










Servicios Personalizados
Revista
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista mexicana de ciencias forestales
versión impresa ISSN 2007-1132
Rev. mex. de cienc. forestales vol.8 no.42 México jul./ago. 2017
Articles
Susceptibility and prevalence of Eucalyptus pellita F.Muell. to the cancer caused by Chrysoporthe cubensis (Bruner) Gryzenhout & M. J. Wingf. at the Colombian Orinoquia
1Universidad Distrital Francisco José de Caldas.
2Universidad Distrital Francisco José de Caldas.
3Corporación Nacional de Investigación y Fomento Forestal - CONIF.
Eucalyptus cancer caused by Chrysoporthe cubensis is one of the most limiting diseases for Eucalyptus plantations in the tropical region. In order to confirm the cancer causal fungus observed in the commercial plantations of E. pellita in the Colombian Orinoquia and explore the possible difference in the susceptibility of some commercial clones of the region, the pathogen was isolated from diseased tree tissue. The strains obtained were inoculated in young plants belonging to ten clones, including one control of E. urophylla under greenhouse conditions. The mean wound length was compared for up to 53 days after inoculation, with two isolates of the pathogen. The prevalence of cancer was also estimated in a 7-year-old E. pellita plantation. Based on symptoms, and macro and microscopic morphological characters, the isolates were identified as Chrysoporthe cubensis and with the use of the molecular markers ITS5 and ITS4 as C. cubensis and C. doradensis. All clones evaluated showed susceptibility to Chrysoporthe attack. Although eucalyptus cancer is a common disease, this is the first record of C. cubensis and C. dorandesis in E. pellita plantations in Colombian Orinoquia. Given the favorable conditions for the development of the disease in the region, it is recommended to continue the selection and evaluation efforts to have the disease tolerant material.
Key words: Eucalyptus canker; Chrysophorte doradensis Gryzenh. & M.J. Wingf.; incidence; Colombian Orinoquia; severity; symptoms
El cancro del eucalipto causado por Chrysoporthe cubensis es una de las enfermedades más limitantes para las plantaciones de Eucalyptus en la región tropical. Con el objeto de confirmar el hongo causal del cáncer observado en plantaciones comerciales de E. pellita en la Orinoquia Colombiana y explorar la posible diferencia en susceptibilidad de algunos clones comerciales de la región, se aisló el patógeno a partir de tejido enfermo de árboles adultos y las cepas obtenidas se inocularon en plantas jóvenes pertenecientes a diez clones, incluido un testigo de E. urophylla en condiciones de invernadero. Se comparó la longitud promedio de la herida desarrollada durante 53 días después de la inoculación, con dos aislamientos del patógeno. También se estimó la prevalencia del cancro en una plantación de E. pellita de 7 años de edad. Con base en los síntomas, y a caracteres morfológicos macro y microscópicos, los aislamientos se identificaron como Chrysoporthe cubensis y con el uso de los marcadores moleculares ITS5 e ITS4 como C. cubensis y C. doradensis. Todos los clones evaluados evidenciaron susceptibilidad al ataque de Chrysoporthe. Aun cuando el cáncer del eucalipto es una enfermedad común, constituye el primer registro de C. cubensis y C. dorandesis en plantaciones de E. pellita en la Orinoquia Colombiana. Dadas las condiciones favorables para el desarrollo de la enfermedad en la región, se recomienda continuar los esfuerzos de selección y evaluación para contar con material tolerante a la enfermedad.
Palabras clave: Cáncer del eucalipto; Chrysoporthe doradensis Gryzenh. & M.J. Wingf.; incidencia; Orinoquia Colombiana; severidad; síntomas
Introduction
The disease known as cancer or Eucalyptus cancer, caused by Chrysoporthe cubensis Brunes Gryzenhout & M. J. Wingf., is one of the most limiting diseases in Eucalyptus commercial plantations (Van Heerden and Wingfield, 2001; Gryzenhout et al., 2004; Souza, 2008).
Chrysoporthe cubensis infects the trees through wounds, usually at the base of the stem of young trees, causing stunting, wilting, banding and cracking of stems (Gryzenhout et al., 2004; Nakabonge, 2006); besides, produces exudations or enzymatic secretions that can be toxic (Bernal et al., 2009), and that in advanced conditions can lead to death, mainly of young plantations in several species of eucalyptus (FAO, 2008).
