











 text in
text in  English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mail Cited by SciELO
Cited by SciELO  Similars in
SciELO
Similars in
SciELO 
 Permalink
PermalinkIntroducción
Debido a la alta fragmentación y reducción de las poblaciones forestales de las zonas áridas y semiáridas del norte-centro de México, el mezquite [Prosopis laevigata (Humb. & Bonpl. ex Willd.) M. C. Johnst.] es una de las principales especies arbóreas o arbustivas que más se utilizan para restaurar ecosistemas forestales (Ríos et al., 2012). Sin embargo, las reforestaciones con este taxon no han sido rentables ecológica ni económicamente por las altas tasas de mortalidad y pobre crecimiento de las plantas en el campo. Dicho desempeño se asocia con un efecto sinérgico entre condiciones ambientales adversas en campo y uso de planta de vivero con capacidad baja de aclimatación a la heterogeneidad de condiciones edafoclimáticas de los sitios por plantar (Ríos et al., 2012; Contreras, 2017).
Para mejorar el desempeño de las reforestaciones con mezquite, se ha enfatizado la necesidad de mejorar la calidad de la planta producida en vivero, en cuanto a atributos morfo-fisiológicos, mediante el estudio de la efectividad de algunas prácticas culturales. El interés principal se ha concentrado en su efecto para promover tasas elevadas de crecimiento tanto del componente aéreo, como del radical y el equilibrio entre ambos. El fin es obtener tallas conforme a la norma NMX-AA-170-SCFI-2016 (DOF, 2016) de 20-30 cm en altura da la parte aérea y diámetros en cuello de la raíz de 3.5 mm o más, dentro de un ciclo de producción de cuatro a cinco meses.
Los trabajos en vivero con la especie referida hasta ahora realizados (Prieto et al., 2013) no tienen el enfoque adecuado para definir una o más prácticas con las que se modifiquen el crecimiento y desarrollo de las plantas, conforme a especificaciones de calidad en los atributos morfo-fisiológicos definidos por las condiciones del sitio que se pretende reforestar; es decir, si se requiere planta grande, con tallo robusto, con área foliar elevada para sitios con competencia vegetal o pastoreo; o planta pequeña, con tallo robusto y un sistema radical suficiente para sitios secos. Lo anterior responde a que, tanto en condiciones de campo como de laboratorio, el crecimiento y el desarrollo de las plantas son afectados por diversos factores ambientales, de los cuales uno es la luz solar disponible.
El fotoperiodo, calidad y cantidad de la luz solar inciden, directamente, en la fotosíntesis de la plantas y demás características fenotípicas y funcionales (Peixioto et al., 2014); por lo tanto, la efectividad de una práctica cultural podría variar de acuerdo con los requerimientos de luz de cada especie en vivero, especialmente, en los aspectos de calidad y cantidad de la radiación solar. Por ejemplo, en la producción de plántulas de Nothofagus leonii Espinosa con diferentes niveles de sombra y fertilización hubo cambios morfológicos y fisiológicos solo por efecto de la sombra (Santelices et al., 2015); en cambio, plántulas de Shorea leprosula Miq. incrementaron su biomasa por la adición de fósforo, cuando crecieron en ambientes de alta luminosidad (Brearley et al., 2007).
Las respuestas fenotípicas de las plantas ante condiciones variadas de luz han sido aprovechadas para modificar sus características morfo-fisiológicas conforme a especificaciones de calidad deseadas; al respecto, Kelly et al. (2015) analizaron el impacto de la cantidad y calidad de la luz para promover en plántulas de Populus tremuloides Michx. una relación elevada entre la proporción de raíz y la parte aérea, y un incremento en los niveles de carbohidratos no-estructurales de las raíces, puesto que esos rasgos predicen un desempeño satisfactorio en campo en los trabajos de reclamación de suelos.
