











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Artículo en XML
Artículo en XML Referencias del artículo
Referencias del artículo
 Enviar artículo por email
Enviar artículo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La distribución de carbono se refiere al carbono destinado a la producción de diferentes partes de la planta (Friend et al., 1994); se han propuesto dos teorías: la óptima (Bloom et al., 1985), que establece que una planta lo dirige hacia el órgano que adquiere la fuente más limitante, y la de biomasa alométrica (Niklas y Enquist, 2001) que lo refiere a la sección aérea o a la raíz, en función de su tamaño, calculado por peso seco o por longitud. La cantidad de carbono que se dedica a cada una de estas dos estructuras depende de las condiciones de crecimiento e historia de vida, y es importante porque influye en la capacidad de supervivencia de los pinos después de la plantación (South, 2000). Por lo tanto, es conveniente conocer esta información en especies utilizadas en reforestación y en conservación.
En relación a las condiciones de crecimiento, una disminución en la masa de la parte aérea y de la masa radical en Pinus pinaster Ait., P. pinea L., P. canariensis C. Sm. ex DC., P. halepensis Mill. se presentó en situaciones de poca disponibilidad de agua en la cámara de crecimiento (Chambel et al., 2007) y en invernadero (Aranda et al., 2010); en este último ambiente se ha observado el mismo comportamiento en latifoliadas cultivadas (Guarnaschelli et al., 2003).
Por otra parte, las plantas confieren una cantidad de carbono mayor para la construcción de raíces laterales en sitios infértiles (Paz, 2003) o húmedos (Markesteijn y Poorter, 2009), así como para construir una raíz profunda si las semillas proceden de sitios con baja disponibilidad de agua y se desarrollan en invernadero con suficiente agua y nutrientes (Climent et al., 2008).
Así, la distribución de carbono está sujeta a factores como la procedencia o el ámbito de crecimiento y es factible evaluarla de forma indirecta mediante variables asociadas al mismo, lo que facilita la identificación de especies con mejor respuesta en condiciones naturales. Por lo tanto, en la hipótesis de este trabajo se planteó que existe variación significativa en la distribución de carbono en especies con diferente supervivencia en campo: Pinus michoacana Martínez con 76 % (Gómez-Romero et al., 2012); P. pseudostrobus Lindl. de 2 y 68 % (Muñoz et al., 2011; Gómez-Romero et al., 2012) y P. martinezii E. Larsen, por su valor de conservación.
Materiales y Métodos
Las variables que se consideraron fueron peso seco y fresco de la parte aérea y de la raíz, longitud específica de la raíz y su morfología en plántulas de las especies seleccionadas cultivadas en vivero.
Se recolectaron 6 960 semillas de Pinus martinezii en Los Azufres, Municipio Ciudad Hidalgo, Michoacán, México (Morales, 2014) equivalentes a 100 g y lo mismo de semillas de P. michoacana Martínez y de P. pseudostrobus Lindl., que fueron proporcionadas por la Comisión Forestal del Estado de Michoacán.
De este material, se tomaron cien semillas por especie para este estudio. Se les aplicó un tratamiento pregerminativo de estratificación, que consiste en someterlas a una temperatura de 4 °C durante 15 días. Las semillas ya pre-tratadas se sembraron en cavidades de unicel en sustrato turba-agrolita en una proporción de 1:2 (volumen:volumen), para luego colocarlas en una cámara de crecimiento- germinación bajo condiciones de 25 °C, 75 % de HR y 14 horas de fotoperiodo.
De las plántulas obtenidas (con un porcentaje de germinación entre 85 y 95 %), se seleccionaron 20 de 6 cm de altura por especie, que se les trasplantó en un sistema de mini-rizotrones (Climent et al., 2008). Estos mini-rizotrones se formaron a partir de tubos de polivinilo de cloruro (PVC) de 10 cm de diámetro y 35 cm de longitud, los cuales fueron cortados por el centro, de lo que resultaron dos mini-rizotrones de cada tubo (Figura 1).