In Colombia one of the diseases most frequently found in eucalyptus plantations is the cancer caused by C. cubensis in E. grandis W. Hill ex Maiden and the only effective management measure is the selection and sowing of tolerant clones (Rodas et al., 2005; Mizerit, 2009). In the Colombian Orinoquia the selection of species of the genus Eucalyptus has been made, from different species and provenances, from which it is concluded that E. pellita F.Muell. is the species that reaches the highest volume per unit area with 350 m³ ha-1 (Reforestadora de la Costa, 2009).
Clonal propagation, while maximizing commercial timber production, also has precautionary factors to evaluate, such as the susceptibility of a clone to the attack of pathogens. It is to be expected that for this species, the number of hectares planted with clones in the medium and short term will increase significantly (Nieto and Gasca, 2010). For this reason, it is important to incorporate the disease resistance variable into the selection programs of E. pellita clones suitable for extensive planting in the Colombian Orinoquia, even though the species is recognized in the literature because of its low incidence of pests and diseases (Alfenas et al., 1983; Borralho and Nieto, 2012). Therefore, the present investigation was aimed at confirming the identity of the causing agent of the cancer of E. pellita and to estimate the susceptibility of some clones to the cancer caused by this pathogen in seedlings of 3 months old, as well as the prevalence of the problem in seven-year-old plantations.
Materials and Methods
Study area
The study was carried out in a forest core of 3 000 ha of which 265.21 ha have been established with species of the Eucalyptus genus. The plantations are located on the plateau of San Pedro in the municipality of Villanueva, Department of Casanare, Colombia, at the geographical coordinates 4°57 "N and 73°94" W with an average altitude of 420 m.
The study area is located in the tropical moist forest, with a monomodal precipitation regime which favors the development and dispersal of eucalyptus cancer throughout the year (excepting December, January, February and March), since C. cubensis presents optimum growth at elevated temperatures and precipitation of more than 2 000 mm annually (Gryzenhout et al., 2004).
Obtaining of isolates and identification of the pathogen
Samples of plant material with symptoms and signs of Eucalyptus cancer were obtained from bark, wood and apical meristems of two-year old E. pellita and E. urophylla S. T. Blake four-year-old trees. All samples were placed in properly labeled plastic bags and transported in a portable refrigerator at a temperature of about 4 °C (Bernal et al., 2009), prior to their analysis in the Health Laboratory of the Distrito Francisco José de Caldas, Vivero headquarters, in Bogotá.
The thirty-one field-collected samples were individually disinfected in a 2 % sodium hypochlorite (NaCl) solution for two minutes, and rinsed twice with sterile distilled water (Bernal et al., 2009). Two replicates were obtained from each sample, one of which was kept in refrigeration at 4 °C and another was used to obtain reproductive structures of the fungus and the first isolates. Each sample was examined in detail with the help of a Discovery V8 Carl Zeiss stereomicroscope with which the ascostroma, perithecial neck and mycelium were identified on the plant tissue. Isolates were obtained from these samples by direct transfer of conidia masses, monosporic culture from suspension of conidia in sterile distilled water (ADE) and seeding sections of infected plant tissue in culture medium. In all cases, Merck-branded 2 % Agar Maltose (Merck) culture medium was used (Gryzenhout et al., 2004; Nakabonge 2006; Bernal et al., 2009, Juárez et al., 2013). The isolates were preserved by the cryopreservation, lyophilization and sterile mineral oil methods.
Corroboration of the causal agent was based on macroscopic characteristics such as color and texture of the mycelium and microscopic morphologies by the observation and identification of conidiophores and conidia. The microscopic characterization of C. cubensis and C. doradensis Gryzenh. & M.J. Wingf. was performed by direct staining and staining in lactophenol blue. In addition, sections of the 25 μm thick infected material were obtained with a LEICA RM 2255 microtome and added with a non-coagulating fixative with chromic acid and acetic acid (adapted from Reig et al., 2002). Transverse sections of tissues were made with Lactophenol Blue and basic Fushina (González et al., 2011). The macroscopic and microscopic characteristics were compared with that described for C. cubensis by Gryzenhout et al. (2004) and Nakabonge (2006) and C. doradensis were compared with those described by Gryzenhout et al. (2005). In addition, sequences of the molecular markers ITS4 and ITS5 were obtained using the sequencing and molecular analysis service of the Instituto de Genética de la Universidad Nacional de Colombia. (Genetics Institute of the National University of Colombia).