Este enfoque podría implementarse durante la producción de plantas de mezquite en vivero, para manipular su crecimiento e inducir características morfo-fisiológicas más adecuadas a las condiciones del posible sitio de trasplante, principalmente, dado a la disponibilidad de insumos de bajo costo en el mercado que modifican la luz disponible; una de ellas son las mallas sombra, las cuales han tenido un uso amplio en la horticultura para mejorar la calidad de los cultivos e incrementar su rendimiento mediante el color y su fotoselectividad asociada (Stamps, 2009); no obstante, su valoración es necesaria. Por el motivo, el estudio que se describe a continuación tuvo como objetivo cuantificar morfológica y fisiológicamente los cambios de plántulas de mezquite sometidas a condiciones distintas de luz solar.
Materiales y Métodos
Propagación de las plántulas de mezquite
A inicios de marzo de 2016, aproximadamente 300 plántulas de mezquite (Prosopis laevigata) fueron cultivadas a partir de semilla en el vivero forestal del Campo Experimental Valle del Guadiana del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), en Durango, México (23°59'17.1" N, 104°37'35.7" O, 1 880 msnm).
Las semillas de mezquite, cuya germinación se verificó en alrededor de 95 %, provenían de rodales naturales distribuidos en el municipio Aldama, Chihuahua, México. La siembra fue directa en tubetes negros de plástico rígido de 380 mL, llenados con un sustrato compuesto por turba, vermiculita y perlita en proporciones 2:1:1. Previo a la siembra, de acuerdo con las recomendaciones de Prieto et al. (2012), las semillas recibieron un tratamiento pregerminativo con agua caliente y se trataron con Benomilo: Metil 1-(butilcarbomoil) bencimidazol-2-il carbamato 50 % en una dosis de 2 g L-1. Durante el periodo de germinación se aplicaron riegos constantes a saturación.
Tratamientos y diseño experimental
Cuatro semanas después, una vez que la germinación y emergencia se estabilizaron, 225 plántulas uniformes en altura (~5 cm) fueron seleccionadas y organizadas aleatoriamente en grupos de 15, lo cual correspondió al tamaño de la unidad experimental.
Cada una de las cinco condiciones ambientales de luz (intemperie o cielo abierto; malla sombra azul, malla sombra negra, malla sombra roja y malla sombra verde) tuvieron tres repeticiones y se asignaron a las unidades experimentales mediante un diseño completamente al azar. Las mallas utilizadas fueron de la marca PROTEXOL® 80 %.
Se construyeron estructuras metálicas en forma de prisma rectangular, cubiertas completamente por la malla respectiva, a las cuales se les determinó su transmitancia (T; %); para ello, se midió la radiación fotosintéticamente activa (RFA) dentro y por fuera con un ceptómetro (Modelo SF-80, Decagon Pullman) y se empleó la fórmula T= RAF interna/RAF externa. Las mallas presentaron los siguientes valores de transmitancia: malla azul, 26 %; negra, 24 %; roja, 40 %; y verde, 25 %.
En todas las condiciones se monitoreó, diariamente, el valor de temperatura (°C), humedad relativa (%) y radiación global (Wm-2). Las primeras dos variables se midieron con un higrotermómetro digital (Modelo 445702, Extech Instruments) y la última con un piranómetro (Modelo SP Lite, Kipp & Zonen, Delf).
Los valores de radiación global se multiplicaron por una constante de 0.50 para determinar los de RFA, los que, a su vez, se multiplicaron por 4.15 para transformarlos a µmol m-2s-1(Terradas, 2001). Tras ello, se calculó el valor de RFA de las mallas, en función del porcentaje respectivo de transmitancia. En el Cuadro 1 se presentan los valores de las variables climáticas monitoreadas.
Cuadro 1 Valores de las variables climáticas monitoreadas en cada una de las condiciones ambientales de luz.

1T = Temperatura; 2HR = Humedad relativa; 3RFA = Radiación fotosintéticamente activa.
Durante el lapso de crecimiento en cada condición de luz (tres meses), las plántulas se regaron dos veces por semana a saturación y fertilizaron semanalmente con un fertilizante hidrosoluble de propósito general 20-10-20 [20 N: 10 P2O5: 20 K2O + microelementos] (Peters Professional ®), en una concentración base de nitrógeno de 150 mg L-1.
