los menores valores de MOEdin (de 8.09 hasta 9.52 %), mientras que los más altos (de 17.75 hasta 29.07 %) correspondieron a Fagus sylvatica, Quercus sideroxyla y Arbutus spp. (Cuadro 3). Gatto et al. (2012) documentan que los factores intrínsecos de la madera, como las propiedades anatómicas (dimensión de las fibras y frecuencia de rayos), físicas (peso específico y contenido de humedad) y morfológicas (tipos de leño y ángulo del grano); así como el grado de deterioro inciden directamente en la propagación de las ondas de ultrasonido. Sotomayor (2014) y Halabe y Franklin (1998) indican que el uso de métodos no destructivos para conocer alteraciones en la madera, producto de su degradación por factores bióticos y abióticos, está fundamentado en la medición de la velocidad de las ondas mecánicas a través de la estructura, ya que si existe una alteración en la madera por ejemplo, el deterioro del tejido leñoso o colapso del material, la velocidad de la onda mecánica variará como consecuencia del cambio en la estructura del material, el cual resulta en una disminución de sus propiedades elásticas; en este caso, en el módulo de elasticidad calculado a partir de la velocidad y de la densidad del material. Blanchette et al. (1990) argumentan, además, que la velocidad de transmisión de una onda mecánica en la madera se basa en su propiedad para almacenar y disipar energía. Cada especie posee una densidad, una velocidad de onda y un módulo de elasticidad específicos, todos ellos relacionados con la composición química y la morfología de su madera.
Cuadro 3 Análisis de varianza no paramétrica para la pérdida de MOEdin en microcosmos terrestre.

*Intervalos con la misma letra no son significativamente diferente, Kruskal Wallis (α=0.05); **Significativo al 5 %; GL = Grados de libertad; H = Valor estadístico de la prueba Kruskal Wallis.
Machek et al. (1997) señalan que el MOEdin es un buen indicador para evaluar la pérdida de masa en la madera con indicios de deterioro o pudrición, aun durante los primeros estadíos de la infestación, sin que se observe una marcada disminución en la pérdida de masa. Schmidt (2006) establece que la pérdida de masa se debe, principalmente, a la degradación intensiva de los carbohidratos por parte de los hongos de pudrición suave, que pueden causar una pérdida de hasta 50 % en los esfuerzos de flexión, con tan solo 5 % de la pérdida de masa.
Carrillo-Parra et al. (2011) registran que la reducción de MOEdin producida por el deterioro en la estructura de la madera, ocasionado por los organismos presentes en el microcosmos terrestre en la semana 16 para Fagus sylvatica y en Pinus sylvestris es de 79 y 40 %, respectivamente; mientras que en Prosopis laevigata Humb. & Bonpl. ex Willd. M.C.Johnst., los valores de pérdida de MOEdin variaron de 12 a 24 % en los diferentes sitios evaluados. En el presente estudio solo se obtuvieron valores máximos de pérdida del MOEdin inferiores a 29 %.
Exposición a hongos xilófagos
Pérdida de masa por especie de hongo. A través de la prueba no paramétrica de U de Mann-Whitney se determinó que la pérdida de masa fue significativa (p=0.0081) al tipo de hongo empleado en la degradación de la madera (Cuadro 4). Kollmann y C Côté (1968) citan que la distinta actividad de los hongos xilófagos es la causa de la existencia de enzimas diferentes, pues mientras los hongos parásitos xilófagos se alimentan de los compuestos del jugo celular, los saprófitos xilófagos lo hacen de la pared celular; unos atacan, preferentemente, a la celulosa y otros a la lignina, por esto se les puede atribuir la mayor o menor pérdida de masa observada en las especies consideradas en el presente trabajo.
Cuadro 4 Prueba no paramétrica de U de Mann-Whitney para la pérdida de masa por exposición a hongos.
| Hongo | Pérdida de masa promedio (%) | Desviación estándar (%) | Medianas (%) | Promedio intervalos* | U | p |
|---|---|---|---|---|---|---|
| Coniophora puteana (Schumach.) P. Karst | 3.96 | 4.25 | 2.11 | 85.88 a | 5628 | 0.0081** |
| Trametes versicolor (L.: Fr.) Quél. | 16.15 | 21.35 | 4.01 | 107.13 b |
*Intervalos con la misma letra no son significativamente diferente, U de Mann-Whitney (α=0.05). **Significativo al 5 %; U = Valor estadístico de la prueba U de Mann-Whitney.
Carrillo-Parra et al. (2011) evaluaron la durabilidad de Prosopis laevigata, mediante la exposición a hongos degradadores, sus resultados evidencian que Coniophora puteana degradó la madera en menor proporción, con 1.38 % y la disminución de pérdida de masa más alta la provocó Trametes versicolor, con 1.45 %. En el estudio que aquí se documenta, la pérdida de masa inducida por C. puteana fue una cuarta parte menor a la provocada por T. versicolor. Lo anterior se explica por la eficiencia que tienen los hongos en degradar los polisacáridos contenidos en la lignina, ya que después de la ligninólisis, los hongos de pudrición blanca asimilan mejor los polisacáridos restantes, mediante sistemas de glucósido hidrolasas convencionales que contienen enzimas, tanto endo como exoactivas; en cambio los hongos de pudrición parda, también degradan la lignocelulosa de manera eficiente, pero sus sistemas biodegradativos son menos completos (Hatakka y Hammel, 2011).
Pérdida de masa ocasionada por Coniophora puteana. Los resultados del análisis de varianza no paramétrica de Kruskal-Wallis evidenciaron diferencias significativas (p<0.0001), lo cual indica que la pérdida de masa entre especies es diferente. La madera de Arbutus spp., Quercus sideroxyla y Fagus sylvatica presentaron las pérdidas más grandes y P. durangensis, P. strobiformis, P. cooperi y Juniperus deppeana, menores. De acuerdo a lo anterior, la madera de P. durangensis, P. strobiformis, P. cooperi y Juniperus deppeana, según la Norma Europea EN 350-1, se clasificaron como muy durables, consideradas dentro de la clase de durabilidad 1; como durable (clase 2), la madera de P. teocote; moderadamente durable (clase 3) Arbutus spp., poco durable (clase 4) la madera de Quercus sideroxyla (Cuadro 5).
Cuadro 5 Análisis de varianza no paramétrica para la pérdida de masa por exposición a Coniophora puteana (Schumach.) P. Karst.