In order to select two virulent isolates, four of the isolates obtained from monosporic cultures were compared and inoculation was performed in 100 branches from ten clones of seven - year - old E. pellita following the methodology developed by Juárez et al. (2013). The length of the lesion developed from the point of inoculation with C. cubensis was used as the response variable and the treatments were determined by the clone and the inoculation strain (strain 17, 26, 29, 31 and control). Additionally, the daily mycelial growth of each isolate in culture medium in the Petri dishes was compared as another parameter equivalent to the invasion capacity that was used as an indicator of virulence (Díaz and Lecuona, 1995; Ochoa, 2004). Based on these tests, isolates 17 and 26 (obtained in E. pellita) were selected to perform the assay.
Susceptibility of Eucayptus pellita and Eucalyptus urophylla seedlings to the inoculation by C. cubensis under greenhouse conditions
Seedlings from ten E. pellita clones and one from E. urophylla (Table 1) were displayed in 17 cm diameter plastic pots and kept in an isolated greenhouse built for this study. The second species was also selected for the inoculation in the greenhouse, from its resistance to the attack by C. cubensis (Alfenas et al., 1983; Van Heerden and Wingfeld, 2001).
Table 1 Eucayptus pellita F.Muell. and Eucalyptus urophylla S. T. Blake clones used to compare their susceptibility to Chrysoporthe cubensis (Bruner) Gryzenhout & M. J. Wingf.
| Clone | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | URO1 |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of seedlings | 30 | 30 | 30 | 15 | 11 | 8 | 30 | 30 | 27 | 30 | 18 |
The assay was arranged in three completely randomized blocks determined by the isolation (17 and 26) and the other block for the control treatment (inoculation of seedlings with 2 % AM culture medium). Each block in turn was divided into sub-blocks, which corresponded to the number of replicates, so that in each sub-block all the clones evaluated were represented.
From the subcultures of each isolate of the pathogen, 4 mm diameter mycelial discs were cut with the aid of a pressure cuff and sterile needles. Each disc was stored inside pieces of sterile, non-adherent gauze within sterile Petri dishes, moistened with ADE.
To perform the inoculation, a wound was initially made at the base of the stem of each seedling, using a 4 mm diameter punch. Subsequently, the wet gauze section was placed with the mycelial disk, in direct contact with the wound made in the stem of the seedling and covered with Parafilm.
The development of the lesion was measured weekly for one month with a calibrator (Mitutoyo, CD 6 "CS of 0.01 mm accuracy) for a total of four measurements. At each measurement the precaution was taken to use nitrile gloves and seal again with Parafilm to prevent the entry of any type of contaminant.
Estimation of prevalence of the Eucalyptus cancer in a seven-year old E. pellita plantation
The prevalence of the disease (incidence and severity) was assessed in a stand made up by 570 seven-year old E. pellita individuals. Incidence was considered as the affected trees per cent, while severity was graded with a nominal scale for estimation of this condition of C. cubensis in E. pellita plantations, with a total number of six categories (adapted from Paredes et al., 2010). (Table 2).
Table 2 Scale for estimating Chrysoporthe cubensis Brunes Gryzenhout & M. J. Wingf.severity in Eucayptus pellita F.Muell. plantations.

Source: Paredes et al., 2010.
The calculation of the average damage degree, incidence and severity was made as follows (Parra et al., 1999):
Average damage degree (GMD)
GMD=∑((Ni *CDi)/(N))
Where:
i = 1 to 6
Ni = Number of trees in the damage category i
CDi = Damage category i
N = Total of measured trees
Incidence (INC)
INC = (Number of affected trees * 100)/total number of plants
Intensity or damage severity
I = (∑((Ni * CDi)/(K * N))*100
Where:
i = 1 to 6
Ni = Number of trees in the damage category i
CDi = Damage category i
N = Total of measured trees
K = Greatest category mayor
Data processing and analysis
Data obtained from daily mycelial growth of the isolates, lesion size of inoculated branches and inoculum seedling lesion size were organized and processed with descriptive statistics, and the assumptions of normality, homoscedasticity and independence were evaluated by means of the tests of Kolmogorov-Smirnov and Levene, prior to comparison using a variance analysis (ANOVA) and Duncan's post-hoc test. When the data did not present normal distribution, transformations were tested by the Box Cox method, using the SPSS 15.0 statistical program Otherwise, the analysis was performed using non-parametric statistics. The response variables evaluated were diameter of mycelial growth for the daily evaluation of isolates in laboratory as virulence factor and length of the lesion developed from the point of inoculation for the inoculation in branches and seedlings.