Evaluación del crecimiento, morfología y eficiencia fotosintética
El crecimiento de las plántulas se analizó en términos de ganancia de biomasa a nivel de planta y se midió como tasa de crecimiento relativo (TCR; en mg g-1 día-1). La morfología se determinó mediante el cálculo del área foliar específica (AFE; cm-2 g-1) y con patrones de asignación de biomasa a nivel de hojas tallo y raíz, por lo que se estimó la proporción de biomasa foliar (PBF; %), la proporción de biomasa de tallo (PBT; %) y la proporción de biomasa radical (PBR; %). La eficiencia fotosintética se evaluó con el registro de la tasa de asimilación neta (TAN; mg cm-2 dia-1). El cálculo de las variables de respuesta requirió el muestreo destructivo de una muestra de 15 plántulas de cada condición de luz, al inicio (marzo de 2016) y al final del ensayo (junio de 2016). El procedimiento comenzó con la medición del área foliar utilizando un medidor de área foliar (CI-202, CID, Inc.); continuó con la separación de los demás componentes de las plántulas (tallo y raíz), y finalizó con el secado de estos (incluso las hojas) en una estufa de aire forzado (Felisa FE133A) a 70 °C durante 72 h, y su pesaje en una balanza analítica (Ohaus Pioneer PA224C).
Las observaciones de área foliar y peso de biomasa seca se utilizaron para estimar los valores de cada variable, de acuerdo con las fórmulas descritas por Hunt et al. (2002) que se presentan a continuación:
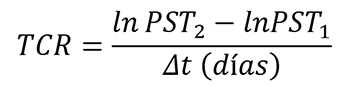
Donde:
PST 1 y PST 2 = Peso seco total de las plántulas en la evaluación inicial y final
Δt = Tiempo trascurrido entre ambas evaluaciones (90 días)
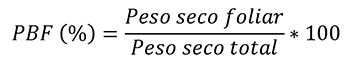
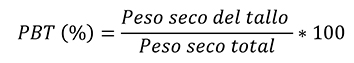


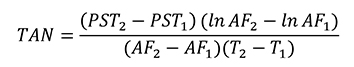
Donde:
PST = Peso seco total de las plántulas en valores absolutos y logarítmicos (ln)
AF = Área foliar de las plántulas valores absolutos y logarítmicos (ln)
T = Tiempo. En los casos 1 y 2 se refiere a la evaluación inicial y final con un lapso entre ellas de 90 días
Análisis estadístico
Se realizó un análisis de varianza no paramétrico mediante la prueba Kruskal-Wallis. En casos requeridos, las medias se compararon por pares. Diferencias estadísticas de P <0.05 fueron consideradas significativas. Los datos se presentan en medias ± desviación estándar. El análisis estadístico se realizó en infoStat (Di Rienzo et al., 2012).
Resultados
El crecimiento, la morfología y la eficiencia fotosintética de las plántulas de mezquite se modificó en función de la condición de luz en la que crecieron; por lo tanto, todas las variables evaluadas (TCR, PBF, PBT, PBR, AFE y TAN) registraron diferencias altamente significativas (P ≥ 0.0001).
La tasa de crecimiento relativo más elevada se tuvo en las plántulas que crecieron en la malla sombra roja. El valor obtenido en esta condición de 7.23 mg g-1 día-1 fue 24 % superior al valor obtenido en la condición de intemperie, que registró la tasa más baja de 5.85 mg g-1 día-1 (Cuadro 2).
Cuadro 2 Valores promedio de las variables de crecimiento, asignación de biomasa y eficiencia fotosintética evaluadas en plántulas de mezquite (Prosopis laevigata (Humb. & Bonpl. ex Willd.) M. C. Johnst.) sometidas a condiciones variadas de luz.

Medias con letras iguales entre filas no son significativamente diferentes (P>0.05). 1TCR = Tasa de crecimiento relativo; 2AFE = Área foliar específica; 3TAN = Tasa de asimilación neta.