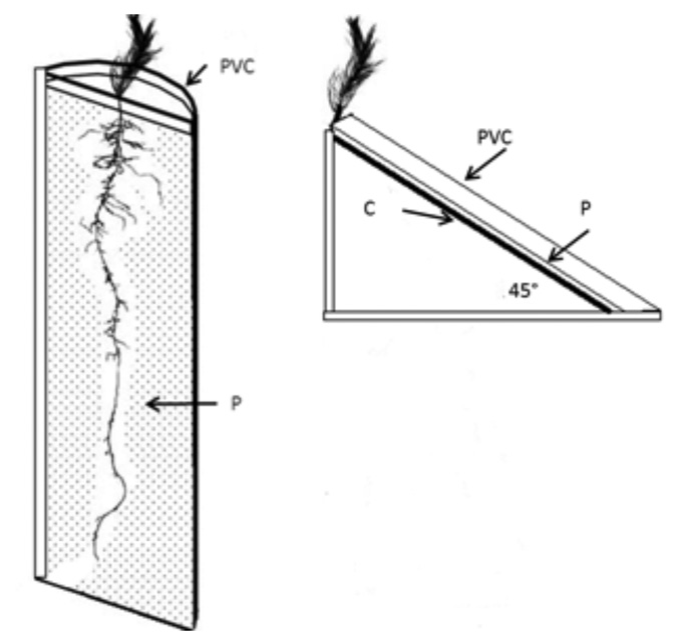
P = Policarbonato celular transparente; PVC = Pilivinilo de cloruro; C = Lámina de contrachapado de madera de 3 mm de espesor.
Figura 1 Mini-rizotron para el crecimiento de plantas de Pinus pseudostrobus Lindl., Pinus martinezii E. Larsen y Pinus michoacana Martínez.
A los 150 días del trasplante, se practicó un análisis destructivo con 10 plantas por especie seleccionadas al azar. De cada una se registró el peso fresco y el peso seco de la parte aérea y de la raíz obtenidos mediante balanza analítica (Ohaus modelo PA214).
De la segunda se determinaron, también, la longitud especifica de raíz (LER) (relación entre la longitud de raíz y peso seco de la raíz en cm g-1), la densidad (relación entre peso seco de raíz y el volumen fresco en g cm-3 y volumen fresco de raíz (método de desplazamiento de agua en una pipeta graduada de 1 ml). Se evaluaron todas las raíces laterales para efectos del número, longitud (cm) y diámetro (mm). Su clasificación se basó en la raíz principal como referente, a partir de la cual las raíces más cercanas, serían las de primer orden. La longitud total (LTRL) se calculó mediante la sumatoria de todas ellas.
Después de las mediciones en fresco, las plántulas se secaron en un horno (Terlabo modelo TE-H35D) a 60 °C por siete días hasta llegar a peso constante (g) en cada sección.
El análisis del sistema radical se llevó a cabo mediante el programa ImageJ 1.4 (Macinstosh). Se utilizó un ANOVA mediante un diseño completamente al azar con el paquete S-pluss (Tibco Software Inc., 2000) con un valor de P < 0,05 para la significancia.
Resultados y Discusión
Los resultados indican que Pinus michoacana alcanzó valores significativamente mayores en peso fresco y seco de las partes estudiadas respecto a P. martinezii y P. pseudostrobus. El crecimiento de primera especie fue de entre 72 y 97 % en la parte aérea y de 70 y 140 % en la raíz. Entre P. martinezii y P. pseudostrobus no se presentó diferencia significativa en dichas variables (Cuadro 1).
Cuadro 1 Valores promedio en biomasa aérea y biomasa de raíz.
PF = Peso fresco; PS = Peso seco; A/R = Proporción parte aérea y raíz; ± = Error estándar. Letras diferentes indican diferencia estadísticamente significativa (P < 0.05 y n = 10 por especie).
El crecimiento de las tres especies no se ha reportado considerando un mismo sistema. Estos resultados sugieren que P. pseudostrobus y P. martinezii provienen de sitios similares, tal como se ha observado en otras especies de pinos (Salazar et al., 1999) y que la supervivencia de estas dos especies posiblemente es similar.