Results
Pathogen agent
The affected trees in the field and from which samples of diseased material were taken showed symptoms and signs characteristic of the cancer caused by C. cubensis, such as swelling and fissures in the stem, gum redness associated with cracks on the cortex, presence of pycnidia, long pycnidial neck and globose base with reddish, coffee and black hue. In addition, the release of conidia was observed in mucilaginous masses of yellow tones accumulating on the ostium, composed of conidia (Figure 1). Conidiophors with bifurcations, hyaline with rectangular basal cells and globular appearance (Figure 2B-D). The conidia are unicellular, oval, non-septate hyaline, with bright exudates (Figure 2C), the size of conidia, conidiophores and pycnidia agree with those described in literature (Table 3). Isolates obtained from the field samples produced pycnidia and conidia after approximately 30 days of incubation (27 °C).

A. Grouping of pycnidia on wood. B - C. Pycnidia on bark with mass of conidia in the ostium. D-E. Pycnidia on wood with mass of conidia in the ostium.
Figure 1 Pycnidia of Chrysoporthe cubensis Brunes Gryzenhout & M. J. Wingf. with mass of conidia ejected through the ostium.
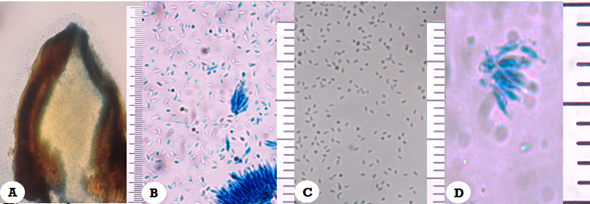
A. Transverse section of pycnidia with conidia (blue-stained by Lactophenol Blue staining (10 X); B, D, Conidiophores (40 X, 100 X); C. Oval-shaped conidia (40 X)
Figure 2 Photomicrographs of Chrysoporthe cubensis Brunes Gryzenhout & M. J. Wingf.
Table 3 Size of conidia, conidiophores and pycnidia of Chrysoporthe cubensis Brunes Gryzenhout & M. J. Wingf.

Identification by DNA extraction and sequencing with the ITS markers are consistent in 99.9 % with sequences published in GenBank as C. cubensis for the isolation 17 obtained in E. urophylla (Access number JN942342.1) and C. doradensis for the strain 26 (Access number JN942330.1) from E. pellita.
The colonies of the four assessed isolates (codes 17, 26, 29 and 31) did not differentiate each other in terms of average mycelial growth (Kolmogorov-Smirnov, for K-S of a sample: 0.365). Strain 17 had the highest average of daily growth (0.75 mm day-1), followed by strain 31 (0.74 mm day-1). Strain 17 exhibited colonies that reached maximum diameter of mycelium (9 cm) at 5 days of measurement, and for strain 31 (9 cm) at 8 days of measurement (Table 4).
Table 4 Growth of Chrysoporthe colonies in laboratory conditions.
| Isolation | Host species | Diameter (cm) | S |
|---|---|---|---|
| 17 | Eucayptus urophylla | 0.75 | 2.7 |
| 26 | Eucayptus pellita | 0.62 | 1.8 |
| 29 | Eucayptus pellita | 0.65 | 2.3 |
| 31 | Eucayptus urophylla | 0.74 | 2.7 |
N = Five colonies
The inoculation of eucalyptus branches with the isolates with Chrysoporthe induced the development of typical lesions (Figure 3) and were significantly higher in comparison with the control (inoculated with AM), (df = 4, p> 0.05, p = 0.000). The average length of lesions was 63.4 mm while in the control, 26.3 mm. No significant difference was observed between the strains in the lesion size nor in the lesion induced in the different clones (Table 5).

Figure 3 Injury caused by Chrysoporthe cubensis Brunes Gryzenhout & M. J. Wingf. in Eucayptus pellita F.Muell. branches in the second week of inoculation. Necrosis in branches of E. pellita associated to the inoculation point of C. cubensis.