En cuanto a la asignación de biomasa, más de 30 % de la producida por las plántulas correspondió a biomasa foliar en la mayoría de las condiciones de malla sombra, excepto en la roja, la cual registró un valor similar a la condición de intemperie con 24 % de PBF. Tal respuesta fue contraria en la PBT, pues acumuló 39 % de biomasa en el tallo, que superó los valores de las otras condiciones, principalmente a los de la intemperie en la que las plántulas producidas ahí destinaron menos de 25 % de la biomasa total a dicho tejido. No obstante, la PBR mostró un efecto distinto en la condición de intemperie a lo anteriormente registrado. En este caso, las plántulas asignaron más de la mitad de su biomasa a la raíz, por lo que superó en más de 50 % las cifras arrojadas por las otras condiciones de luz (Figura 1).
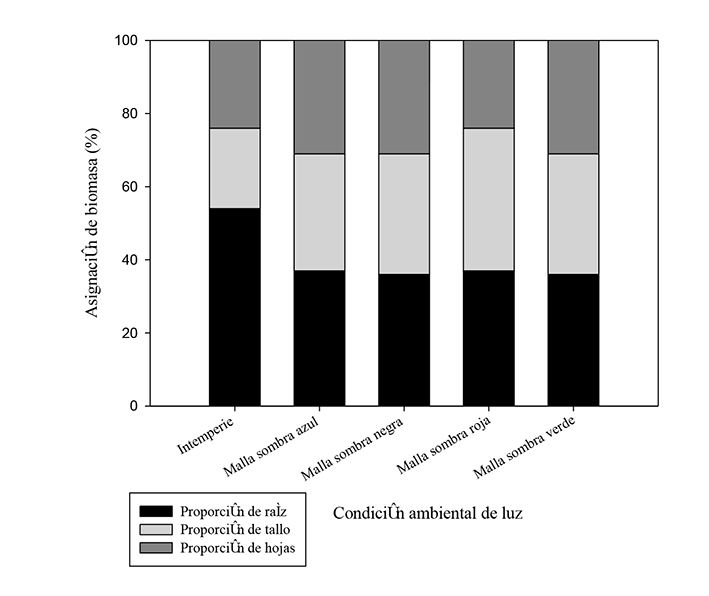
Figura 1 Patrones de asignación de biomasa de plántulas de Prosopis leavigata (Humb. & Bonpl. ex Willd.) M. C. Johnst. ante ambientes contrastantes de luz en vivero.
Por otro lado, la variable AFE, tendió a disminuir conforme aumentó el nivel de radiación, por ello tuvo superioridad en las plántulas que crecieron en malla sombra negra, con valores registrados de 132.50 cm-2 g-1, los cuales fueron 106 % más elevados en comparación con las plántulas de la intemperie, que tuvieron los más bajos, de 64.18 cm-2 g-1 (Cuadro 2).
Por último, fisiológicamente, las plántulas de mezquite experimentaron cambios en su eficiencia fotosintética en función del ambiente de luz al que fueron expuestas. Cuando las plántulas crecieron a la intemperie mantuvieron tasas altas de asimilación neta de 1.55 mg cm-2 dia-1, pero se redujeron hasta 128 % en el ambiente de malla sombra negra que registró el valor mínimo, de 0.68 mg cm-2 dia-1 (Cuadro 2).
Discusión
Durante la estancia de las plántulas de mezquite en el vivero, las condiciones ambientales de luz modificaron su crecimiento, morfología y eficiencia fotosintética. Uno de los factores implicados en este efecto fue el microambiente generado en cada condición, específicamente, en lo que concierne a temperatura y humedad. Tal y como se describe en la horticultura (Stamps, 2009). Las condiciones microambientales de temperatura y humedad provocadas por la malla sombras (Cuadro 1) al parecer favorecieron el crecimiento de los ejemplares estudiados, en especial a las de la malla roja, cuyo crecimiento fue 24 % superior en comparación con aquéllas que se produjeron a la intemperie.