Además, P. michoacana podría tener mayor potencial de supervivencia que P. pseudostrobus y P. martinezii, dado que presenta mayor capacidad de almacenamiento de agua (peso fresco de parte aérea y raíz) e incorpora como tejidos cantidad de carbono asimilado (peso seco) bajo condiciones similares de crecimiento. Esto coincide con los resultados reportados por Gómez-Romero et al. (2012), quienes observaron mayor supervivencia de plantas de P. michoacana respecto a plantas de P. pseudostrobus en un sitio degradado.
La proporción parte aérea/raíz (A/R) presentó valores entre 1.58 y 1.96 en las tres especies. Estudios previos mostraron que las especies de coníferas destinan mayor carbono para construir la parte área que la raíz (Levy et al., 2004; Mokany et al., 2006), lo cual coincide con los resultados de este experimento. Esta distribución de carbono (proporción A/R) coincide con la teoría de la distribución de biomasa alométrica (Niklas y Enquist, 2001) e indica que las tres especies provienen de sitios similares en relación a la disponibilidad de agua. En este sentido, Cregg (1994) observó mayor distribución de carbono a la parte aérea en las plantas de Pinus ponderosa Dougl. ex Laws. de diferentes localidades con alta disponibilidad de agua.
Las variables de la raíz se presentan en el Cuadro 2 y éstas no mostraron diferencia significativa entre P. martinezii y P. pseudostrobus, mientras que la diferencia entre estas dos especies y P. michoacana fue significativa, lo cual es similar a lo observado en peso de raíz y parte aérea. La distribución de carbono medida a través de variables morfológicas de raíz no se ha reportado en condiciones de adecuada disponibilidad de agua. La longitud específica de raíz (LER) varió entre las diferentes especies evaluadas en este experimento y fue mayor al 100 % en las plantas de P. pseudostrobus y P. martinezii en relación a P. michoacana. Dichos resultados coinciden con los reportados por King et al. (1997), quienes observaron diferencia significativa de la LER en dos especies de pino. La LER mostró una relación inversamente proporcional con las variables longitud total de raíces laterales (LTRL), longitud promedio de raíces laterales (LPRL), la densidad de raíces (DR) y volumen de raíz (VR). La diferencia entre P. pseudostrobus y P. martinezii en relación a P. michoacana fue entre 40 y 90 %. La relación inversa entre dichas variables coincide con previos reportes. Ostonen et al. (2007) observaron plantas con menor LER y mayor capacidad de exploración de suelo. Por lo tanto, P. michoacana podría tener una mayor supervivencia en comparación a las otras especies por su capacidad de explorar más volumen de suelo debido a su menor LER. La relación entre la LER y la mayor exploración de suelo en estas especies de coníferas no depende del número total de raíces laterales, sino que la exploración depende de la longitud de raíces laterales del primer orden.
Cuadro 2 Valores promedio de las variables de la raíz.
| Especie | LER (cm g-1) | LTRL (cm) | LPRL (cm) | DR (g cm-3) | VR (cm3) |
|---|---|---|---|---|---|
| Pinus pseudostrobus Lindl. | 335.3 ± 22.1 a | 182.9 ± 8.0 b | 1.3 ± 0.08 b | 0.21 ± 0.02 ab | 0.49 ± 0.06 b |
| Pinus martinezii E. Larsen | 331.7 ± 43.9 a | 199.3 ± 26.9 b | 1.0 ± 0.09 b | 0.18 ± 0.02 b | 0.56 ± 0.04 b |
| Pinus michoacana Martínez | 154.2 ± 15.0 b | 327.9 ± 21.9 a | 1.9 ± 0.17 a | 0.26 ± 0.01 a | 0.83 ± 0.05 a |
LER = Longitud especifica de raíz; LTRL = Longitud total de raíces laterales por planta; LPRL = Longitud promedio de raíces laterales, DR = Densidad (DR); VR = Volumen de la raíz. ± = Error estándar. Letras diferentes indican diferencia estadísticamente significativa (P < 0.05 y n = 10).