Table 5 Sub-sets (1, 2) statistically different from mean length of the lesion caused by four strains of Chrysoporthe sp. in eucalyptus branches (Duncan's test).
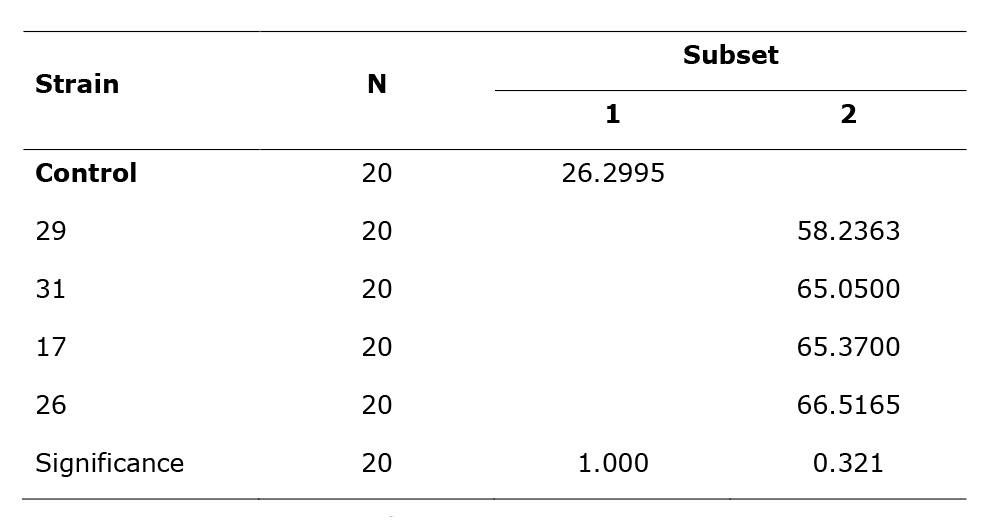
Duncan. Average
The means for the groups in the homogeneous subsets are shown in Table 5. The sample size of the harmonic mean was 20.00.
Susceptibility of greenhouse seedlings
All clones evaluated showed susceptibility to inoculation with strains 17 and 26 of Chrysoporthe (df = 222, p <0.05); however, no statistically significant differences were found between the 11 clones nor between the two assessed Chrysoporthe isolates (Table 6).
Table 6 Length (mm) of injury after 53 days of inoculation with Chrysoporthe cubensis Brunes Gryzenhout & M. J. Wingf.

Seedlings developed progressive and rapidly advanced symptoms of cancer. After 53 days of inoculation with Chrysoporthe, they presented increased wound length, forming larger sores. In the inoculated seedlings in turn, exudation of reddish shade of viscous and bright texture was evidenced, which is also characteristic in Eucalyptus trees affected by the fungus; also, these seedlings formed additional sores at the inoculation site and which were displayed along the stem (Figure 4).

A, B, C, D. Seedlings with presence of exudates of reddish hue at the inoculation site. E. Primary bifurcated or dead meristems. F. Galls along seedling stem.
Figure 4 Development of wounds caused by Chrysoporthe infection in Eucayptus pellita F.Muell. seedlings, 53 days after inoculation.
The disease progressed gradually and caused the death of several of the evaluated seedlings (Table 7). Treatment with strain 26 brought together the largest number of dead individuals (25). Clones 5 026 (20 %) and 5 002 (16 %) achieved the highest mortality rates attributable to treatment.
Table 7 Absolute and relative Eucalyptus seedling mortality inoculated with Chrysoporthe per treatment.

With strain 17, a 22.2 % mortality rate was verified, corresponding to 20 of the total assessed (90) in this treatment; the clone with the highest number of deaths was 5 013 with 30 % of the seedlings in the treatment described, followed by 5 632 (20 %). Finally, in the control, the death of five seedlings was recorded in the last measurement, which constitutes 5.68 %, of which three were of clone 1 110 (Table 7).
Prevalence of Chrysoporthe sp. in plantations
Of all clones tested (20), 54 % had an incidence of 90 to 100 %. Despite the high values, 51 % had a severity between 50 and 60 %, so the disease was not in advanced stages of development.