No obstante, la evidencia obtenida sugiere una mayor atribución del crecimiento y de las modificaciones morfológicas y fisiológicas de las plántulas a la cantidad y calidad de irradiación solar que recibieron en cada condición, en particular en las malla sombras en las que se registraron niveles diferentes de radiación y una fotoselectividad, posiblemente distinta en función del color de la malla.
Este punto es coherente con los aspectos que se atribuyen al uso de mallas sombra de colores en el cultivo de hortalizas (Stamps, 2009); con lo demostrado de forma experimental en condiciones de vivero con plántulas de Bauhinia variegata (L.) Benth. (Mazzini y Pivetta, 2014); y con las respuestas de las plántulas de mezquite en las variables evaluadas.
Al respecto, las mallas sombra promovieron que morfológicamente las plántulas manifestaran las respuestas fenotípicas típicas de aclimatación de una especie pionera, ante un ambiente heterogéneo de luz (Pearcy, 2007). Niveles bajos de radiación se tradujeron en una mayor asignación de biomasa a las hojas y tallos; mientras que, niveles altos favorecieron una mayor asignación de biomasa a la raíz.
De esa manera, cuando las plántulas crecieron en las mallas, más de 60 % de la biomasa fue asignada a la parte aérea (hojas y tallo); a la intemperie, más de 50 % de la biomasa fue destinada a la raíz (Figura 1).
Estos cambios en la relación raíz/parte aérea se pueden atribuir, aunque este aspecto no fue evaluado, a las variaciones de la cantidad de luz recibida en la relación rojo/rojo lejano, puesto que dicho factor es importante para las respuestas mediadas fitocromáticamente (Batschauer, 1998).
Kelly et al. (2015) sustentaron con plántulas de Populus tremuloides, que una relación rojo/rojo lejano elevada propició el crecimiento y desarrollo de la raíz; en cambio, una relación rojo/rojo lejano baja estimula el crecimiento y desarrollo aéreo. Así, la condición de intemperie supone una elevada exposición a esta relación, mientras que las condiciones de malla sombra implican una exposición baja, la cual a su vez pudo variar hacia niveles más inferiores, debido al color de las mallas, tal y como se demuestra en un estudio realizado por Shahak et al. (2004).
Sin embargo, contrario a lo expuesto previamente, Kelly et al. (2009) demostraron de forma experimental, cambios opuestos entre los patrones de asignación de biomasa de seis especies australianas subtropicales sujetas a diferentes gradientes de luz, y los promovidos en las plántulas de mezquite, pues en estos taxa una mayor disponibilidad de luz resulta en un incremento de biomasa aérea y una disminución de la biomasa asignada a la raíz.
Lo anterior puede asociarse con el grado de plasticidad fenotípica de las especies, puesto que Cheng et al. (2013) en un estudio cuyo contexto es similar al de Kelly et al. (2009) y al del presente trabajo revelaron una plasticidad fenotípica baja de los patrones de asignación de biomasa a la luz, en cinco taxa subtropicales siempre-verdes de hoja ancha.
Por otra parte, a nivel foliar, las variaciones en la disponibilidad de luz inducidas en cada condición evaluada promovieron cambios morfológicos, de tal manera que una escasa disponibilidad de luz hizo que las hojas fueran más delgadas e incrementaran su área foliar específica (AFE), mientras que una oferta más grande de luz derivó en un decremento de AFE, y, por lo tanto, en un engrosamiento de las hojas. Lo anterior se constata tanto con los resultados de la malla sombra negra como con los de intemperie, ya que representaron cada caso, respectivamente (Cuadro 2).
Varios autores coinciden con los hallazgos de este trabajo y argumentan que el efecto en las hojas corresponde a cambios morfo-funcionales que las plantas promueven, para usar más eficientemente la radiación disponible y así mantener una capacidad fotosintética adecuada, lo que depende de la disponibilidad de la misma, ya sea aumentado la superficie foliar por cantidad de biomasa invertida, o a través del incremento de la densidad de las hojas (Cheng et al., 2013; Yang et al., 2013; Tang et al., 2015).