“x” = Índice de durabilidad de la madera; Na = No aplica; *Intervalos con la misma letra no son significativamente diferente, Kruskal Wallis (α=0.05). **Significativo al 5 %; GL = Grados de libertad; H = Valor estadístico de la prueba Kruskal Wallis.
Pérdida de masa ocasionada por Trametes versicolor. En el Cuadro 6 se resume el análisis de varianza no paramétrica de Kruskal-Wallis para la pérdida de masa. Se observa la existencia de diferencias significativas entre las especies evaluadas. Mediante la prueba de comparación de intervalos de la mediana se evidencian dos grupos estadísticamente diferentes: Arbutus spp., Quercus sideroxyla y Fagus sylvatica que tuvieron la mayor pérdida de masa promedio (30.67 %) y la menor, correspondió a Juniperus deppeana, P. durangensis, P. teocote y P. strobiformis (0.97 %). Con base en la Norma Europea EN 350-1, la madera de Juniperus deppeana, P. durangensis, P. teocote y P. strobiformis se clasifica como muy durable (clase 1); en la clase de durabilidad 3 (moderadamente durable) se ubicó la de P. cooperi y Arbutus spp., la de Quercus sideroxyla se clasificó como poco durable (clase 4).
Cuadro 6 Análisis de varianza no paramétrica para la pérdida de masa por exposición a Trametes versicolor (L.: Fr.) Quél.

“x” = Índice de durabilidad de la madera; Na = No aplica; *Intervalos con la misma letra no son significativamente diferente, Kruskal Wallis (α=0.05). **Significativo al 5 %. GL = Grados de libertad, H = valor estadístico de la prueba Kruskal Wallis.
Carrillo-Parra et al. (2013) determinaron la durabilidad natural de 10 taxones nativos del noreste de México al exponerlas a Trametes versicolor y Coniophora puteana; sus resultados evidencian valores de pérdida de masa por Trametes versicolor de 7.1 hasta 54.1 %, y por Coniophora puteana de 6.3 hasta 48.5 %. Ebenopsis ebano (Berland.) Barneby & J.W.Grimes registró los valores más bajos de disminución de su masa, en ambas especies de fúngicas; y Fagus sylvatica registró la mayor pérdida de masa. Brischke et al. (2017) evaluaron el umbral mínimo de humedad en la madera que permite el inicio de la descomposición fúngica en Picea abies (L.) H.Karst. y Fagus sylvatica con Coniophora puteana y Trametes versicolor bajo simulación real en diferentes escenarios de exposición; confirmaron que los basidiomicetos que destruyen la madera son capaces de degradarla a una humedad relativa alta, sin ninguna fuente externa de agua líquida disponible; ya que los hongos fueron capaces de causar degradación significativa en contenidos de humedad considerablemente por debajo del punto de saturación de la fibra.
Conclusiones
La durabilidad de la madera para Juniperus deppeana, P. cooperi y P. durangensis, mediante microcosmos terrestre se clasifica como moderadamente durable; poco durable para P. teocote y como no durables P. strobiformis, Quercus sideroxyla y Arbutus spp. según la clasificación de la Norma Europea EN 350-1.
La pérdida del módulo de elasticidad dinámico en microcosmos terrestre para la madera de Quercus sideroxyla y Arbutus spp. es de 25 %; mientras que, las cuatro especies de Pinus y Juniperus deppeana pierden 9 % de elasticidad.
La mayor degradación de la madera por tipo de hongo se registra con Trametes versicolor, con una pérdida de masa promedio de 16 %, por 4 % de Coniophora puteana.
Por exposición a Coniophora puteana, la madera de P. durangensis P. strobiformis, P. cooperi y Juniperus deppeana se clasifica como muy durable; P. teocote como durable; Arbutus spp., moderadamente durable; y Quercus sideroxyla, como poco durable.
Por exposición a Trametes versicolor la madera de Juniperus deppeana, P. durangensis, P. teocote y P. strobiformis se clasifica como muy durable; P. cooperi y Arbutus spp., como moderadamente durable; y Quercus sideroxyla como poco durable.
Para usos en los cuales la madera deba estar en contacto directo con el suelo, se recomienda la de Juniperus deppeana, Pinus cooperi y Pinus durangensis, ya que tanto al exponerse a microcosmos terrestres como a hongos degradadores tienen la mejor durabilidad natural. El resto de las especies se recomienda utilizarlas en ambientes protegidos, donde se minimice el contacto con agentes degradadores de la madera.