The 570 trees evaluated in the plantation of E. pellita revealed an average degree of damage (GMD), that between 0.6 and 3.5, that is, with mild to intermediate conditions. However, 51.3 % of the clones had mean damage levels of 2.5 to 3.0, corresponding to deep fissures with exudate of reddish tonality, reproductive structures (pycnidia) and, in some cases, swelling at the base of the tree.
At the plantation, the ten clones of E. pellita evaluated in greenhouse had an average degree of damage between 2.47 and 3.33. They exhibited bark detachment and reddish exudations mainly in clones 1 110 and 5 026. On the other hand, the incidence of C. cubensis of these clones in particular, is damaging between 80 and 100 % of the total of individuals, with severity of 49 % and 67 %.
Discussion
The symptoms and signs of the pathogen observed in the E. pellita plantation and those analyzed under laboratory conditions coincide with that reported in previous studies for the attack of C. cubensis (Van Heerden and Wingfield, 2001; Gryzenhout et al., 2004; Nakabonge, 2006; FAO, 2008; Paredes et al., 2010; da Silva et al., 2010; Juárez et al., 2013).
The expressions of cancer observed in the field are characteristic of the disease caused by C. cubensis. Likewise, the macroscopic morphological study of the fruiting bodies and the fruiting pattern, and the laboratory microscopic study coincided with the descriptions of Van Heerden and Wingfield (2001), Gryzenhout et al., (2004) and Nakabonge (2006) for this pathogen.
The symptoms, signs and morphological characteristics of C. cubensis and C. doradensis are similar and at the microscopic level they are only differentiable by discrete differences in the shape of the conidia and coloration of the conidial mass (Gryzenhout et al., 2004; Gryzenhout et al., 2005). For the above, it was initially considered that the two isolates corresponded to two isolates of C. cubensis. However, based on the ITS sequence, it was possible to determine the presence of C. doradensis, which has also been recorded in Eucalyptus plantations in Ecuador (Gryzenhout et al., 2005). In spite of being a common occurrence in Eucalyptus, in this work, C. cubensis and C. doradensis are known for the first time in E. pellita plantations in Colombian Orinoquia.
The virulence of a pathogen depends, to a large extent, on the capacity for invasion and the production of toxic substances (Ochoa, 2004). In addition, mycelial growth allows the development of the fungus as it absorbs nutrients, which, in turn, make it possible to form new mycelia (Chávez et al., 2011). In this study, colony size as a criterion for selection of the most virulent strains of obtained in the laboratory was not easy to apply, since there were no significant differences in the size of the colony in relation to time.
Probably the strains of C. cubensis obtained correspond to sources of inoculum of low genetic variation, since they were collected from hosts within the same batch or nearby; therefore, the branch inoculation response did not show a significant difference. It is possible that the hosts from which the isolates were obtained have been infected by a common primary inoculum, that is, with genetically related.
The E. pellita and E. urophylla seedlings studied showed susceptibility to the attack of C. cubensis and C. doradensis isolates, and there were no differences between clones or between isolates of the fungus. Similar results were reported by Juárez et al. (2013) in which all clones of E. urophylla and E. grandis evaluated were attacked by C. cubensis, but not consistent with other studies in which E. pellita is classified as one of the less susceptible species of the Eucalyptus genus To Chrysoporthe (Alfenas et al., 1983), nor with studies in E. urophylla and E. grandis that showed different levels of tolerance between clones to the attack of Chrysoporthe cubensis and, in turn, different degrees of virulence between strains of cancer (Van Heerden and Wingfeld, 2001,;Chen et al., 2010; Juárez et al., 2013).
Cloning of C. cubensis-resistant genotypes has allowed the control of eucalyptus cancer (Van Heerden and Wingfeld, 2001; da Silva et al., 2010). Although laboratory and greenhouse tests can be used as indicators of susceptibility of E. pellita, they should be interpreted with caution, since in previous studies the behavior of the disease in juveniles does not necessarily represent susceptibility in plantations of greater development (Juárez et al., 2013); so, it is advisable to continue with evaluation of productive clones at different ages of development. The establishment of future plantings of these clones may represent economic and environmental risks, according to the planted area objectives in Orinoquia with E. pellita (Mizerit et al., 2013).
It is important to emphasize that the study area meets the appropriate climatic conditions for the development of eucalyptus cancer caused by C. cubensis and C. dorandesis as high temperatures and precipitation. Therefore, it is advisable to continue with research aimed at establishing in more detail the behavior of the fungus in its hosts and, from there, to establish which of them are optimal to carry out a forest plantation of E. pellita (da Silva et al., 2010). In spite of finding Chrysoporthe cubensis in the Eucalyptus plantations in Villanueva, the presence of the disease in the native vegetation associated to the plantations was not evident.