Fisiológicamente, los resultados obtenidos comprueban que las plántulas de mezquite experimentaron cambios en su eficiencia fotosintética en función del ambiente de luz al que fueron expuestas. El hecho de que su TAN, cuando crecieron a la intemperie, haya sido 128 % superior a las de malla sombra negra, refleja que una mayor disponibilidad de luz promueve ganancias elevadas de carbono a nivel de planta. De ahí, se esperaría que entre más se limite el paso de la luz con las mallas, menores son las ganancias de carbono. Si bien, los valores de TAN de la condición de intemperie sugieren una ganancia superior de carbono en las plantas, por ser los más altos (aunque no fueron estadísticamente distintos a los de la malla roja); su crecimiento, comparado con el de las plántulas de la malla roja contradicen tal supuesto, ya que en realidad las de las mallas fueron las que presentaron crecimientos más grandes, al registrar un valor de 24 % superior (Cuadro 2).
El efecto positivo de la malla roja podría atribuirse a la promoción de un balance más adecuado entre las ganancias de carbono, debidas a la TAN y las pérdidas de carbono, asociadas con la respiración, las cuales, probablemente fueron menores en comparación con las plántulas de intemperie; esto se debe a que en sombra, la baja disponibilidad de luz limita la ganancia de carbono, pero reduce las tasas de respiración, por lo que las plantas mantienen un balance positivo de carbono (Pearcy, 2007).
Además, como se consigna en un estudio con plántulas de Torreya grandis Fortune ex Lindl. en el que se les determinó la intensidad óptima de luz (Tang et al., 2015), la sombra generada por la malla pudo haber evitado daños por fotoinhibición e incrementado la eficiencia de las hojas para captar y absorber mayor radiación con el aumento del valor de AFE, que, aunque fue menor comparado con las plántulas de las otras mallas sombra, superó al de la intemperie en 34%, pero fueron estadísticamente similares (Cuadro 2).
Por lo tanto, los valores de TAN y los de AFE pudieron contribuir al crecimiento de las plántulas de la malla roja, independientemente, del grado de variación que cada uno tiene para explicar el crecimiento, aspecto en el que aún existe controversia dada la naturaleza de cada variable y de los trabajos realizados al respecto, por ejemplo Villar et al. (2005) y Tomlinson et al. (2014).
Finalmente, cabe señalar que las plántulas de todas las condiciones evaluadas alcanzaron los estándares morfológicos mínimos de calidad requeridos por la Norma Mexicana NMX-AA-170-SCFI-2016 (DOF, 2016) dentro de un ciclo de producción de cuatro meses. De ahí que los hallazgos de este estudio tienen implicaciones prácticas para los trabajos de reforestación, ya que pueden manipularse características de las plántulas en vivero de acuerdo con especificaciones morfológicas y fisiológicas definidas por las condiciones edafoclimáticas del sitio que se desea reforestar.
Sin embargo, se requieren nuevos planteamientos experimentales para evaluar el desempeño de las plantas en campo ante una variedad de condiciones ambientales, tras haber sido aclimatadas a un ámbito específico de luz en el vivero.
Conclusiones
Las plántulas de mezquite tienen buena capacidad de aclimatación a condiciones contrastantes de luz (lo cual es una expresión de su plasticidad fenotípica) durante su periodo de crecimiento en vivero. Una elevada exposición a la luz reduce el crecimiento y la formación de biomasa aérea, pero estimula una mayor formación de biomasa radical; en cambio, un sombrado con malla sombra, independientemente, del color promueve un crecimiento superior e invierte los patrones de asignación de biomasa, que favorece la formación de biomasa en el componente aéreo, mismo que experimenta cambios morfológicos a nivel foliar con el incremento de su superficie foliar, lo cual es contrario al engrosamiento que las hojas presentan cuando se les expone a niveles altos de radiación, sin la protección de las mallas, de las cuales, la roja aumenta la eficiencia fotosintética, permitiéndoles tener mayores ganancias de carbono.