Los valores de morfología de raíz muestran una clara diferencia en la distribución de carbono en las tres especies (Cuadro 3). P. martinezii presentó una longitud de la raíz principal y un número de raíces de primer orden mayor significativamente en relación a P. pseudostrobus y P. michoacana. La longitud y el diámetro de las raíces laterales de primer orden fueron mayores significativamente en P. michoacana en relación a las otras dos especies. El mayor diámetro las raíces del primer, segundo y tercer orden se presentó en P. michoacana y P. pseudostrobus en relación a P. martinezii. Estos resultados de morfología de raíz hacen evidente la variación en cuanto a distribución de carbono entre especies de pino y aportan más evidencia de que la mayor supervivencia de P. michoacana podría estar relacionada con la longitud y diámetro de las raíces laterales, seguido de P. pseudostrobus y por último P. martinezii.
Cuadro 3 Valores promedio de número, longitud (cm) y diámetro (mm) de raíces laterales en los diferentes ordenes del sistema radical.
| Número de raíces | Longitud de raíz | Diámetro de raíz | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Pma | Pp | Pmi | Pma | Pp | Pmi | Pma | Pp | Pmi | |
| Principal | 1 ± 0.0 a | 1 ± 0.0 a | 1 ± 0.0 a | 30.1 ± 2.4 a | 4.6 ± 0.3 b | 4.6 ± 0.5 b | 1.1 ± 0.005 c | 1.9 ± 0.009 b | 2.4 ± 0.015 a |
| Primer orden | 49 ± 6.6 a | 13 ± 0.9 c | 16±0.7 b | 2.8 ± 0.5 c | 8.6 ± 0.8 b | 12.6 ± 0.7 a | 0.6 ± 0.002 b | 0.9 ± 0.004 a | 0.9 ± 0.002 a |
| Segundo orden | 108 ± 14.2 a | 111 ± 10.1 a | 126±10.9 a | 0.6 ± 0.1 a | 0.7 ± 0.1 a | 0.9 ± 0.2 a | 0.5 ± 0.001 b | 0.7 ± 0.002 a | 0.7 ± 0.002 a |
| Tercer orden | 32 ± 7.9 ab | 22 ± 3.2 b | 39 ± 7.2 a | 0.4 ± 0.1 a | 0.3 ± 0.1 a | 0.4 ± 0.1 a | 0.5 ± 0.003 b | 0.6 ± 0.001 a | 0.6 ± 0.002 a |
| Cuarto orden | - | 0.72 ± 0.5 a | 3 ± 1.6 a | - | 0.1 ± 0.02 a | 0.3 ± 0.20 a | - | 0.1 ± 0.0006 a | 0.2 ± 0.009 a |
| Total raíz lateral | 189.81 ± 17.8 a | 146.72 ± 7.5 b | 183.81 ± 14.7 a | ||||||
Pma = Pinus martinezii E. Larsen; Pp = Pinus pseudostrobus Lindl.; Pmi = Pinus michoacana Martínez.
± = Error estándar. Letras diferentes indican diferencia estadísticamente significativa (P < 0.05 y n = 10 por especie).
Conclusiones
Existe variación en la distribución de carbono a nivel inter-especie y coincide con el porcentaje de supervivencia. Esta relación no fue consistente mediante variables como peso seco de parte aérea y raíz, proporción parte aérea y raíz, volumen y longitud de raíz, sino que otras variables como longitud específica de raíz (LER), longitud total de raíces laterales, la longitud y el diámetro de raíces por orden de raíz son parámetros con mayor exactitud. Por último, se sugiere que en trabajos posteriores se considere la evaluación morfológica y de tamaño, tanto en parte aérea como de raíz bajo diferentes condiciones de crecimiento para elucidar con mayor exactitud la variación de la distribución de carbono de las especies de coníferas en México y poder incluir la LER como característica morfológica para calificar la calidad de planta.