Conclusions
Seedlings of the E. pellita and E. urophylla clones evaluated showed similar response to inoculation with C. cubensis and C. doradensis under greenhouse conditions. Laboratory and greenhouse tests should be interpreted with caution so the recommendation is to continue the evaluation of productive clones at different ages.
The susceptibility observed in the assessed clones could represent risks for the establishment of future plantations in the region where there are naturally both inoculum and climatic conditions appropriate for the development of the disease, i. e., high temperatures and precipitation, for long periods.
Acknowledgements
The Corporación Nacional de Investigación y Fomento Forestal (CONIF) financed the present study through the agreement Colciencias - Reforestadora de la Costa - Refocosta SAS - CONIF within the project " Consolidación de la estrategia de mejoramiento genético de la especie Eucalyptus pellita F. Muell., para el establecimiento de una fuente de suministro de material clonal de alta calidad en la región de la Orinoquia colombiana”(Consolidation of the breeding strategy of the species Eucalyptus pellita F. Muell. for the establishment of a source of supply of high-quality clonal material in the region of the Colombian Orinoquia) executed under the Cierre de Brechas # 586 (Closing Breaches # 586) announcement of Colciencias of 2012. Refocosta company facilitated access to plant material, plantations and installation and provided logistical support and the Engineer Mónica Sarmiento supported the planning and development of field work. The Universidad Distrital Francisco José de Caldas (Francisco José de Caldas District University) facilitated the use of equipment and physical spaces.
REFERENCES
Alfenas, A., R. Jeng and M. Hubbes. 1983. Virulence of Cryphonectria cubensis on Eucalyptus species differing in resistance. European Journal of Plant Pathology 13:197-205. [ Links ]
Bernal L., I. et al. 2009. Identificación del agente patógeno del cancro del eucalipto en plantaciones del sureste de México. Revista Ciencia Forestal en México 34 (105): 17-36. [ Links ]
Borralho, N. y M. Nieto. 2012. Eucalyptus para la Orinoquia: retos y oportunidades. El mueble y la madera 75: 26-33. [ Links ]
Chávez, K., S. Rodríguez, J. Cervantes y J. Barranco. 2011. Enzimas y toxinas de hongos entomopatógenos, su aplicación potencial como insecticidas y fungicidas. Sociedades Rurales, Producción y Medio Ambiente 11(22):143-160. [ Links ]
Chen, S., M. Gryzenhout, J. Roux, Y. Xie, M. Wingfield and X. Zhou. 2010. Identification and Pathogenicity of Chrysoporthe cubensis on Eucalyptus and Syzygium spp. in South China. The American Phytopathological Society 94 (9): 1146-1150. [ Links ]
da Silva G., L., M. de Resende, D. Lau, L. Rosse, A Alves and A. Alfenas. 2010. Genetic control of Eucalyptus urophylla and E. grandis resistance to canker caused by Chrysoporthe cubensis. Genetics and Molecular Biology 33(3): 525-531. [ Links ]
Díaz, B. y R. Lecuona. 1995. Evaluación de cepas nativas del hongo entomopatógeno Beauveria bassiana Bals. (Vuill.) (Deuteromicotina) como base para la selección de bioinsecticidas contra el barrenador Diatraea saccharalis (F.). Revista Agriscientia 12: 33-38. [ Links ]
Drumond, M., S.Á. Iêdo B., M. C. Oliveira y E. M. Leite. 2001. Selección de especies y procedencias del género Eucalyptus para la región costera de Bahía, Brasil. In: Proceedings of IUFRO International Symposium. 3-15 September. 2001. Valdivia, Chile. CD-ROM. [ Links ]
Food and Agriculture Organization of the United Nations (FAO). 2008. Forest pest species profile, Chrysophorte cubensis (Bruner) Gryzenhout & M. J. Wingfield. http://www.fao.org/forestry/13579-02412a08b7a10ee4b3bfc2aa0f8754d82.pdf (24 de julio de 2013). [ Links ]
González, M., D. Diamont y B. Gutiérrez. 2011. Técnica de tinción de estructuras fúngicas con colorantes vegetales como una alternativa no contaminante. Revista Biagro 23(1): 65-68. [ Links ]
Gryzenhout, M. et al. 2004. Chrysoporthe, a new genus to accommodate Cryphonectria cubensis. Studies in Mycology 50: 119-142. [ Links ]
Gryzenhout, M. et al. 2005. Chrysoporthe doradensis sp. nov. pathogenic. Fungal Diversity 20: 39-57. [ Links ]
Juárez P. et al. 2013. Patogenicidad de Chrysoporthe cubensis en clones de Eucalyptus grandis y E. urophylla en el sureste de México. Revista Madera y Bosques 19(1): 17-31. [ Links ]
Mizerit T., L. H. 2009. Estudio comparativo en dos áreas semilleras establecidas en plantaciones de Eucalyptus urophylla en Oaxaca. Tesis de maestría. Colegio de Postgraduados. Campus Montecillo. Montecillo, Edo. de Méx. pp. 53-70. [ Links ]
Mizerit T., L. H. et al. 2013. Impacto de Chrysoporthe cubensis (Bruner) Gryzenhout & M. J. Wingfield en dos áreas semilleras de Eucalyptus urophylla S. T. Blake en Santiago Yaveo, Oaxaca. Revista Mexicana de Ciencias Forestales 4(16): 64-75. [ Links ]
Nakabonge, G. 2006. A study of Chrysophorthe and Cryphonectria species on Myrtales in Southern and Eastern Africa. Doctoral dissertation. University of Pretoria. Pretoria, South Africa. 158 p. [ Links ]
Nieto, V. y G. Gasca. 2010. Experiencias y avances en el manejo de Eucalyptus pellita F. Mell. en la Orinoquia Colombiana. CONIF, Ministerio de Agricultura, Refocosta S.A. Bogotá, Colombia. 98 p. [ Links ]
Ochoa A., N. A. 2004. Evaluación de los factores de virulencia de hongos infecciosos en camarón blanco (Litopenaeus vannamei). Tesis de Maestría en Manejo de Recursos Marinos. Centro Interdisciplinario de Ciencias Marinas. Instituto Politécnico Nacional. La Paz, BCS., México. 85 p. [ Links ]
Paredes D., E., J. Jasso M., J. López U. y D. Alvarado R. 2010. Chrysoporthe cubensis (Bruner) Gryzenhout & M. J. Wingf. en plantaciones de Eucalyptus grandis Hill ex Maiden en Oaxaca. Revista Mexicana de Ciencia Forestales 1(2): 17-36. [ Links ]
Parra, P., J. Valencia y M. González. 1999. Manual de detección y evaluación sanitaria en eucalipto. INFOR. Santiago de Chile, Chile. Manual 24. 124 p. [ Links ]
Reforestadora de la Costa. 2009. Programa generación de una estrategia de manejo tecnológico para Eucalyptus pellita F. Muelluna especie de rápido crecimiento. CONIF, Ministerio de Agricultura, Refocosta S.A. Bogotá, Colombia. 2 p. [ Links ]
Reig, J., F. García, A. Ibars y E. Estrelles. 2002. Aproximación al estudio anatomo-histológico y de la micorriza de Ophioglossum lusitanicum L. Butil. Sociedad Micológica de Madrid 7: 55-197. [ Links ]
Rodas, C. A., M. Gryzenhout, H. Myburg, B. Wingfield and M. Wingfield. 2005. Discovery of the Eucalyptus canker pathogen Chrysoporthe cubensis on native Miconia (Melastomataceae) in Colombia. Plant Pathology 54 (4): 460-470. [ Links ]
Souza, H. G. 2008. Resistencia do Eucalipto ao cancro de Chrysoporthe cubensis e Botryosphaeria sp. Dissertação (mestrado) em Agronomia (Proteção de Plantas). Facultade de Ciências Agronômicas. Câmpus de Botucatu. Universidade Estadual Paulista “Julio de Mesquita Filho”. Botucatu, São Paulo, Brasil. 109 p. [ Links ]
Van Heerden, S. W. and M. J. Wingfeld. 2001. Genetic diversity of Cryphonectria cubensis isolates in South Africa. Mycology Research 105(1): 94-99. [ Links ]
Received: November 12, 2016; Accepted: February 06, 2017
 Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
Este es un artículo publicado en acceso abierto bajo una licencia Creative Commons
